
Abstract
Despite inherent limitations, the ease and rapidity of their use make transiently expressed reporter gene assays the most frequently used techniques for analyzing promoters and transcriptional regulators. The results of transient reporter gene assays are generally accepted to reflect transcriptional processes correctly, though these assays study regulatory sequences outside of the chromosomal environment and draw conclusions on transcription based on enzyme activity determination. For transient reporter gene assays, often more than one promoter is introduced into one cell. In addition to the one driving the primary reporter gene expression, a further one might serve to ensure the production of an internal control second reporter or/and a trans-acting factor. We demonstrate here by various examples that interference between physically unlinked promoters can profoundly affect reporter expression. Results of reporter gene assays performed by combinations of the cytomegalovirus promoter and various other promoter constructs (human immunodeficiency virus [HIV], Human T-cell Leukemia Virus Type I (HTLV-I), NF-κB-responsive, and p53-responsive) and trans-activator factors (HIV-Tat and p53) in different host cell lines (U2OS, HeLa, and L929) prove that interference between active transcription units can modify transcription responses dramatically. Since the interference depends on the promoters used, on the amount of transfected DNA, on the host cells, and on other factors, extra caution is required in interpreting results of transient reporter gene assays.
Introduction
Materials and Methods
Cell lines, media, culture conditions
U2OS, HeLa, and L929 cells were maintained at 37°C in humidified atmosphere of 5% CO2 and 95% air in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 1 mM
Plasmid constructs
pGL2-MDM2-Luc (Bereczki et al., 2008) and pCMV-Luc (Tombacz et al., 2009) plasmids were previously described. pHC624 is a high-copy-number derivate of pBR322 (Boros et al., 1984). pcDNA3 and pEGFP-N3 plasmids were purchased from Invitrogen and Clontech company, respectively. pHIV-Luc, pHTLV-1-Luc, pNF-κB-Luc, pSV-Tat reporter, and trans-activator-expressing plasmids were provided generously by the Chou-Zen Giam (USUH) and Jeang Kuan-Teh (NIH/NIAID) laboratories.
Transfection and luciferase activity measurement
U2OS, HeLa, and L929 cells were seeded in six-well plates at 2.5×105 cells/well in supplemented DMEM 24 h before transfection.
Plasmids were transfected into the cells using ExGene 500 or Turbofect reagent (Fermentas) according to the instructions of the manufacturer. Reporter constructs, trans-activator producing, and other constructs were added in different amounts as indicated in micrograms in figure legends. After 24 h incubation, the medium was removed, and the cells were washed and harvested. Cells were resuspended in 1× lysis buffer (Promega, Cell Culture Lysis Reagent 5×) and incubated on ice for 30 min. Next, the supernatants were collected in a centrifuge (at 13000 rpm at 4°C for 5 min), and used to determine protein concentration by the Bradford method and perform luciferase enzyme assay. Luciferase activity in the samples was determined using a Promega luciferase assay kit and Orion L Microplate Luminometer (Berthold Detection System, Simplicity 4.2 software). Transfections were performed in at least three repeats, and cell extracts generated from each transfection were used only once to determine reporter gene activity. Results are presented as mean±SEM of the results of three or more independent experiments. Luciferase activity values are expressed in percentage of a reference sample as indicated in the figure legends. Statistical analysis was done using SPSS for Windows version 15.0. Statistical significance was determined using the Mann–Whitney U nonparametric test of significance with a p<0.05 considered statistically significant.
Results
The CMV promoter interferes with human immunodeficiency virus-Tat and HTLV-I transactivation
The human immunodeficiency virus (HIV) promoter is one of the most (if not the most) extensively studied eukaryotic promoter. The HIV promoter is activated by the viral trans-activator protein Tat, which increases production of HIV genomic and mRNA by orders of magnitude. In recent years, the structure of HIV promoter and the mechanism of HIV promoter activation by Tat have been elucidated in details (Brigati et al., 2003). Results on transient expression of reporter genes introduced into cells by transfection of suitable plasmid constructs contributed significantly to the dissection of the HIV promoter. Moreover, this approach is still used to identify Tat targets and factors that might have an effect on the HIV promoter (recent examples, Gibellini et al., 2010; Deshmane et al., 2011; Jeong et al., 2012; Narasipura et al., 2012). If a luciferase-coding sequence under the control of the HIV promoter is introduced into U2OS cells, it produces a well-detectable reporter gene activity that significantly exceeds the background activity (Fig. 1A, B columns 1 and 2.). Coexpression of the HIV transactivator Tat in the same cells (under the control of the SV40 promoter in the example shown) further increases the HIV promoter-driven reporter gene activity (Fig. 1B, column 3.). Cotransfection with the HIV promoter-luc and SV40-Tat constructs, a further reporter plasmid, which carries the CMV promoter, and enhanced green fluorescent protein (EGFP)-coding region has a negative effect on the luciferase expression. Cotransfected CMV-EGFP dose dependently reduces the HIV promoter-driven reporter gene transcription. In the presence of the cotransfected second reporter, the luciferase activity drops well below the nontransactivated basal activity (Fig. 1A column 3., 1B columns 4–6.)
