
Abstract
Liquid crystalline phases of DNA and nucleosome core particles can be formed in vitro, indicating the crucial roles of these phases in the maintenance and compaction of genomes in vivo. In the present study, sequential levels of liquid crystalline decompaction were identified in highly purified nuclei of Karenia papilionacea in response to the gradual chelation of divalent counterions by ethylenediaminetetraacetic acid (EDTA); the decompaction was observed using polarizing light microscopy, confocal laser scanning microscopy, and transmission electron microscopy and further confirmed utilizing microcalorimetry. Nested fibrous coils in 150 nm arc-like bands of chromatin were observed in the early stages of chromosomal decompaction. The microcalorimetry spectra of isolated nuclei revealed that the dynamic processes of nuclear decompaction occurred in a nonlinear manner; in addition, an EDTA-sensitive thermal transition between 60°C–70°C, corresponding to a liquid-crystalline-phase transition of chromosomes, was found. The results suggested that nested coils of fibrous chromatin filaments are responsible for the establishment and stabilization of the liquid crystalline and birefringence features of the chromosomes of dinoflagellates. The results also indicated that positively charged divalent counterions play significant roles in modulating liquid crystalline phases to compact the chromosomes of dinoflagellates.
Introduction
DNA-binding divalent cations have been quantitatively analyzed in native chromosomes using scanning ion microscopy; this analysis suggested that a high concentration of the chromatin-binding cations Ca2+ and Mg2+ is essential for the neutralization of negatively charged DNA and the maintenance of chromosome structure in both classical eukaryotes and dinoflagellates (Strick et al., 2001; Levi-Setti et al., 2008). Basic core histones in eukaryotes significantly contribute to the neutralization of the charge of chromosomal DNA (Levi-Setti et al., 2008). However, this counterionic interaction will be maximally reduced in dinoflagellates because their chromosomes lack core histones and have only a small number of histone-like proteins (Levi-Setti et al., 2008). Ultrastructures of ethylenediaminetetraacetic acid (EDTA)-treated cells and spectroscopic mapping of divalent cations in chromosomes confirmed a role of counterions in the neutralization of DNA in liquid crystalline chromosomes (LCCs) (Herzog and Soyer, 1983; Levi-Setti et al., 2008). The LCCs reported in dinoflagellates are mainly from the haploid stage and have simple armless karyotypes and large sizes, which are especially suited for studying chromosome organization. Studying the condensation and decompaction of LCCs could reveal the possibly unique mechanism this special organism possesses for regulating the chromosomal structure; the structure that is responsible for the establishment of the liquid crystalline and birefringence features of the chromosome might also be determined. The present study reports the use of highly purified dinoflagellate nuclei, a semi-in vivo system, in the study of chromosomal decompaction.
Materials and Methods
Culture conditions
All experiments employed batch cultures of Karenia papilionacea, which was isolated from the pier of the Hong Kong University of Science and Technology (Yeung et al., 2005). K. papilionacea was maintained in the L1 medium at 18°C under a photon flux of 50 μmol m−2·s−1 from fluorescent tubes (Phillips daylight) in 12-h light/dark (L/D) cycles. The cells were routinely harvested at the stationary phase with a concentration of ∼103 cells·mL−1.
Isolation of nuclei
All isolation steps were performed on an ice surface. The stationary-phase cells were harvested in Falcon tubes using centrifugation at 1000 g for 10 min at 4°C. The cell pellets were resuspended in a total of 50 mL of culture per 100 μL of freshly prepared nuclei extraction buffer (NEB), which consisted of 1 mM Tris, 1.5 mM CaCl2, 10 mM KCl, 0.1 mM dithiothreitol (DTT), 0.5% NP-40, and 1× protease inhibitors (0.5 μg·mL−1 PMSF, 0.01 μg·mL−1 aprotinin, 0.01 μg·mL−1 pepstatin, and 0.01 μg·mL−1 leupeptin) at pH 7.4. Then, the suspension was transferred to a 1.5-mL Eppendorf tube and incubated for 10 min. After centrifugation at 14,000 g for 10 s using an Eppendorf bench-top centrifuge, the supernatant was decanted, and the pellet was resuspended in 500 μL of NEB. This step was repeated until the pellet became mostly white in color. Finally, the high-yield purified intact nuclei were stored at the desired concentration with NEB. The integrity of the isolated nuclei was checked using polarizing light microscopy (PLM) and fluorescence microscopy.
PLM and metripol system
For PLM observations, cultured cells or isolated nuclei were sealed between a glass slide and a coverslip using Cytoseal 60. The birefringence of the liquid crystalline nuclei was examined using an Olympus BX51 polarizing microscope. Photomicrographs were taken using an attached Pixera Penguin 600 CL digital camera.
The anisotropic optical properties of intranuclear chromosomes were measured using a Metripol birefringence imaging system (Oxford Cryosystems) (Geday et al., 2000). The system employed a motorized rotating polarizer and a fixed circular analyzer. The use of the Metripol system in quantifying the birefringence of LCCs has been recently demonstrated (Chow et al., 2010). The sample preparation for the Metripol system is similar to the preparation procedure for polarizing microscopy. A series of 50 images for every 7.2° of polarizer rotation in each sample was taken for quantification. The integrated software was used to control the measurement process and analyze the resulting data in three separated quantitative images as the magnitude of phase retardance (|sinδ|), which is directly proportioned to the birefringence and thickness of the sample, the orientation angle, which is equivalent to the extinction angle (φ) and related to the angle between the extraordinary and ordinary optical axes and the transmittance (Io) along the pathway direction of incident polarizing light.
