
Abstract
Neuropathy target esterase (NTE), which has been proposed as the primary target of organophosphorus compounds that cause delayed neuropathy with degeneration of nerve axons, is expressed primarily in neural cells but is also detected in non-neural cells. However, little is known about the regulation of NTE gene in cells. We found that a cyclic-AMP (cAMP)-response element (CRE) exists in the 5′ flanking sequence of NTE gene in HeLa cells, which implies that NTE may be regulated by the transcription factor cAMP-response element-binding protein (CREB). In the study, knockdown of CREB decreased the protein and mRNA levels of NTE and inhibited the upregulation by cAMP/PKA signaling. Moreover, we observed that knockdown of CREB significantly decreased luciferase activity of the NTE gene promoter, while it had no effect on that of the CREB binding sites of mutated NTE gene promoter and truncated NTE gene promoter lacking the CREB binding site. cAMP/PKA signals could increase NTE reporter gene activity, while knockdown of CREB inhibited the increase. We found that the transcription factor CREB can bind to the promoter sequence of NTE by chromatin immunoprecipitation. In conclusion, we provided evidence that CREB is required for cAMP/PKA signals upregulating NTE expression in HeLa cells.
Introduction
Previous studies showed that NTE displayed potent lysophospholipase activity in the mouse brain. This activity has been characterized as a novel phospholipase B responsible for the conversion of phosphatidylcholine (PC) to glycerophosphocholine (GPC) and regulating PC homeostasis in Drosophila and the mouse (Zaccheo et al., 2004; Muhlig-Versen et al., 2005; Read et al., 2009). In mice, complete inactivation of the NTE gene led to embryonic lethality resulting from placental failure and impaired vasculogenesis (Winrow et al. 2003; Moser et al. 2004). In adult Drosophila, loss of Swiss cheese/NTE activity causes neuronal and glial death (Akassoglou et al. 2004; Muhlig-Versen et al. 2005). Taken together, these studies suggest that NTE is essential for embryonic and nervous development.
Elucidation of the role played by NTE in OPIDN and neural development is of obvious neurobiological interest. However, NTE is present not only in neurons but also in non-neural tissues including those found in the intestines, placenta, lymphocytes, kidneys, liver, and testicles (Williams, 1983; Li et al., 2003; Winrow et al., 2003; Chen et al., 2012). Therefore, NTE may play a universal role in mammalian neurons and non-neural cells. Further, there is a putative homologue in Saccharomyces cerevisiae (Lush et al., 1998), indicating that NTE might be involved in a common physiological process from yeast to mammal (Li et al., 2003).
Although the characteristics of NTE have been elucidated in some animals, little is known about the regulation of NTE. In a previous study, we demonstrated that NTE can interact with the protein Gβ2 through its catalytic domain. We also showed that pertussis toxin, a G protein signal pathway influencing factor, and the depletion of Gβ2 by RNA interference, can downregulate NTE activity in vivo (Chen et al., 2007). NTE expression can also be regulated by cyclic-AMP (cAMP)/PKA signal in HeLa cells (Chen et al., 2010). The TFSEARCH software was used to identify putative consensus binding sites for cAMP-response element-binding protein (CREB) on the NTE gene promoter. NTE proximal promoter contains a cAMP-responsive element (CRE) between −83 and −76 base pairs (bp). However, there is no existing evidence whether the NTE expression can be regulated by CREB in mammalian cells.
The aim of the present study is to investigate whether cAMP/PKA/CREB signals regulate NTE expression in mammalian cells. This finding will facilitate the future investigation of the molecular mechanism governing the human NTE gene expression.