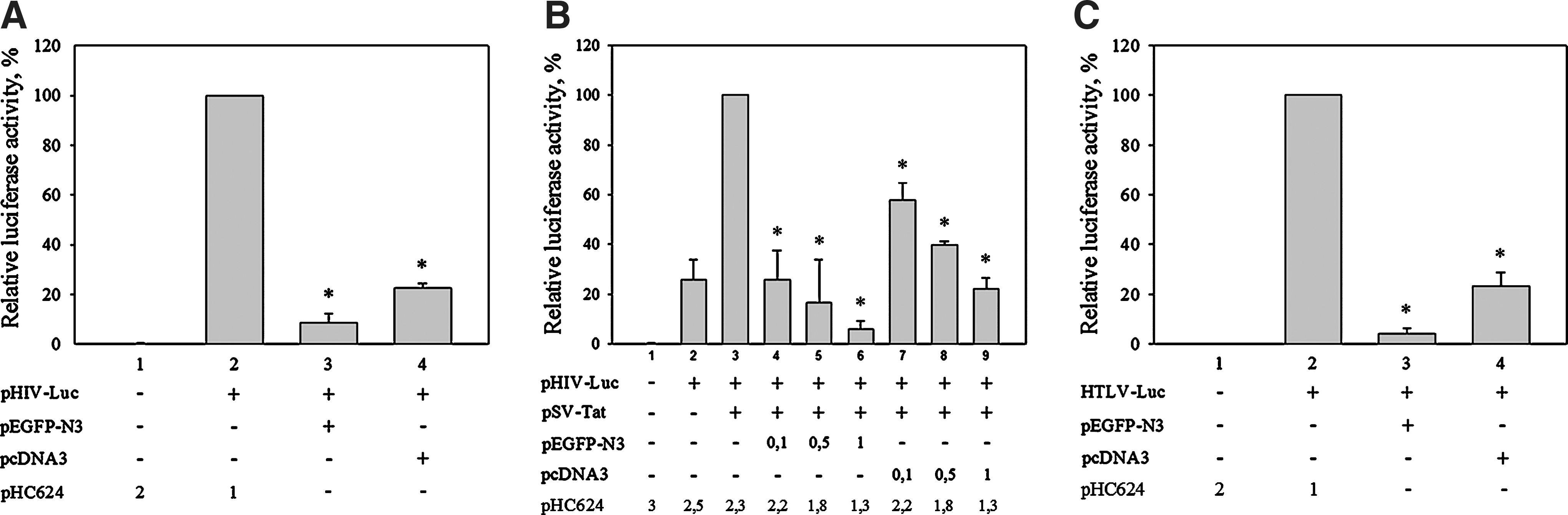
The presence of cytomegalovirus (CMV) promoter interferes with HIV and HTLV-1 promoter-driven transcription.
To exclude the possibility that the reduced reporter gene activity resulted from a toxic effect of EGFP (Liu et al., 1999), we cotransfected an empty vector pcDNA, which carries a CMV promoter without inserted open reading frame, therefore representing a short transcription unit (Fig. 1A column 4; Fig. 1B columns 7–9.). This too, resulted in a less strong, but very well-detectable decrease in luciferase activity.
The strong negative effect of the CMV promoter on HIV promoter-directed luciferase expression raises several questions; among them, most importantly, whether this effect is related to the unusual mechanism of Tat transcriptional regulation, or is a universal effect. To address this question, we tested whether the Human T-cell Leukemia Virus (HTLV) promoter was also affected in the presence of the CMV promoter. Transfection of an HTLV-1-promoter–luciferase reporter plasmid caused increased luciferase activity compared to the transfection control (Fig. 1C, columns 1 and 2.). Cotransfection of an EGFP producer plasmid or a CMV promoter containing empty pcDNA vector decreased the luciferase activity dramatically (Fig. 1C, columns 3 and 4.). Thus, these data indicate that the inhibitory effect of the CMV promoter on that of HIV is not promoter specific and is mediated via factors involved in the transcription of both viral promoters.