Confocal laser scanning microscopy
For confocal laser scanning microscopy (CLSM), isolated nuclei pellets from 100 mL of cultured cells were suspended in 100 μL of NEB buffer with various EDTA concentrations and stained using 50 μg·mL−1 Sytox Green. The nuclei were then observed using an inverted Leica SP2 CLSM with a pair of orthogonal polarizers in the transmission channel to observe intranuclear liquid crystalline features. Digitized images of the optical series of sections of nuclei from different focal planes were acquired using a 488-nm argon laser source and a 63× oil immersion objective. The images were recorded using the attached CCD camera. The digitized images of the optical sections were stacked and reconstructed using the Leica software offline.
Transmission electron microscopy
The isolated nuclei pellets of cultured cells were suspended in 500 μL of 1× NEB buffer with various EDTA concentrations. The nuclei suspensions were prefixed using NEB with 2% freshly paraformaldehyde (PFA), and then postfixed with 2% osmium tetraoxide (OsO4) in 1× NEB at room temperature for 2 h. The fixed nuclei were dehydrated using a series of graded ethanol. The nuclei were embedded in Spurr's epoxy resin; then, the ultrathin sections were cut. The ultrathin sections were mounted on carbon-coated copper grids and observed in a JEOL 100CX TEM after double-positive staining with 1.5% lead citrate and 2% uranyl acetate for 15 min.
Microcalorimetry
Isolated nuclei from 800 mL of cultured cells (dry isolated nuclear pellet of ∼2 mg) were used for each microcalorimetry examination of isothermal titration calorimetry (ITC) with a normal cell (1.45 mL) and differential scanning microcalorimetry (Micro-DSC) with a 0.5-mL coined cell (Microcal Inc.). The data were fitted and analyzed using Origin software (OriginLab Corporation, Northampton, MA) supplied by the manufacturer. The nuclei suspension and all solutions for microcalorimetry were degassed at 25°C for 15 min before the experiments. For ITC, the DTT in the NEB was replaced with Tris(2-Carboxyethyl)phosphine hydrochloride because the presence of DTT in a solution could cause a large fluctuation in the baseline. For each ITC titration, 50 injections of 5 μL of 50 mM EDTA in NEB were titrated into the cell with a 180 s interval between each injection at 25°C. For the DSC experiments, the nuclei were equilibrated at 10°C for 2 h, and then scanned from 10°C to 130°C with a heating rate of 1°C·min−1.
Results
The nuclei of K. papilionacea in living cells show a strong birefringence, a distinctive feature of liquid crystals under a polarizing light microscope. Previous studies of dinoflagellates chromosomes focused on mounted whole cells or spread chromosomes (Herzog and Soyer, 1983; Chow et al., 2010). To study the condensation and decompaction of LCCs in an intact state, we performed our experiments in isolated nuclei, an environment similar to that of living cells. Accordingly, the karyotypes of LCCs in isolated nuclei differ from those observed in fixed whole cells or after processing by chromosome spreading. When isolating the nuclei, intact intranuclear chromosomes were preserved in a liquid crystalline state similar to that found in vivo (Fig. 1A–C). We were able to observe intact LCCs with an identical birefringence in isolated nuclei even after extended periods (days) of storage. Quantitative images show that intranuclear birefringent LCCs, which were initially in a condensed state, were distributed with optical symmetry in isolated nuclei.
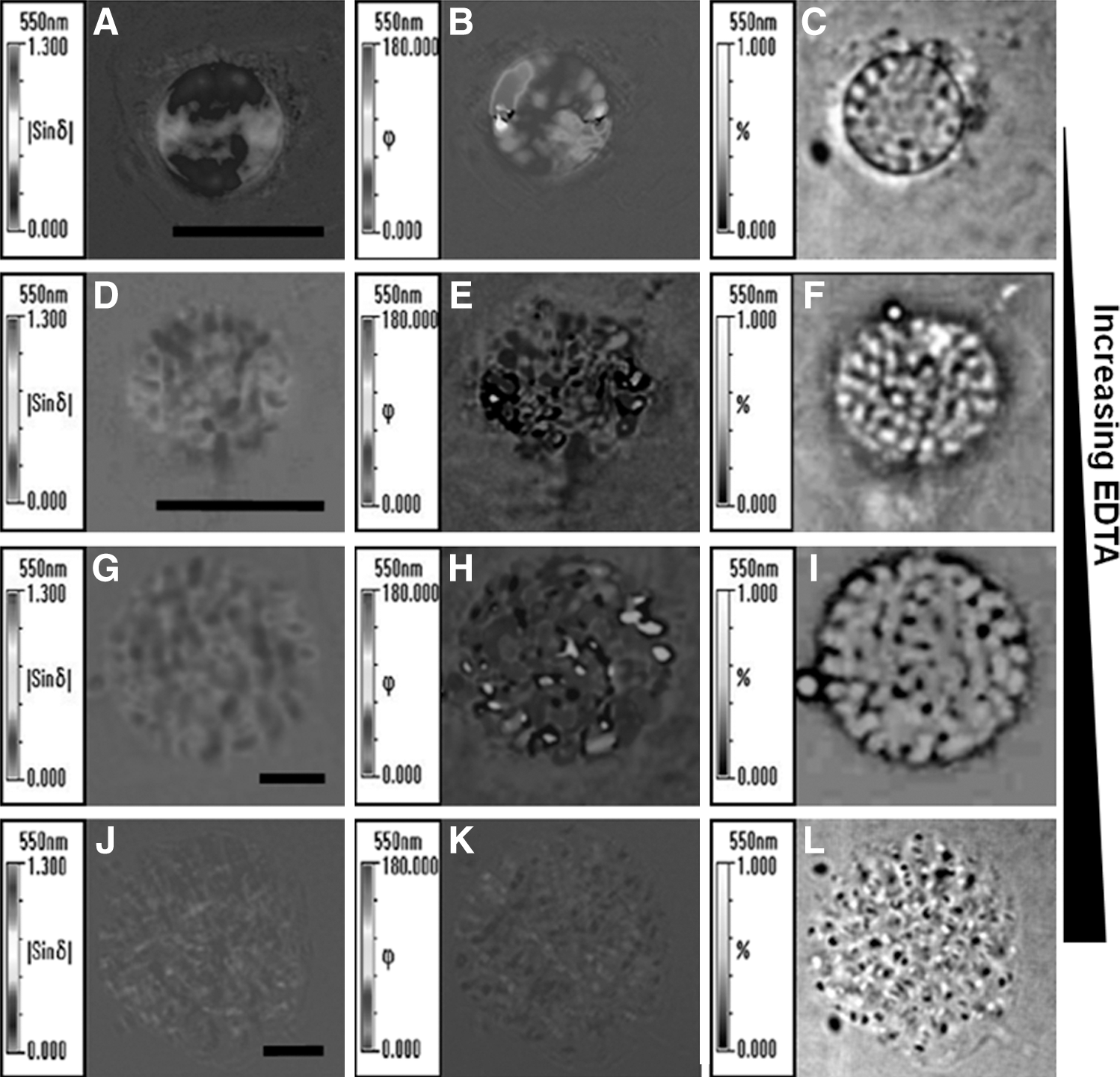
Quantitative mapping of the optical properties of isolated Karenia papilionacea nuclei at different decompaction states. The different decompaction states of nuclei were obtained using various ethylenediaminetetraacetic acid (EDTA) concentrations in the nuclei storage buffer.
