
Abstract
MicroRNAs (miRNAs) are a class of regulatory small RNAs that have fundamentally transformed our understanding of how gene networks are regulated representing one of the most exciting areas of the modern cardiology research. Among all known miRNAs, miR-208a is one of the most important heart-enriched miRNA playing a crucial role in the heart health and disease. miR-208a is a member of a miRNA family that also included miR-208b and is encoded by an intronic region of the Myh6 gene. Within the heart, miR-208a and miR-208b are involved in the regulation of the myosin heavy chain isoformswitch during development and in pathophysiological conditions. miR-208a is sufficient to induce arrhythmias, cardiac remodeling, and to regulate the expression of hypertrophy pathway components and the cardiac conduction system. Recently, the identification of miR-208a in the bloodstream has led to a great clinical interest to use this molecule as a potential noninvasive biomarker of myocardial injuries.
Introduction
In the last two decades, one family of regulatory ncRNAs called microRNAs (miRNAs), because of their short length (∼22 nucleotides), has fundamentally transformed our understanding of how gene networks are regulated representing one of the most exciting areas of the modern cardiology research. Since the discovery, the number of known miRNAs has expanded substantially. Today, more than 1500 different miRNAs have been identified in the human genome (
Biology and Role of miR-208a in the Heart
miR-208 is a highly conserved miRNA family constituted of two members, miR-208a and miR-208b, which have very similar nucleotide sequences (Fig. 1;
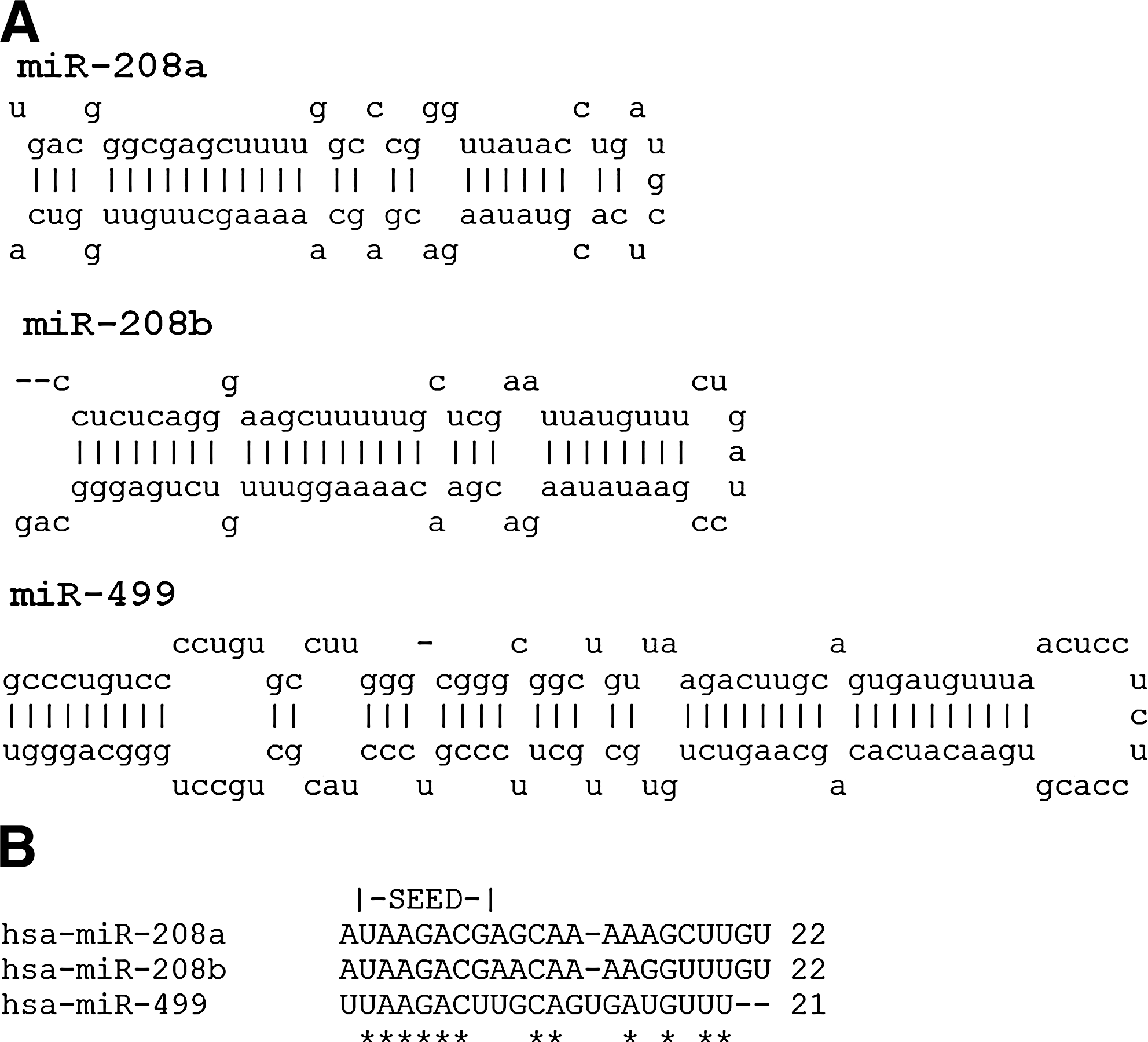
The efficiency of myocardial contractility is largely dependent on the contractile protein Myosin Heavy Chain (MHC). The MHC is the major contractile protein of the cardiac muscle and represented by two MHC isoforms: α-MHC (fast myosin) and β-MHC (slow myosin), which are encoded by Myh6 and Myh7 genes, respectively (Weiss and Leinwand, 1996). However, there are more ancient myosin genes, such as Myh7b. Although Myh7b is closely related to α-MHC and β-MHC, it does not generate a new myosin isoform with related or novel physiological function (Bell et al., 2010). The proportion of MHC isoforms may vary according with species and developmental stage. In adult mice, α-MHC is the predominant isoform, while in adult humans and larger mammals, β-MHC is the predominant cardiac MHC isoform. Some authors showed that Myh6 and Myh7 (genes encoding α-MHC and β-MHC, respectively), are differentially expressed in the adult heart, paralleling the expression of miR-208a and miR-208b, respectively (van Rooij et al., 2007; Callis et al., 2009; van Rooij et al., 2009). In rodents, miR-208b seems to be expressed largely in fetal hearts, reducing its expression levels until becoming adult. In contrast, miR-208a has lower expression levels during heart development, becoming predominant in adulthood. The switch from fetal miR-208b to miR-208a expression occurs shortly after birth indicating that if miR-208a and miR-208b target the same mRNAs, they do so at different developmental stages (Callis et al., 2009). Apparently, miR-208b and miR-499 are functionally redundant, and play a dominant role in the specification of muscle fiber identity by activating slow and repressing fast myofiber gene programs (van Rooij et al., 2009).
The deletion of miR-208a results in decreased β-MHC expression in the adult heart, providing evidence that miR-208a is involved in the regulation of Myh7 and Myh7b expression and their intronic miRNAs, miR-208b and miR-499 (Callis et al., 2009; van Rooij et al., 2009).
Implication of miR-208a in Heart Disease
The expression of miR-208a is not just developmentally regulated in the heart, but is also deregulated in some pathological processes (Table 1). Cardiac contractility depends on the expression of the two MHC isoforms, α- and β-MHC, and changes in their proportions may lead to hypertrophy, fibrosis, and serious effects on the contractile function of the heart. Indeed, increase dexpression of β-MHC in the myocardium, a common feature of cardiac hypertrophy and heart failure, may decrease power output and can contribute to depressed systolic function in end-stage heart failure (Stelzer et al., 2007). Within adult heart, a switch in the isoforms arrangement from α- to β-MHC can be induced by mechanical stress and hypothyroidism (Gupta, 2007).