Materials and Methods
Cells and chemicals
HeLa cells were purchased from the Cell Center of the Chinese Academy of Medical Sciences (Beijing, China). Cell culture reagents were obtained from Gibco BRL. Rabbit anti-NTE polyclonal antibody (H-200), rabbit anti- actin monoclonal antibody (C-2), Rabbit anti-CREB1 polyclonal antibody (sc-58), and goat anti-rabbit IgG-HRP were purchased from Santa Cruz Biotechnology. Forskolin and N-[2-(p-bromocinnamylamino)ethyl]-5-isoquinolinesulfonamide (H89) were obtained from Sigma. The pGenesil-1 vector was obtained from Genesil Biotechnology Co., Ltd. The enhanced chemiluminescence (ECL) reagent was obtained from Pierce Biotechnology. The TRIzol reagent, transfection reagent Lipofectamine 2000, and chromatin immunoprecipitation (ChIP) system were purchased from Invitrogen Life Technologies. First-strand cDNA Synthesis Kit (ReverTra Ace-α-™) for reverse transcription–polymerase chain reaction (RT-PCR) was obtained from Toyobo Co. Ltd. EX Taq Hot Start DNA polymerase was purchased from Takara. Site-Directed Mutagenesis Kit was obtained from TransGen. Dual-Luciferase® Reporter Assay System was purchased from Promega.
Cell culture and transfection
Cells were grown and maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 IU/mL penicillin, and 100 μg/mL streptomycin. Incubations were carried out at 37°C in a humidified atmosphere of 5% CO2/95% air. The cells were maintained in the logarithmic phase of growth and subcultured at 3–4 days intervals. Transient transfection of control shRNA and RNAi-CREB in HeLa cells was performed using Lipofectamine 2000 according to the manufacturer's protocol.
Construction of CREB shRNA vector
To construct shRNA vector for human CREB (NM_004379), the pGenesil-1 vector (Genesil Biotechnology Co., Ltd.) was digested by BamHI and HindIII. Next, CREB-F: 5′-
Cell treatment
HeLa cells were transiently transfected with control shRNA and RNAi-CREB for 24 h; the cells were treated with forskolin (10 μM), H89 (10 μM), an inhibitor of PKA activity, and H89 plus forskolin for 24 h to observe the effect of cAMP/PKA/CREB on the mRNA and protein levels of NTE.
Western blotting analysis
The cells were trypsinized, washed twice with ice-cold PBS, and homogenized in 1 mL TE buffer (50 mM Tris-HCl and 0.2 mM EDTA, pH 8.0) on ice. The homogenate was then centrifuged at 100 g at 4°C for 2 min after which the supernatant was then centrifuged at 100,000 g at 4°C for 45 min. The deposit was dissolved in TN buffer (150 mM NaCl, 1% Triton X-100, and 50 mM Tris-HCl, pH 8.0) and the protein concentration determined according to the method of Lowry et al. (1951) using bovine serum albumin as a standard. The protein samples were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis with a 5% stacking gel and 8% separating gel and transferred to polyvinylidene fluoride membrane (Millipore Corporate). Following transfer, membranes were blocked with 1× Tris-buffered saline buffer containing 0.05% Tween-20 and 5% nonfat milk for at least 1 h at room temperature, then incubated with rabbit polyclonal anti-NTE antibody (diluted 1:1000), and finally incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (diluted 1: 5000). Immunoreactive bands were detected using a ChemiDoc XRS system (Bio-Rad) and standard ECL reagents. The blot was stripped and re-probed with anti-actin antibody to detect the loaded protein.
RT-PCR analysis
Total RNA was isolated from HeLa cells with TRIzol reagent according to the manufacturer's instructions and spectrophotometrically quantified after DNase I treatment. The total RNA (1 μg) was reverse transcribed using the first-strand cDNA synthesis kit in a final volume of 20 μL, according to the manufacturer's instructions. PCR analyses were then performed on the aliquots of the cDNA preparations to detect NTE and GAPDH gene expression using a thermal cycler (Eppendorf AG 22331 MasterCycler). The reactions were carried out in a volume of 25 μL containing 2.5 U of Taq DNA polymerase, 0.2 mM deoxyribonucleoside triphosphates, 1×reaction buffer, and 100 pmol of specific 5′ and 3′ primers. Specific primers were designed for the NTE sequence with Primer Express software (Applied Biosystems). The forward primer was 5′-CCAAGAGTTCCGGCTGTCA-3′ and the reverse primer was 5′-CACAATGAGGATGCAGTCGG-3′. The forward and reverse primers for CREB sequence were 5′-CACCAGGAGTGCCAAGGATT-3′ and 5′-CTGCTGCATTGGTCATGGTT-3′. The forward and reverse primers for GAPDH sequence were 5′-ATGCTGGCGCTGAGTACGTC-3′ and 5′-GGTCATGAGTCCTT CCACGATA-3′, respectively. For the indicated gene amplification, the denaturation cycle (94°C, 5 min) was followed by 35 cycles, each cycle consisting of DNA denaturation at 94°C for 30 s, primer annealing at 52°C for 30 s, and elongation at 72°C for 1 min, and an elongation cycle at 72°C for 10 min at the end of amplication. PCR products were detected with a ChemiDoc XRS (Bio-Rad) using 1% agarose gel electrophoresis and ethidium bromide staining.