We reasoned that increasing the EDTA concentration would result in the gradual extraction of the counterions from the LCCs, which would induce gradual decompaction of the LCCs. Interestingly, EDTA-treated nuclei had three levels of decompaction: I, II, and III states (Fig. 1D–L), corresponding to treatments with 2, 5, and 10 mM EDTA, respectively. The intranuclear LCCs adopted a disorderly orientation in the presence of 2 mM EDTA concentrations (Fig. 1E). The weakly birefringent intranuclear LCCs show a homogenous distribution in the decompaction II state (Fig. 1G). In the decompaction III state, the totally decompacted intranuclear LCCs lost birefringence and did not show any anisotropic orientation (Fig. 1J–L).
To reveal intranuclear 3D rearrangements in response to varied extranuclear EDTA concentrations, a 3D reconstruction was performed using the results from CLSM. The intranuclear chromatin was stained using Sytox Green, a fluorescent nucleic acid dye. A pair of orthogonal polarizers was used in the transmission channel to observe variations in the birefringence of the intranuclear LCCs as measured using ordinary polarizing microscopy. A typical reconstructed nucleus with various EDTA concentrations is shown in Figure 2. The condensed nucleus has a spherical conformation with a diameter of 8.3 μm (Fig. 2A, B). The intranuclear LCCs were highly compacted when there was a strong birefringence. However, the intranuclear chromosomes were indiscernible using CLSM. In the presence of 2 mM EDTA, the isolated nuclei were adopted to the decompaction I state with a diameter of 10.3 μm (Fig. 2C, D). From the CLSM images, we observed that the granular chromatins were packaged inside the nucleus with an estimated width of 240 nm and length of 900 nm. There were also numerous granular subunits along the decompaction chromosome. However, the constituent subunits of these granular chromatins cannot be observed using CLSM because of CLSM resolution limits. As the EDTA concentration increased to 5 mM, the nuclei transitioned to the decompaction II state with a diameter of 22 μm (Fig. 2E, F). The isolated nuclei lose their birefringence in the decompaction II state, but single chromosomes are still distinguishable in the image from the fluorescence channel (Fig. 2E). Each chromosome in the decompaction II state consists of 14 to 18 subunits with an average length of 7.3 μm and width of 1.4 μm. In the presence of 10 mM EDTA, the nuclei were totally decompacted and in the decompaction III state (Fig. 2G, H). The mean size of these nuclei is similar to the size of the nuclei in the decompaction II state (Fig. 2F, H). Compared with those in the decompaction II state, the completely decompacted chromosomes were expected to extend to a larger size. However, it would not be possible to accurately measure the dimensions of individual chromosomes because chromosomes in the completely decompacted state lack distinct features in both stained fluorescence and birefringence.