Loss of miR-208a in mice leads to a pronounced expression of numerous fast skeletal muscle contractile protein genes, which are normally not expressed in the heart (van Rooij and Olson, 2007). This activation of the skeletal muscle gene program is induced by direct inhibition of Med13 (also known as THRAP1), a cofactor of the thyroid hormone nuclear receptor (van Rooij and Olson, 2007; Callis et al., 2009). Med13 is involved in the thyroid hormone signaling to repress β-MHC expression in the adult heart (Morkin, 2000). However, overexpression of miR-208a in the heart results in pronounced repression of Med13 leading to an increasing of β-MHC expression. This high level of miR-208a expression is associated to arrhythmia, fibrosis, and hypertrophic growth in mice (Callis et al., 2009) and poor clinical outcomes in human dilated cardiomyopathy. The high level of miR-208a expression has been proposed as a strong predictor of cardiac death and heart failure (Satoh et al., 2010). On the other hand, an experimental model of miR-208a null mice failed to upregulate β-MHC expression and did not develop cardiomyocytes hypertrophy or fibrosis (van Rooij and Olson, 2007).
Within a hypothyroidism state, β-MHC expression is upregulated; however, mice lacking miR-208a are unable to upregulate β-MHC expression in response to hypothyroidism induction (van Rooij and Olson, 2007). Interestingly, in this model, the expression of β-MHC before birth was normal. This result suggests that miR-208a acts specifically at the postnatal level occurring at the same time of the acquisition of thyroid hormone responsiveness of the gene encoding β-MHC (van Rooij and Olson, 2007).
In addition, the analysis of mice lacking miR-208a revealed that miR-208a is required for proper cardiac conduction and expression of the cardiac transcription factors. They are known as important for the development of the conduction system, such as GATA4 and gap junction protein connexin 40. This study also suggests that these mice suffer atrial fibrillation (Callis et al., 2009).
In a remarkable postmortem study, Boštjančič et al. (2010) show that miR-208a is upregulated in human hearts after myocardial infarction (MI) remains elevated for up to 4 weeks, whereas miR-1 and miR-133a are downregulated. These results showed the involvement of miR-208a as well as miR-1 and miR133a in human MI providing new insights into miRNA expression profiles in the acute, subacute, and chronic phases following MI (Boštjančič et al., 2010).
miR-208a as a Potential Biomarker and Therapeutic Target
Many miRNAs are known as tissue specific. This way, most of the clinical studies have been based on the measurement of expression levels of miRNAs in origin tissue samples. However, numerous studies have demonstrated that some miRNAs, including miR-208a, are not only found intracellularly, but also detectable outside cells, including various body fluids, such as blood, saliva, and urine (Zhu and Fan, 2011). These miRNAs were named circulating miRNA or c-miRNAs. Interestingly, ∼90% of extracellular miRNAs are packed with proteins (i.e., Ago2, HDL, and other RNA-binding proteins) and ∼10% are wrapped in small membranous particles (i.e., exosomes, microvesicles, and apoptotic bodies) (Zhu and Fan, 2011). Recent evidences reveal that some proportion of c-miRNAs is secreted from normal healthy or damaged cells as microvesicles (Hunter et al., 2008). However, whether miR-208a is released by damaged cardiomyocytes into the bloodstream in the form of microvesicles, its precise mechanism of release remains unknown. The fact that miR-208a is enriched in the heart and maybe detected in peripheral blood makes it potentially useful for fast and easy tests, assisting the diagnosis or guiding therapy.
The first study in mice has shown that the plasma concentration of miR-208a is increased in myocardial injury presenting a good correlation with the plasma concentration of cardiac troponin, a gold standard marker of myocardial injury (Jaffe et al., 2000; Ji et al., 2009). In the following year, the levels of miR-1, miR-133 (Wang et al., 2010; Gidlöf et al., 2011), miR-208a (Wang et al., 2010), miR-208b (Corsten et al., 2010; Gidlöf et al., 2011; Devaux et al., 2012), and miR-499 (Corsten et al., 2010; Wang et al., 2010; Gidlöf et al., 2011; Devaux et al., 2012) were showed highly elevated in plasma and significantly reduced in the infarcted myocardium (Kuwabara et al., 2011). However, the plasma level of miR-208a could detect individuals with acute MI with 90.9% sensitivity at 100% specificity, although the levels of miR-499, miR-1, and miR-133a in plasma were less sensitive (36.4%, 33.3%, and 15.2%, respectively) for acute MI diagnosis (Wang et al., 2010).