Cloning of the human NTE promoter and site-directed mutagenesis
A 1756 bp long DNA sequence comprising the 5′ flanking sequence upstream of the transcription start site of the human NTE gene was amplified by PCR using the following oligonucleotides: 5′-CG
The constructs containing mutated CREB binding sites were obtained using the Site-Directed Mutagenesis Kit (TransGen) following the manufacturer's instructions. The nucleotide sequence TGACGACT in the CRE binding site, at −83 to −76 bases in the 5′ flanking region of NTE gene, was mutated to TGAATACT. The forward primer was 5′-AGGTGGGCCCAGATTGAATACTTGCACGGGTT-3′ and the reverse primer was 5′-ATTCAATCTGGGCCCACCTCTCCCCTGCC-3′. The construct was designated pGL3-Mut-NTE promoter. The mutagenesis was performed with pGL3-NTE promoter construct as a template. The successful mutants were confirmed by sequence.
The constructs without CREB binding site were also constructed to pGL3-basic (was designated pGL3-NTE-51 promoter), the forward and reverse primers of the truncated NTE gene promoter were 5′-ATTTCCCAGGACTCCGGGCTACCAGAT CGGCCGTCCAGCTGGAATCAACCG-3′ and 5′-AGCTCGGTTGATTCCAGCTGG ACGGCCGATCTGGTAGCCCGGAGTCCTGGGAAATAGCT-3′.
Reporter gene analysis
To observe whether CREB was required for the NTE promoter gene activity, HeLa cells were transiently transfected with control siRNA or RNAi-CREB, then the different firefly luciferase constructs, such as pGL3-NTE-51 promoter, pGL3-NTE promoter or pGL3-Mut-NTE promoter, and Renilla luciferase phRL-TK vector (Promega) were transfected to determine the transcriptional activity of the NTE reporter gene.
To determine the effect of cAMP/PKA signal on the NTE promoter gene activity, HeLa cells were transiently transfected with pGL3-NTE promoter. Twenty-four hours after transfection, the cells were treated with forskolin (10 μM), H89 (10 μM), or H89 plus forskolin for 24 h. The transcriptional activity of the NTE reporter gene was determined using a dual luciferase assay kit (Promega). Firefly luciferase activity was normalized to Renilla luciferase activity (phRL-TK) and expressed as fold change over the mean of the untreated control group.
Chromatin immunoprecipitation
The ChIP experiment was performed following a previously described protocol (Dammer et al., 2007). Briefly, HeLa cells were cross-linked with 1% formaldehyde for 10 min, followed by termination of the cross-linking with 0.125 M glycine. Cell lysates were prepared and chromosomal DNA was sonicated to reach average sizes between 200–1000 bp. The lysate was immunoprecipitated with either rabbit IgG or an anti-CREB antibody. Upon reversal of the cross-linking, the purified DNA fragments were PCR-amplified for the promoter regions of NTE, GAPDH, and NR4A2 gene. The forward and reverse primers for NTE gene sequence were 5′-TTTGCTCCAT CCCTTAGTCC-3′ and 5′-AAGGAGTCGAAGGAACCCGT-3′; the primers for GAPDH gene were 5′-GAGAAAGTAGGGCCCGGCTA-3′ and 5′-GGTCTTGAGG CCTGAGCTACG-3′; the primers for NR42A2 gene were 5′-CCACCCAAGCTGGC TACCAA-3′ and 5′-GTTTATGTGGCTTGCGCTGC-3′. Output (immunoprecipitated promoter region) was normalized to input DNA.