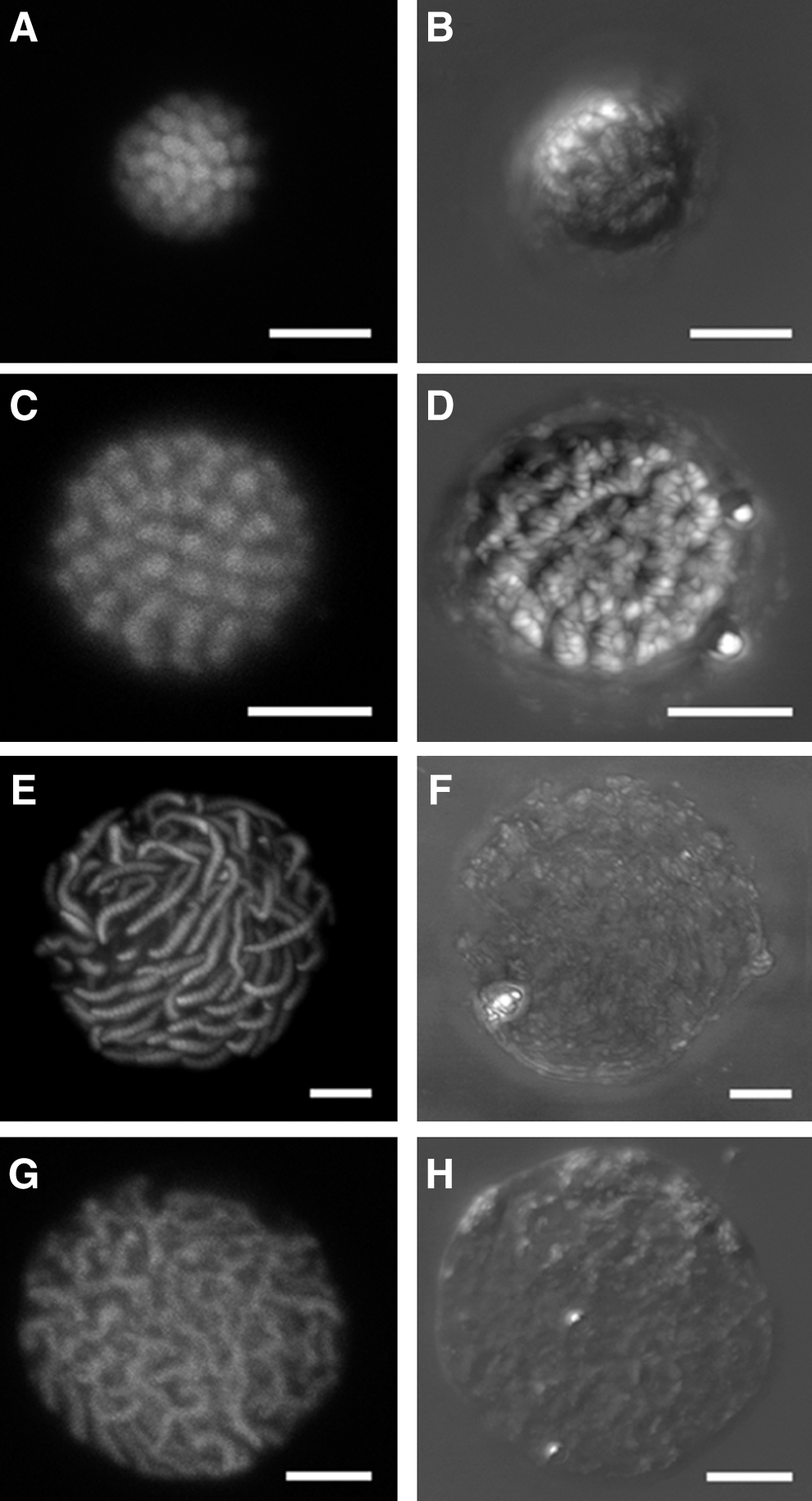
Confocal laser scanning microscopy images of isolated Karenia papilionacea nuclei in different decompaction states. The nuclei for confocal laser scanning microscopy (CLSM) observations were stained using Sytox Green. A pair of orthogonal polarizers in the CLSM transmission channel was used for observing the liquid crystalline behavior of the liquid crystalline chromosomes. The different decompaction states of the nuclei were obtained using various EDTA concentrations in the nuclei storage buffer.
To investigate the variations of the intranuclear structure in response to chromosomal decompaction, ultrathin sections of nuclei with different EDTA concentrations were observed using TEM (Fig. 3). As shown in Figure 3A, a condensed nucleus showed the characteristic patterns of cholesteric nested arch-like organization, which can be found in any published TEM images of dinoflagellate chromosomes (Costas and Goyanes, 2005; Alverca et al., 2007). The condensed chromosome was composed of cholesteric spiral chromatin filaments (Fig. 3B). Previous biochemical analysis has suggested that these chromatin filaments are organized by DNA and histone-like proteins (Costas and Goyanes, 2005). When measured perpendicular to the long axis of the chromosome, the diameter of a chromosome is approximately 1.1 μm (Fig. 3B). The arch-like bands consist of nested fibrous coils with a width of 150 nm. A series of these arched bands are stacked in parallel rows, which can be found using images taken at different orientation angles (compared to direction of the electronic beam) (Fig. 3C). A partly decompacted nucleus is shown in Figure 3D. The chromosomes were disassembled, although the nested arch-like bands remained. An average single chromosome is approximately 4.4-μm long and 1.1-μm wide and consists of 24 nested arch-like bands (Fig. 3E, F). The dense central core of the nuclear matrix was distributed in the chromosomes, which were stained darker than the peripheral chromatin fibers (Fig. 3G) (Minguez et al., 1994). In the completely decondensed decompaction III state, the chromosomes collapsed (Fig. 3H). Most of the peripheral chromatin filaments were lost, and a dense central scaffold-like core of the nuclear matrix remained (Fig. 3I). Secondary ion mass spectrometry (SIMS) has been used to quantitatively image the dinoflagellate chromosomes; the results identified the dense material of the core fibers as nuclear matrix, such as an insoluble protein and a low DNA content (Levi-Setti et al., 2008).