The level of cardiac troponin in blood begins to rise after 4–8 h of myocardial injury (Wu and Feng, 1998), while an apparent elevation of miRNAs is observed within 1 h of coronary ligation (Wang et al., 2010). On the other hand, cardiac troponins are excreted from the kidney and occasionally its levels are increased in end-stage renal disease, even in the absence of heart disease (Abbas et al., 2005). Thus, plasma miR-208a may be superior to cardiac troponin for detecting myocardial injury in individuals with renal dysfunction. Furthermore, in the Wang et al. (2010) study, miR-208a could be detected in all the patients, while parallel cardiac troponin was detected in 85% of patients suggesting the plasma miR-208a might have a higher sensitivity as a potential biomarker for MI diagnosis (Wang et al., 2010).
In a study conducted by Widera et al. (2011), the plasma levels of miR-1, miR-133a/b, miR-208a/b, and miR-499 were measured in a large acute coronary syndrome (ACS) cohort. As result, miR-1, miR-133a/b, and miR-208b were independently associated with high-sensitivity troponin T (hsTnT) levels (all p<0.001). Patients with MI presented higher levels of miR-1, miR-133a, and miR-208b compared with patients with unstable angina. However, all six investigated miRNAs showed a large overlap between patients with unstable angina or MI. miR-133a and miR-208b levels were significantly associated with the risk of death in univariate and age- and gender-adjusted analyses. Both miRNAs lost their independent association with outcome upon further adjustment for hsTnT. However, contrasting results are observed comparing the results of three studies. In the Widera et al. (2011) study, the plasma levels of miR-208a were not elevated in ACS patients and Fichtlscherer et al. (2010) have shown that plasma levels of miR-208a tend to be high in patients with coronary artery disease (CAD). However, De Rosa et al. (2011) have shown that the plasma levels of miR-208a, as well as miR-499 and miR-133a, were significantly elevated in ACS patients compared with CAD patients.
Currently, some miRNA-based therapeutic strategies were developed based on the normalization of deregulated miRNAs levels in pathological processes (Oliveira-Carvalho et al., 2012). The antagomirs or antimiRs (antagonistic nucleotide sequences synthesized to bind in a specific mature miRNA) were developed to act as miRNA inhibitors leading to a decrease in the effect caused by the excessive expression of certain miRNAs. In this field, Montgomery et al. (2011) conducted an elegant study evaluating a therapeutic approach based in the manipulation of miR-208a levels. The authors delivered antimiR-208a by subcutaneous injection to inhibit miR-208a regulation in hypertension-induced heart failure mice. As a result, the miR-208a inhibition prevented pathological myosin switch and cardiac remodeling, while improving cardiac function, overall health, and survival. These results indicate the potential use of miR-208a as a therapeutic target for the modulation of cardiac function and remodeling during heart disease progression.
Recently, another approach around the obesity, type 2 diabetes, miR-208a, and the heart was conducted in a recent study by Grueter et al. (2012). The authors showed that the inhibition of miR-208a in mice leads to an increase in Med13 levels conferring resistance to high-fat diet-induced obesity and improves systemic insulin sensitivity and glucose tolerance. Conversely, deletion of Med13 specifically in cardiomyocytes enhances obesity in response to high-fat diet and exacerbates metabolic syndrome. The metabolic actions of Med13 result from increased energy expenditure and regulation of numerous genes involved in energy balance in the heart. These findings reveal a role of the miR-208a in systemic metabolic control and point to miR-208a and Med13 as potential therapeutic targets for metabolic disorders (Grueter et al., 2012).
Considerations
The emerging knowledge about the role of miR-208a in the heart is changing our understanding about how the molecular network governs the heart development and disease. Evidences suggest that miR-208a is sufficient to induce cardiac remodeling, arrhythmias, and regulate the expression of hypertrophy pathway components, including upregulation of β-MHC and miR-208b in response to stress and hypothyroidism. The abnormal expression of miR-208a may give rise to a heart failure phenotype and an increase in myocardial collagen accumulation, leading to irreversible myocardial injury in humans. On the other hand, the emerging knowledge about the miR-208a opens a wide range of possibilities around the potential use of this miRNA as a biomarker of heart injury as well as new exciting therapeutic tools to improve the function of the diseased heart and clinical outcomes.
Footnotes
Disclosure Statement
No competing financial interests exist.