The CMV promoter inhibits luciferase expression from an NF-κB-responsive promoter
A shared feature of the HIV and HTLV-I promoters is that both contain copies of the NF-κB-response element. Since higher CMV promoter amounts reduced HIV- and HTLV-I LTR-driven luciferase activities below the basal level, we tested whether an NF-κB-responsive promoter–luciferase expression was affected by the cotransfection of the CMV promoter. As shown in Figure 2A, cotransfection of pNF-κB-luc with increasing amounts of either a reporter plasmid in which the green fluorescent protein is produced under the control of the CMV (pEGFP-N3) or CMV promoter-containing empty vector (pcDNA3) resulted in dose-dependent reduction of luciferase expression. Cotransfection of the plasmid producing EGFP has a stronger negative effect than cotransfection of comparable amounts of empty vector, suggesting that transcription over a longer unit might enhance the observed effect.
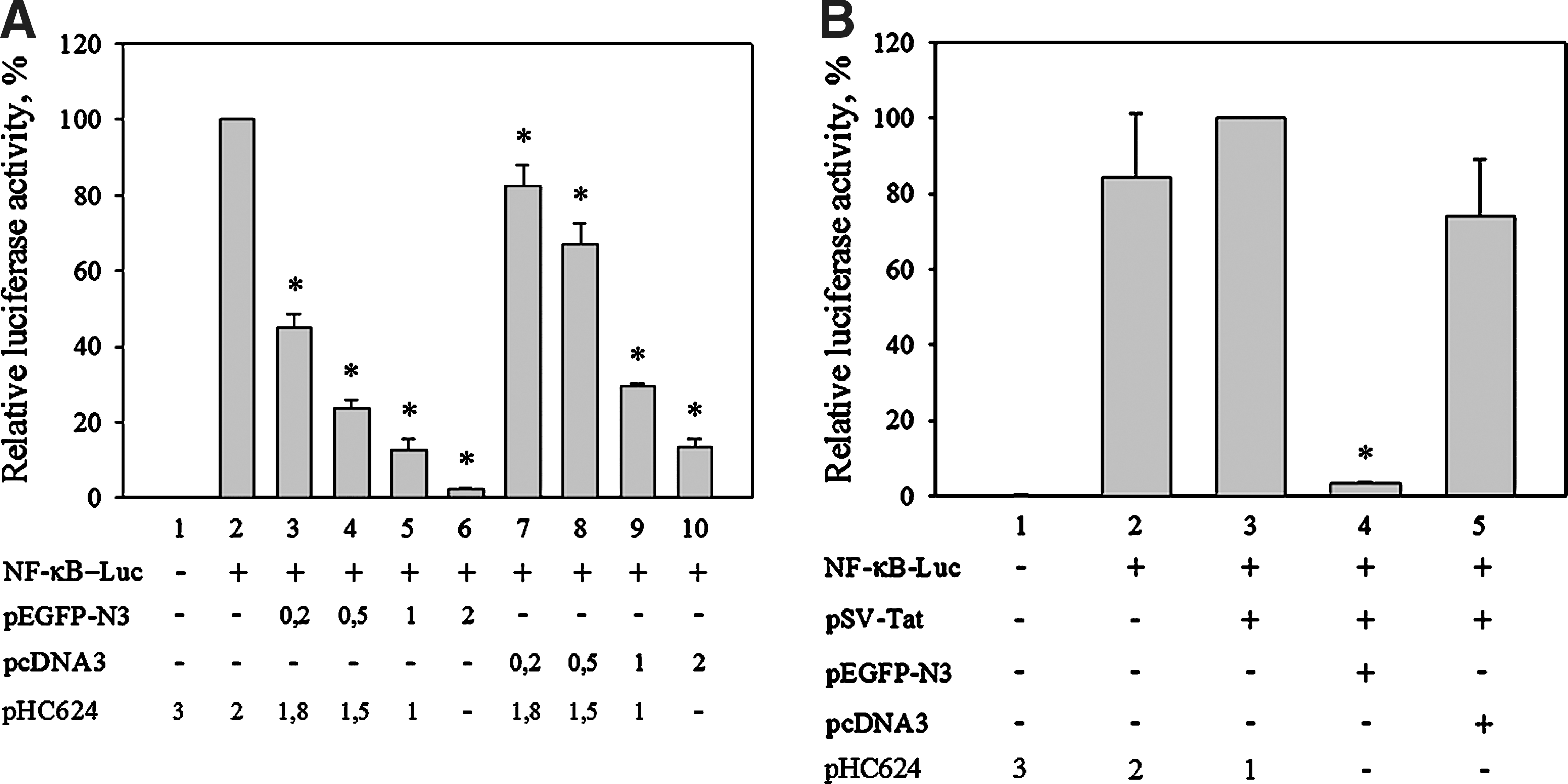
The presence of CMV promoter or CMV promoter-driven transcription unit decreases reporter gene expression from an NF-κB-response element-containing promoter.