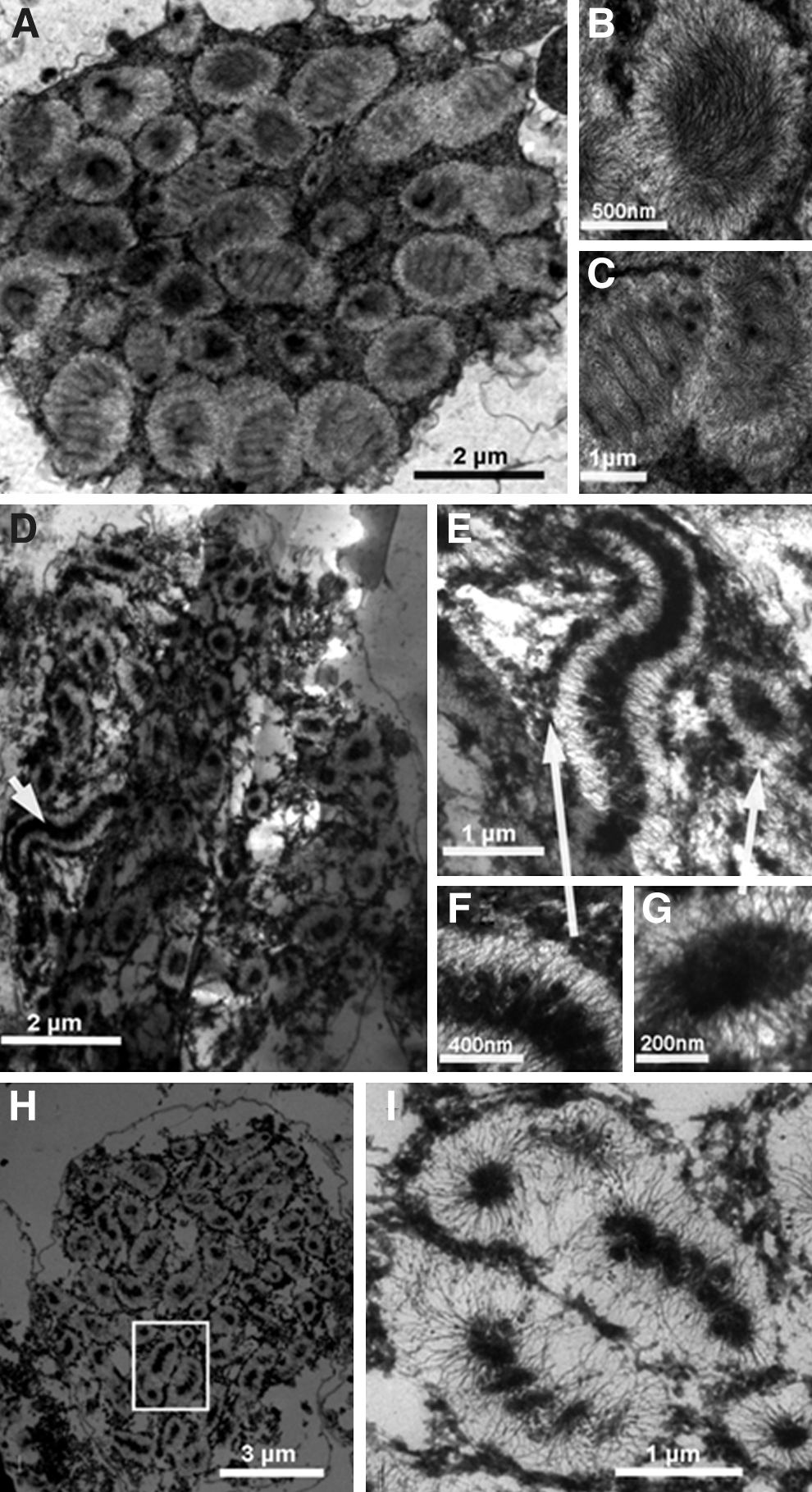
Transmission electron microscopy images of nuclear structures of isolated Karenia papilionacea nuclei in different decompaction states.
ITC has the advantage of a high sensitivity; thus, the thermodynamic processes involved in the decompaction of isolated nuclei in response to a series of continuous decreases in extranuclear Ca2+ concentration via EDTA were measured using ITC. Representative titration curves from applying EDTA to an isolated nuclei suspension are shown in Figure 4, which shows the enthalpy versus the injected volume of EDTA-containing NEB. The integrated data from titrating EDTA in isolated nuclei suspension is exothermic, indicating that titrations are accompanied by decreasing entropy. The amount of heat that generated and reached a maximum value as the concentration of EDTA continuously increased.
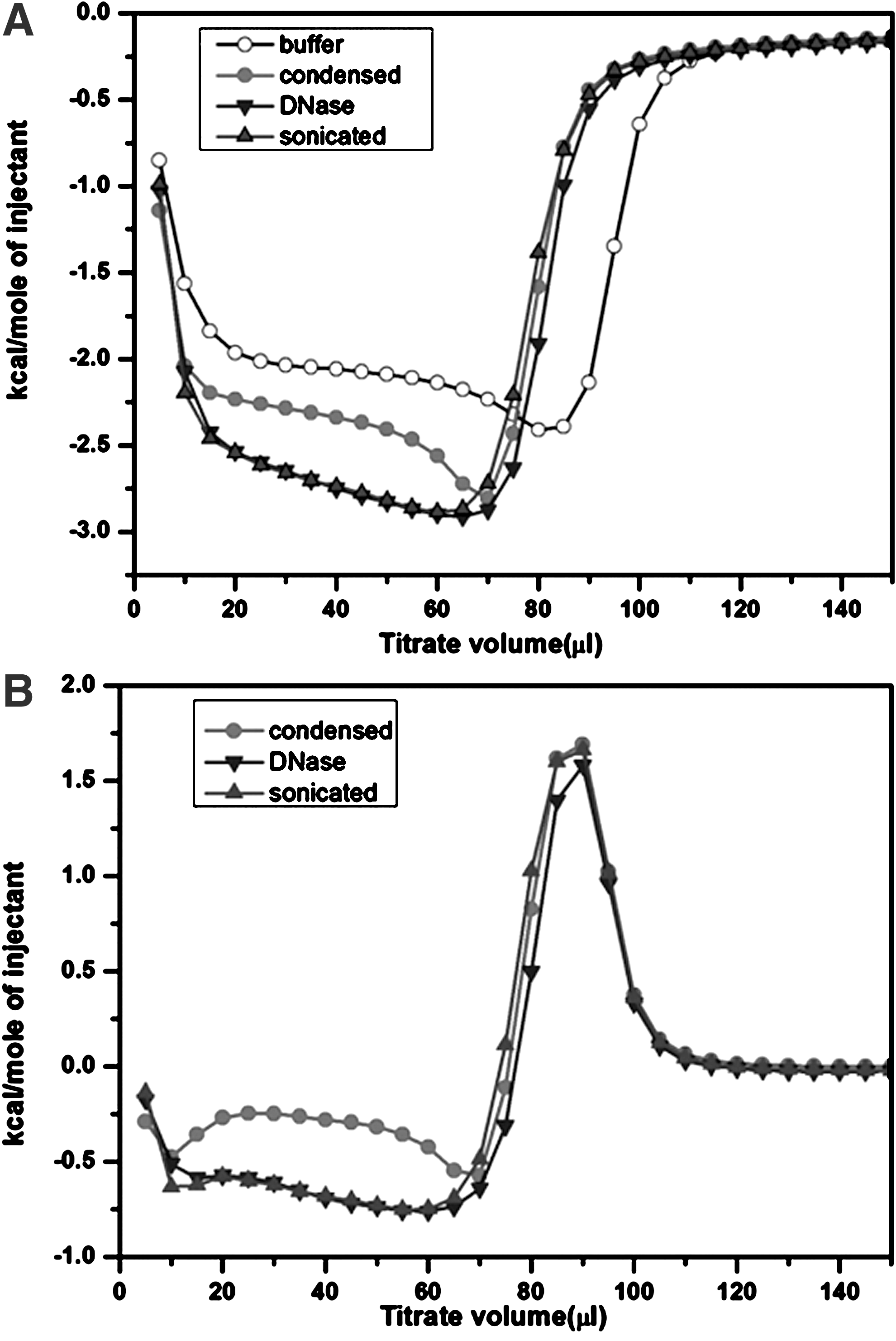
Representative isothermal titration calorimetry curves of an isolated Karenia papilionacea nuclei suspension. Nuclei extraction buffer containing 50 mM EDTA was titrated into the isolated nuclei suspension, which initially contained 1.5 mM Ca2+. The amount of injected solution for each titration point was 5 μL with 3 min between every two injections at 25°C.
When chromosomes become decompacted, many of the divalent cations that stabilize them may be released as a result of the collapse of the higher order organization of chromosomes. Sonication and DNase treatments are well-known disruptive techniques for preparing DNA fragments (Merchant and Rill, 1997; Spadiliero et al., 2002). Consequently, isolated nuclei were treated by ultrasonic disruption or DNase I digestion before ITC titrations. Compared with measurements at similar concentrations of an isolated nuclei suspension without any treatment, much more heat was generated from the ITC titration of treated nuclei. This finding indicates that a significant amount of the chromosome-stabilizing divalent cations, Ca2+ and Mg2+, were released from the fragmented chromosomes (Fig. 4A). The raw ITC curves were subtracted from a control curve of the buffer to display the net thermodynamic responses from isolated nuclei with added EDTA (Fig. 4B). The EDTA titration of Mg2+ and Ca2+, coexisting divalent cations, indicates that the binding is competitive and shows the two distinctive nonlinear stages reported for other systems (Fig. 4B) (Henzl et al., 2003). The initially exothermic stage is related to chelating Ca2+, and the subsequent endothermic stage is associated with chelating Mg2+ (Henzl et al., 2003).