Statistical analysis
Values were expressed as the mean±SEM. For comparison of the two groups, the statistical difference was determined by Student's t-test. Other data were evaluated by one-way analysis of variance with Newman–Keuls Multiple range test. For each test, p-values less than 0.05 (p<0.05) were considered statistically significant.
Results
CREB is required for NTE expression
To investigate whether CREB was required for NTE expression, the control shRNA and RNAi-CREB constructs were transiently transfected to observe NTE expression in HeLa cells. As shown in Figure 1, the protein and mRNA levels of CREB were significantly decreased after transfection with RNAi-CREB, indicating that the constructed RNAi-CREB vector works well. The protein and mRNA levels of NTE were significantly decreased in the HeLa cells transfected with RNAi-CREB compared with the control cells, indicating that CREB is required for NTE expression.
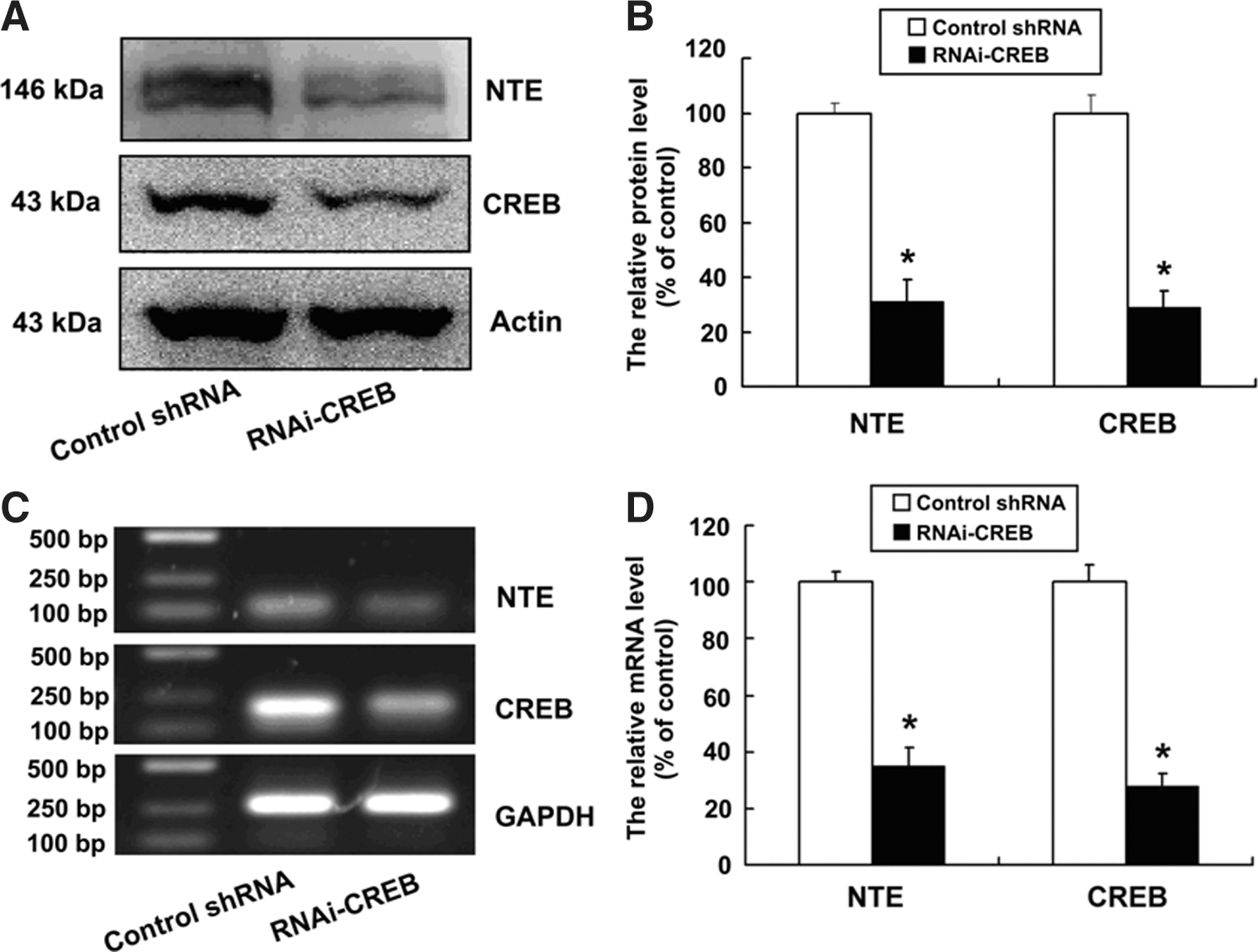
cAMP-response element-binding protein (CREB) knockdown downregulates expression of neuropathy target esterase (NTE) protein and mRNA. Cells were cultured at a density of 1×106 cells per 100-mm culture dish and were transiently transfected with control shRNA and RNAi-CREB for 48 h, respectively.