Coexpression of the transactivator Tat protein increases the NF-κB-driven luciferase activity
Cotransfecting the cells in addition to the NF-κB-luc and SV40-Tat constructs with a further reporter plasmid that carries the CMV promoter and the EGFP-coding region has a strong negative effect on the luciferase expression (Fig. 2B, column 4.). Cotransfection of the NF-κB-luc and SV40-Tat construct with empty pcDNA3 vector resulted in less strong decrease in the reporter activity (Fig. 2B, column 5.).
Promoter competition is universal and should be taken into account in interpreting reporter gene assays
The observed interference between the CMV promoter and three promoters containing NF-κB-responsive elements prompted us to ask whether the CMV promoter interfered with transcription activation by other factors as well. To find an answer to this question, we determined the activation of the MDM2 promoter by p53 in the absence and presence of CMV promoter-containing plasmids (Fig. 3). Similarly to that observed in the case of the NF-κB-responsive promoters, the presence of the CMV promoter inhibited the luciferase production from the MDM2 promoter in a dose-dependent manner. Again, the effect was observable with very small amounts of cotransfected CMV plasmid and was stronger in the presence of a CMV-EGFP construct than in the presence of the CMV promoter-containing empty vector (compare columns 3–6 with 7–10 in Fig. 3). Furthermore, the presence of the CMV promoter reduced the MDM2 promoter-directed luciferase reporter activity similarly in U2OS and (as well as in) L929 (data not shown) cells.
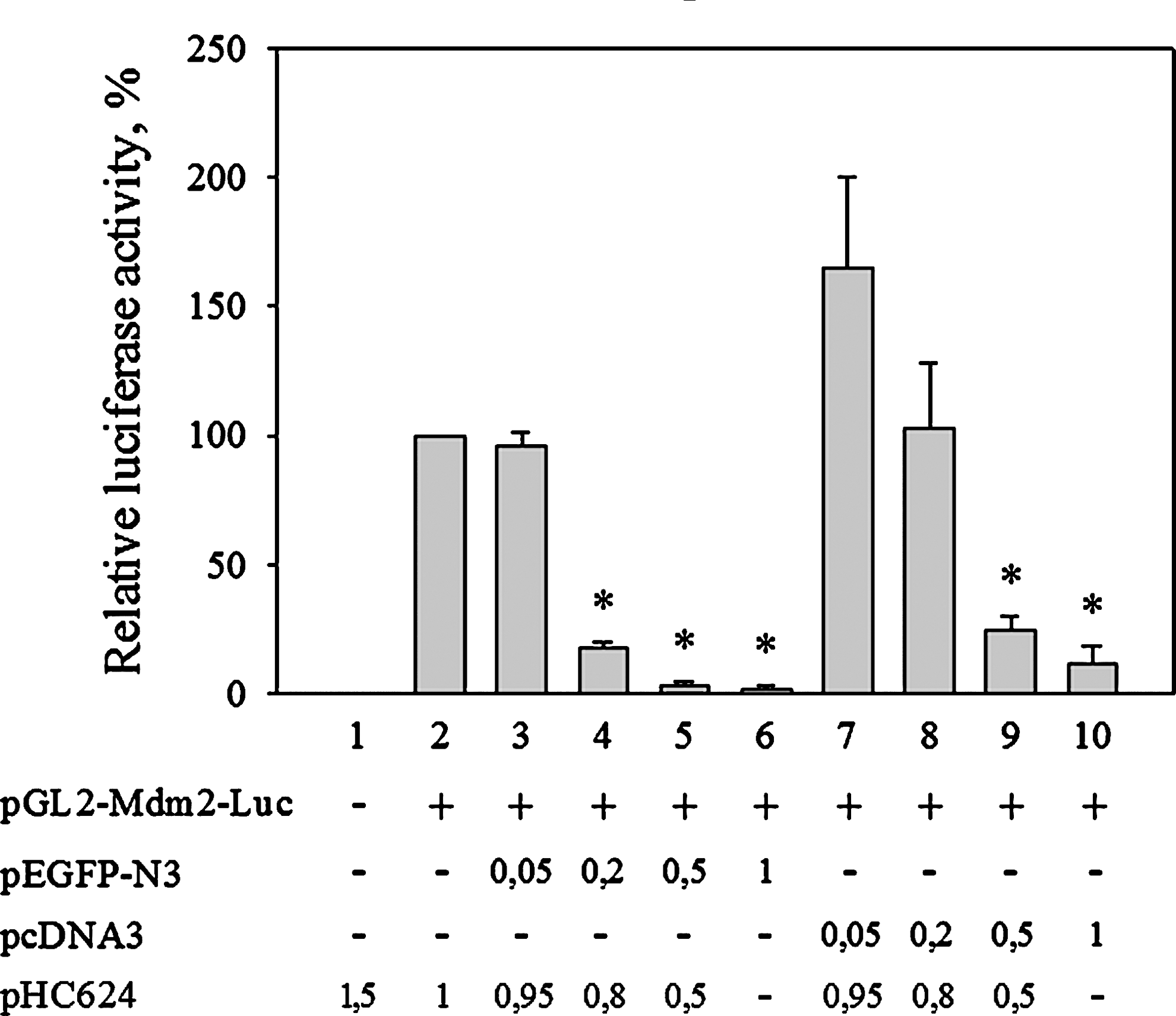
The presence of CMV promoter or CMV promoter-driven transcription unit decreases reporter gene expression from a p53-activated promoter. U2OS cells were transfected with 0.5 μg pGL2-MDM2-Luc, a p53–responsive reporter plasmid and increasing amounts of CMV promoter-containing plasmids: pcDNA3 or pEGFP-N3 as indicated. Luciferase activities were measured 24 h after transfection and are expressed as percent of activity determined in the activated sample (column 2). (*p<0.05, Mann–Whitney test).