The thermal stabilities of isolated eukaryotic nuclei were also examined using Micro-DSC. The thermal transitions of isolated K. papilionacea nuclei have four major endothermic transitions: 64°C (I), 81°C (II), 90°C (III), and 115°C (IV) (Fig. 5). Dinoflagellate nuclei are characterized by the absence of core histones and the presence of a large amount of DNA with a low ratio of basic histone-like proteins to DNA (Rae, 1973; Rizzo, 2003). We assumed that these four thermal transitions fit into the following categories: I, the liquid-crystalline-phase transition of LCCs; II, nucleosome-like domains; III, relaxed DNA; and IV, a higher order organization of chromosome. To verify this hypothesis, the isolated nuclei were treated with EDTA for control experiments (Fig. 5). The DSC profile of decompacted nuclei treated with 15 mM EDTA consists of three transitions (II, III, and IV) and lacks transition I (Fig. 5).
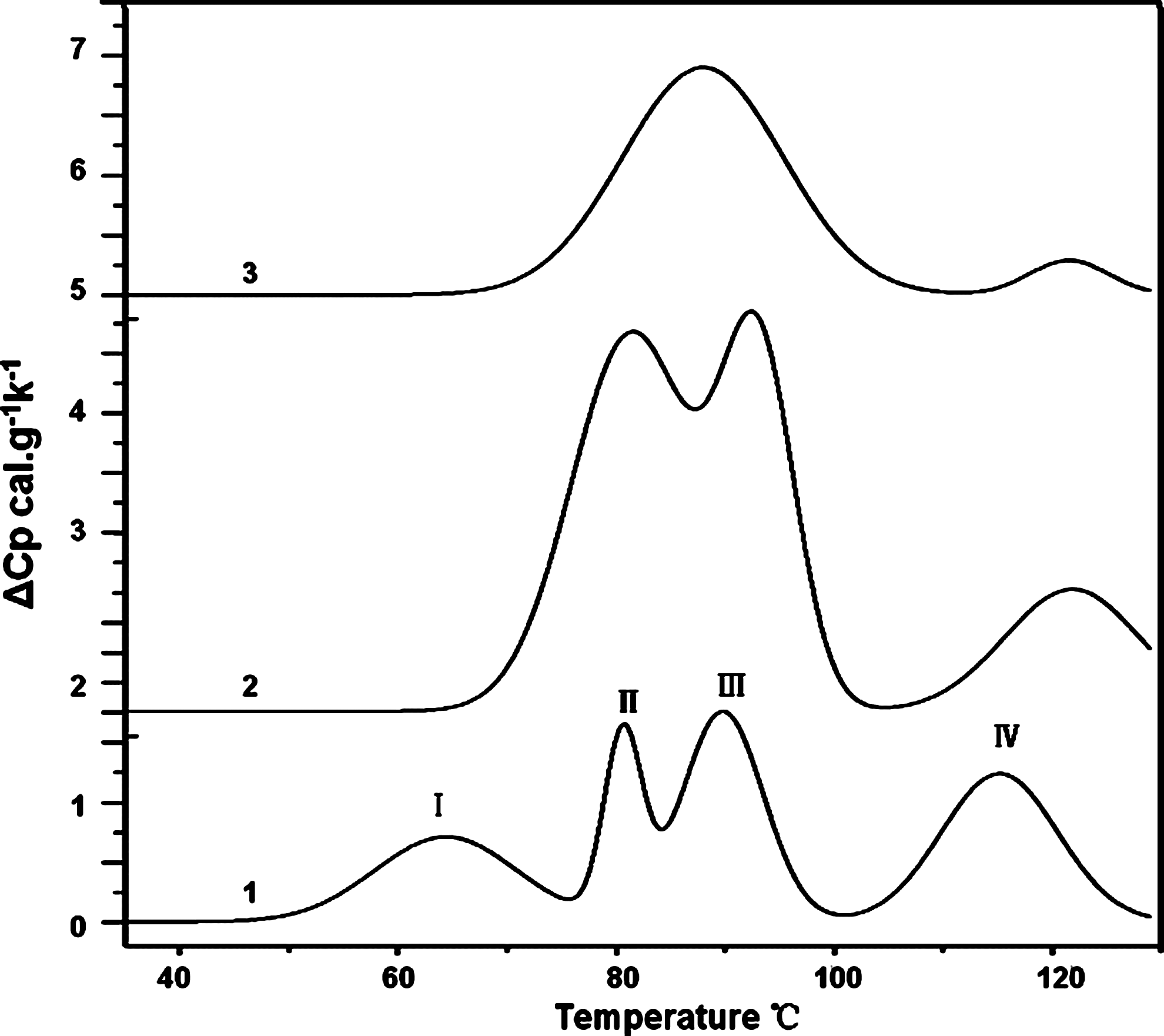
Representative differential scanning microcalorimetry profiles of an isolated Karenia papilionacea nuclei suspension. Curve 1, the isolated nuclei in a condensed state. Curve 2, the decompacted nuclei induced by the application of 6 mM EDTA before the DSC experiment. Curve 3, decompaction nuclei induced by the application of 15 mM EDTA before the DSC experiment. I, II, III, and IV in curve 1 represent the four major thermal transitions.
Discussion
Multiple independent techniques (morphology, ultrastructure, and thermodynamics) were used to reveal variations in the intranuclear chromosomes decompaction in response to changes in the extranuclear concentrations of divalent cations induced by EDTA chelation. The ordinary polarizing microscope has limited ability to distinguish birefringent patterns (Tixier et al., 2005). The spatial variations in the anisotropic birefringence of intranuclear LCCs were investigated using the Metripol birefringence imaging technique. The Metripol orientation angle (φ) is the extinction angle between extraordinary and ordinary light (Geday et al., 2000). In cholesteric LCCs, the orientation angle normally refers to the angle between the long axis of a chromosome and the direction in which the chromatin filament fibers are aligned. Therefore, the colorful patterns in the Metripol orientation micrograph can be considered the orientation of intranuclear LCCs (Fig. 1).