CREB binds to NTE promoter in vivo
Next, we wished to determine whether the CREB protein, a well characterized transcription factor, could bind to the CRE site in the NTE gene promoter. ChIP assay using CREB-specific antibody revealed that CREB could indeed bind to the NTE gene promoter (Fig. 2).

Binding of CREB to the NTE promoter in vivo. Sheared chromatin prepared from HeLa cells was immunoprecipitated (IP) with IgG or anti-CREB antibody and the bound DNA was isolated according to the manufacturer's instructions (Biyotime). The immunoprecipitated DNA was used as a template for PCR using the designed primer sets. Immunoprecipitate of GAPDH and NR4A2 promoter (Conkright et al., 2003) were negative and positive control, respectively.
CREB regulates NTE reporter gene activity
To further define the mechanism by which CREB modulates NTE mRNA expression, we constructed reporter constructs, such as a pGL3-NTE-51 promoter without CREB binding site, a pGL3-NTE promoter having CREB binding site, and a pGL3-Mut-NTE promoter containing mutated CREB binding site and transiently transfected HeLa cells. The cells were co-transfected with these reporter constructs and control shRNA or RNAi-CREB for 48 h. As shown in Figure 3, the luciferase activity of pGL3-NTE-51 promoter was significantly lower than that of pGL3-Mut-NTE promoter and pGL3-NTE promoter, which might be due to pGL3-NTE-51 promoter having shorter bases and less transcription factors than pGL3-Mut-NTE promoter or pGL3- NTE promoter. However, knockdown of CREB significantly decreased luciferase activity of pGL3-NTE promoter, while it had no effect on that of pGL3-Mut-NTE promoter and pGL3-NTE-51 promoter, indicating that CREB could upregulate transcriptional activity of NTE gene.