The finding that the CMV promoter interfered with unrelated promoters suggested a competition for limiting general factors of transcription. This suggested a competition between two CMV promoter constructs as well. Indeed, when we cotransfected HeLa cells with a CMV-luc construct with either an EGFP-containing vector or a vector that contained only the CMV promoter, in both cases, we observed strong decrease in luciferase activity (Fig. 4). On the other hand, promoter-less vectors had no effect on reporter gene expression as had no effect of the cotransfected pHC624 plasmid, which we used in each experiment to normalize the amount of transfected DNA.
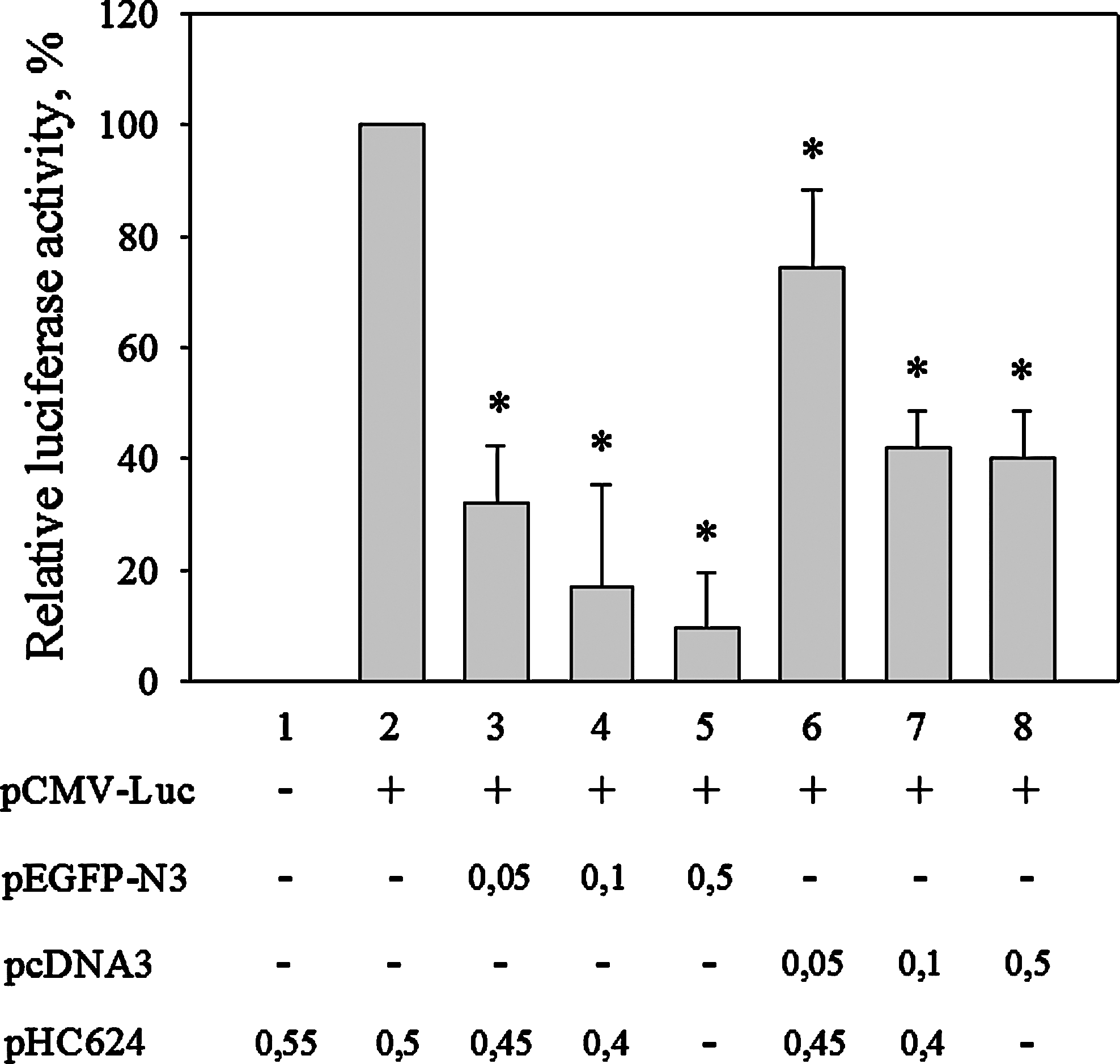
Small quantities of CMV promoters cause competition for limiting transcription factor. About 0.05 μg of CMV promoter-driven luciferase reporter plasmid (pCMV-Luc) was transfected into HeLa cells with increasing amounts of pcDNA3 or pEGFP-N3 CMV containing plasmids as indicated. Luciferase activities were measured 24 h after transfection and are expressed as percent of basal activity determined (column 2). (*p<0.05, Mann–Whitney test).
Discussion
During the course of studying transcriptional regulators, we observed negative effects on reporter gene activities by the cotransfection of a transcription unit containing a strong promoter. Strikingly, small amounts of an empty vector containing the CMV promoter and even less amounts of plasmids containing a transcription unit under the control of the CMV promoter drastically reduced reporter gene activities synthesized from several-fold higher amounts of reporter plasmids under the control of HIV-Tat, HTLV-I, NF-κB, and p53-activated promoters. The negative effect of the CMV promoter on