Rearrangements of birefringent chromosomes throughout decompaction processes induced by EDTA chelation were also investigated using CLSM (Fig. 2). The partly decompacted nucleus had a large granular chromatin that was tightly compacted inside the intranuclear space in the decompaction I state. When comparing the transmission and fluorescence images, which were simultaneously recorded, chromatin was identified as the main component of these particles. The intranuclear LCCs in a condensed state swelled with the addition of EDTA, and there was no visible birefringence in the decompaction II state (Figs. 1G and 2F). The nuclear volume of a decompacted nucleus can swell to almost three times that of the initially condensed state.
As shown in Figure 4, the dynamic ITC titration processes are interpreted as follows: EDTA initially bound extranuclear Ca2+ in the storage buffer of isolated K. papilionacea nuclei. The extranuclear ionic environment changed and, subsequently, allowed intranuclear Ca2+ to pass through the nuclear envelope to reach a new ionic equilibrium. The extraction induced the nucleus to extend to a larger volume, which provided a larger volume for the chromatin package. The continuous increase in the nuclear volume promoted extraction of divalent cations of Ca2+ and Mg2+ from chromosomes and spontaneously caused decompaction of LCCs. The series of decompaction states were achieved by a continuous addition of EDTA in the nuclei suspension. The addition of EDTA could affect the chromosome structure through various mechanisms, such as removal of the histone-like-protein (or any other chromosomal proteins) from DNA or disruption of the nuclear membrane because of depolymerization of inner nuclear membrane proteins. However, it is possible that these proteins require divalent ion to bind to DNA. Moreover, chromosomal decompacted nuclei kept the circularly morphology and the dynamic decompaction processes occur in a nonlinear manner (Fig. 2). These findings imply that the nuclear envelope, a diffusion barrier, remains intact because control titrations using broken nuclei have much smoother curves (Fig. 4B) (Finan et al., 2009). Therefore, the extraction of divalent cations from chromosomes may contribute to this nonlinear aspect. The competitive binding effects of coexisting Ca2+ and Mg2+ must also be considered (Fig. 4B). The titration of divalent cations using EDTA when Ca2+ and Mg2+ coexist shows two distinctive stages (Henzl et al., 2003). Therefore, intrinsic chromosome decompaction may occur during the first stage, in which the binding of EDTA to Ca2+ induces extraction, stabilizing the cations on the chromosome.