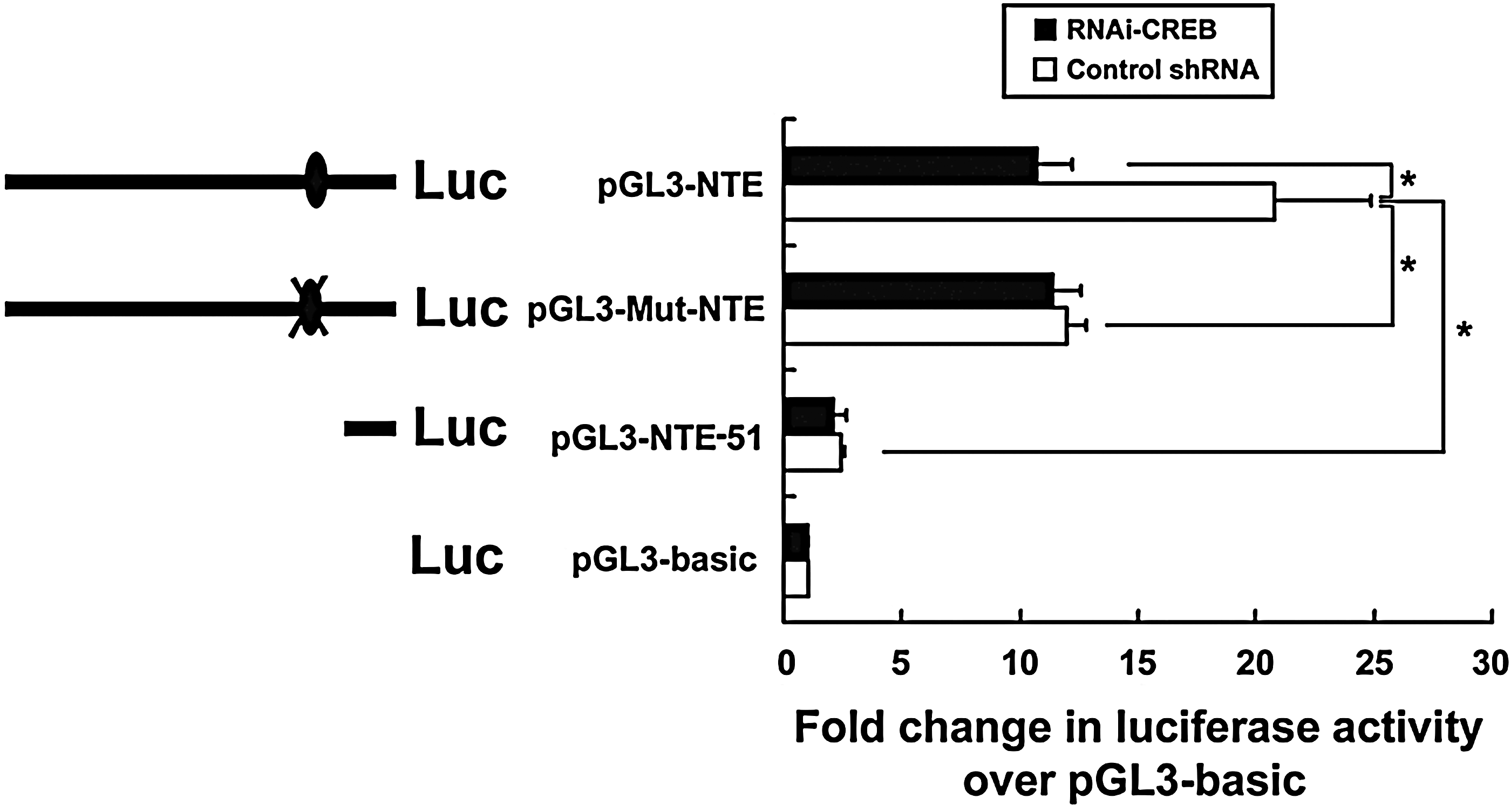
CREB is involved in the regulation of NTE promoter luciferase activity. HeLa cells were transfected with reporter gene plasmids such as pGL3-basic, pGL3-NTE-51 promoter, pGL3-NTE promoter, and pGL3-Mut-NTE promoter in the presence or absence of RNAi-CREB. 48 h after transfection, luciferase activity in the cell lysates was determined by luminometry. The experiments were done in triplicate and repeated three times. Data were analyzed by Student's t-test. The results are presented as Mean±SEM, n=3, *p<0.05.
cAMP/PKA signals is involved in upregulation of NTE by CREB
To determine whether cAMP/PKA signals could increase NTE reporter gene activity, we transiently transfected the cells with pGL3-NTE promoter in the absence or presence of knockdown of CREB, then treated the cells with FSK, H89, and H89 plus FSK for 24 h. As shown in Figure 4, the luciferase activity of pGL3-NTE promoter was significantly decreased after treatment with H89 and increased with FSK treatment, while FSK could not abrogate the decrease. Upon knockdown of CREB, however, H89, FSK, and H89 plus FSK had no effect on the luciferase activity.

cAMP/PKA/CREB signal is involved in the regulation of NTE promoter luciferase activity. HeLa cells were transfected with reporter gene plasmid pGL3-NTE promoter. Twenty four hours after transfection, cells were treated with FSK, H89, and H89 plus FSK for 24 h and luciferase activity in the cell lysates was quantified by luminometry. The experiments were done in triplicate and repeated three times. Data were analyzed by one-way analysis of variance (ANOVA). The results are presented as Mean±SEM, n=3, *p<0.05.