different other promoters is observable in different cell lines and suggests competition for a general component of the transcriptional machinery. The limiting availability of some factors is also indicated by the observation that the expression of luciferase from a very small amount of transfected CMV–luciferase construct can be inhibited by the same promoter. Since the inhibition is more effective with the cotransfection of a longer transcription unit, one might assume that the limiting factor is a component of the transcription machinery that plays a role during elongation. A negative effect of the produced EGFP protein itself, however, cannot be excluded either.
The effect we describe here is probably not mechanistically different from the known transcriptional squelching. However, squelching was described as titration of essential transcription factors by the abundance of an overexpressed transcriptional activation domain, a trans-acting regulator (Prywes and Zhu, 1992; Cahill et al., 1994). We demonstrate here the titration effect of a strong cis-acting element. The recognition of this possibility might gain significance in the interpretation of reporter assay results when the expression of a trans-acting factor or an internal control reporter is driven by a strong promoter such as CMV. The CMV promoter is in fact a favorite element of several vectors and is indeed frequently used in transient reporter assays. In light of the data presented here, the effect of CMV or similar strong promoters on the transcriptional machinery should be taken into account with careful controls.
Footnotes
Acknowledgments
We thank for the plasmid constructs provided by CZ Giam, KJ Teang, E. Balint. The technical help with transfections and luciferase assays of K. Okros is greatly appreciated.
A.S. was supported by the TAMOP program (TAMOP-4.2.2/B-10/1-2010-0012).
Disclosure Statement
The authors have no conflicts of interest to declare.