The reversibility of LCCs compaction/decompaction was checked by add-back of Mg ions to EDTA-treated nuclei. As shown in Supplementary Figures S1 and S2 (Supplementary Data are available online at
Micro-DSC is a highly sensitive microcalorimetry technique for studying the thermal stabilities of biological macromolecules (Lepock, 2005). DSC measurements of isolated nuclei of nucleosome-based eukaryotes revealed four main distinctive transitions; each transition corresponds to the unfolding of a specific level of organization (Spadiliero et al., 2002). The isolated nuclei of general eukaryotes are characterized with four main endothermic transitions at the following transition temperatures (Tm): I 60°C, II 76°C, III 88°C, and IV 105°C, respectively. However, different cellular systems have a slightly different temperature point at each transition, depending on the GC content and their higher order structures (Cardellini et al., 2000; Lepock et al., 2001). These distinguishable transitions have been assigned as follows: I, thermolabile nuclear components, such as the nuclear matrix; II, collapsed nucleosome; III, the main transition, separation of the DNA double helix released from chromatin; and IV, nucleosome-assembled higher order organization (Cardellini et al., 2000; Lepock et al., 2001).
In this study, the isolated K. papilionacea nuclei in condensed states, which are devoid of core histones and canonical nucleosomes, were revealed to also have four major thermal transitions (Fig. 5). Transition I has a wider (60°C–70°C) peak in LCC-based nuclei than in nucleosome-based nuclei; thus, the contribution of transition I to the overall heat capacitance for the disassembly process is greater in LCC-based nuclei. With the addition of EDTA, transition I will disappear first, (Fig. 5), corresponding to the disappearance of birefringence with EDTA treatment (Figs. 1G and 2F). Because birefringence is related to orientation anisotropy, this finding suggests that transition I is the heat capacitance involved in the phase transition during the assembly of LCCs. Cholesteric liquid crystalline DNA has a thermal phase transition near 70°C, which has been characterized using thermal polarizing microscopy and DSC (Olesiak et al., 2009). Therefore, transition I of the condensed K. papilionacea nuclei represents overlapping transitions of thermolabile nuclear components and LCCs. The nuclear matrix most likely denatured at the lower temperature, 55°C. Thus, we can assign the transition at 64°C to phase transition of LCCs.
The relative normalized intensity of transition IV decreases proportionally to the extent of the disorganization of the higher order chromatin structure after treating the isolated nuclei with excessive amounts of EDTA. Therefore, transitions of II and III are overlapping phase transitions and associated with the relaxation and denaturation of DNA. The microcalorimetry results are consistent with the existence of the higher order organization in dinoflagellates chromosomes.
The volume and morphology of isolated nuclei are inversely and nonlinearly proportional to the extranuclear osmolarity (Finan et al., 2009; Finan and Guilak, 2010). Increases in the length of chromosomes and the nuclear volume can be positively correlated with the level of nuclear decompaction that occurs in response to extranuclear physiological changes in ionic conditions (Bojanowski and Ingber, 1998; Finan and Guilak, 2010). Our results demonstrate the nonlinear nature of the decompaction of LCCs in response to decreases in the extranuclear concentration of divalent cations (Fig. 4B). The EDTA chelation of divalent cations induced the extraction of Ca2+ and Mg2+, counterions required for chromosomal maintenance. The increased nuclear volume may influence the transition of LCCs to decompaction states as a result of excluded volume effects. Extending the nuclear volume and extracting chromosomal maintenance divalent counterions are interdependent actions that regulate the nuclear decompaction processes of K. papilionacea.
As shown in TEM images, these higher order nuclear organizations may be correlated to cholesteric arch-like bands, which are assembled by nested coils of fibrous chromatin filaments (Fig. 3). The TEM images of ultrastructure sections showed that the condensed chromosomes were compacted in a typical cholesteric liquid crystalline organization (Fig. 3A–C). Previous results demonstrated that the birefringence of LCCs relates to their structure and organization (Chow et al., 2010). There are neither core histones for structural maintenance nor linker histones. Thus, liquid crystalline structures and LCCs may be assembled from nested coils of fibrous chromatin filaments. Moreover, positively charged divalent counterions play significant roles in modulating liquid crystalline phases for compacting chromosomes of dinoflagellates.
Thermodynamically, the exothermic decompaction processes indicate decreasing entropies (Fig. 4B). In the crowded intranuclear environment, long molecular chains frequently form helices because of crowding and entropic forces (Snir and Kamien, 2005; Hancock, 2007). Therefore, the entropy-driven forces and electrostatic interactions from divalent counterions and histone-like proteins may cooperatively modulate the formation of distinctive liquid crystalline and birefringent chromosomes in dinoflagellates.
Conclusion
In summary, studying the decompaction processes of intranuclear LCCs in isolated nuclei identified three states in the decompaction of the LCCs using EDTA concentrations of 2 mM, 5 mM, and 10 mM. The dynamic processes of nuclear decompaction were found to occur in a nonlinear manner. The thermal stability assessment of condensed and EDTA-treated isolated nuclei revealed that the thermal transition at 64°C corresponded to a liquid-crystalline-phase transition of LCCs. Our results suggest that nested coils of fibrous chromatin filaments are responsible for assembling the liquid crystalline and birefringent structure and that positively charged divalent counterions play significant roles in modulating liquid crystalline phases for the compaction of chromosomes of dinoflagellates. However, in living cells, there are enzymes and cofactors that can facilitate the chromatin condensation in a complex manner in addition to the physical biological processes. It would be of great interest to investigate the possible enzymes involved in the regulation of higher order structures of LCCs.
Footnotes
Acknowledgments
The present work was partly supported by CERG HKUST6421/06M, the National Natural Science Foundation of China (Nos. 41102213, 41130746, and 41272371), SWUST Scientific Research Foundation for Doctoral Scholars (10zx7128), and the opening Project of Key Laboratory of Solid Waste Treatment and Resource Recycle, Ministry of Education (11zxgk05).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.