To further assess whether cAMP/PKA signals are involved in the upregulation of NTE by CREB, HeLa cells were transfected with control shRNA and RNAi-CREB, respectively, and then were treated with H89, FSK, and H89 plus FSK for 24 h. As shown in Figure 5, the protein and mRNA levels of NTE were significantly decreased after treatment with H89 and increased with FSK treatment, while FSK could not abrogate the decrease. Upon inhibition of CREB, however, H89, FSK and H89 plus FSK had no effect on the NTE protein and mRNA level.
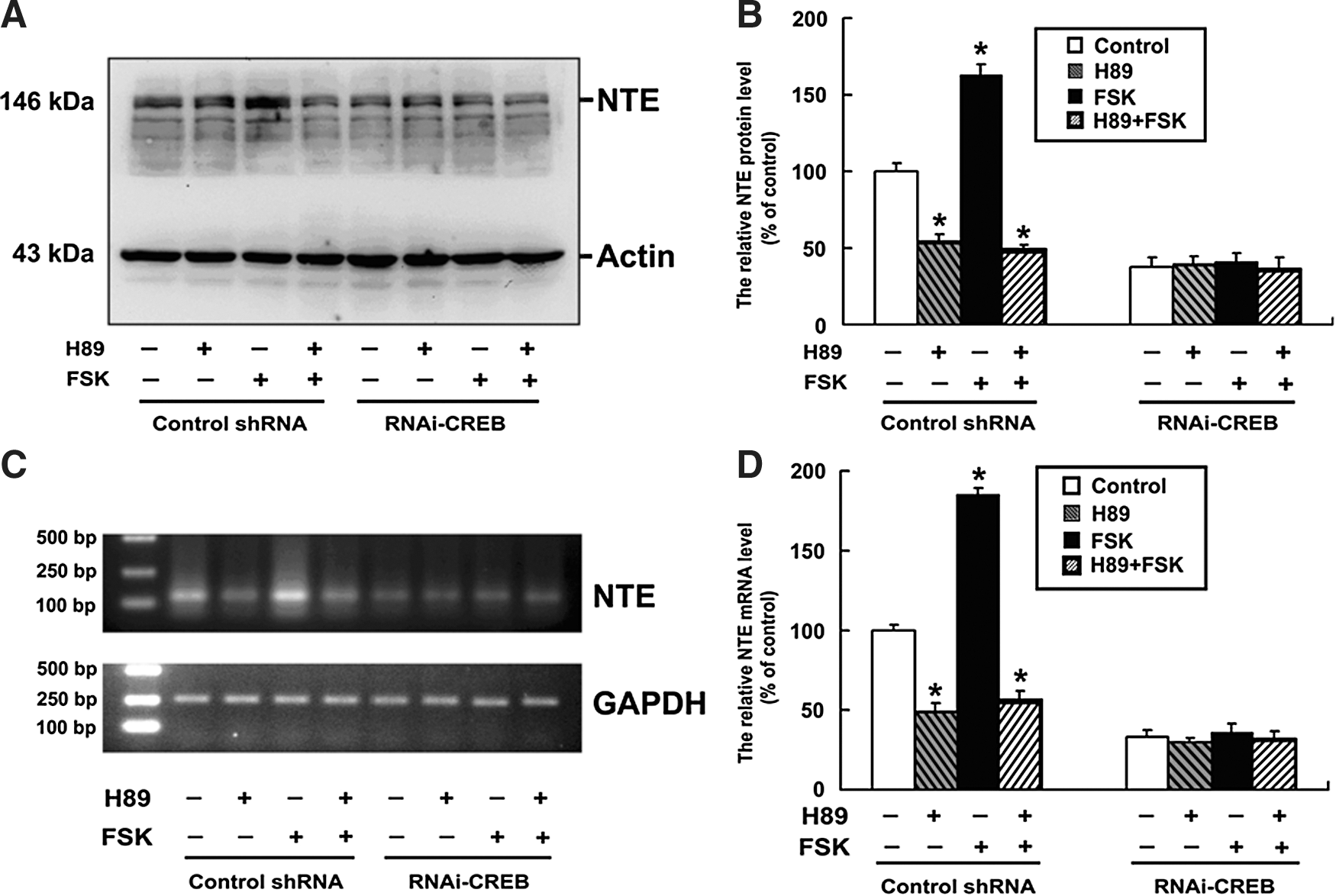
cAMP/PKA/CREB signal is required for the regulation of NTE expression. Cells were cultured at a density of 1×106 cells per 100-mm culture dish and were transiently transfected with control shRNA and RNAi-CREB for 24 h, respectively; the cells were then treated with H89 alone or H89 plus forskolin.
Discussion
In the present study, we provided evidence for the first time that cAMP/PKA/CREB signals could upregulate the protein expression of NTE in HeLa cells.
NTE was originally identified in adult vertebrate neural tissue as a protein targeted by OP, which cause the syndrome OPIDN, characterized by axonal degeneration (Johnson, 1974). However, NTE is present not only in neurons but also in many non-neural tissues such as intestine, placenta, lymphocytes, kidneys, liver, and testicles (Williams, 1983; Li et al., 2003; Winrow et al., 2003; Chen et al., 2012). In addition, NTE has shown to catalyze hydrolysis of PC to GPC (Zaccheo et al., 2004), indicating that NTE may play an important role in maintaining cellular phospholipid homeostasis.
CREB is a leucine zipper-containing transcription factor that regulates the expression of several genes by binding to CRE sequences in the promoter of the target gene (Mayr and Montminy, 2001; Sands and Palmer, 2008; Yamamizu et al., 2012; Wu et al., 2012).
CREB is activated by diverse stimuli such as growth factors, hormones, and cytokines and plays important roles in various cellular processes, including proliferation, survival, development, and adaptive responses (Rudolph et al., 1998; Mantamadiotis et al., 2012; Rexach et al., 2012). We found that there was a putative consensus binding site for CREB in the NTE promoter, indicating that NTE might be regulated by CREB. In this study, knockdown of CREB significantly decreased the protein and mRNA levels of NTE in HeLa cells about 70%. CREB is the common target of cAMP/PKA signaling (Impey et al., 2004; Zhang et al., 2005); PKA can phosphorylate CREB at Ser 133, a post-translational medication that is essential for its transcriptional activity (Gonzalez and Montminy, 1989; Mayr and Montminy, 2001). We also found that NTE expression was upregulated by cAMP/PKA signal (Chen et al., 2010). In the present study, forskolin and H89 significantly increased and decreased NTE expression without knockdown of CREB in the HeLa cells. Upon CREB knockdown, both forskolin and H89 had no effect on the expression of NTE, indicating that CREB was required for regulation of NTE by cAMP/PKA signal.
To investigate whether cAMP/PKA/CREB affects transcription activity of NTE gene, we constructed an NTE gene promoter, a CREB binding site mutated NTE gene promoter, and a truncated NTE gene promoter lacking the CREB binding site into pGL3-basic vector and transfected to HeLa cells. We found that knockdown of CREB significantly decreased luciferase activity of NTE gene promoter about 50%, while it had no effect on the CREB binding site of mutated NTE gene promoter and truncated NTE gene promoter lacking the CREB binding site. The difference in the decrease between the protein and mRNA levels, and luciferase activity of NTE after knockdown of CREB might be due to the difference of the assay methods.
Although the CREB binding site in the pGL3-Mut-NTE promoter was mutated, it might have other transcription factors that might regulate NTE transcription. Thus, the luciferase activity of pGL3-Mut-NTE promoter and pGL3- NTE promoter significantly increases more than that of pGL3-NTE-51 promoter. cAMP/PKA signals increased NTE reporter gene activity, while knockdown of CREB inhibited the increase. The results indicated that cAMP/PKA/CREB could upregulate transcriptional activity of NTE gene. We also identified that transcription factor CREB can bind to the promoter sequence of NTE by ChIP.
It was found that diisopropylphosphorofluoridate, an OPIDN-causing compound, caused a decrease in the levels of PKA and p-CREB in cerebrum nuclear fraction of hens at 8 h and on 10 days after administration (Damodaran et al., 2009). The finding indicated that PKA and p-CREB might play an important role in the initiation, progression, and development of OPIDN, while the inhibition and subsequent aging of NTE has been proposed to be an initiating event in OPIDN (Johnson, 1974, 1977; Hou et al., 2008). These studies indicated that inhibition of PKA and p-CREB causing OPIDN might be associated with NTE inhibition; however, whether cAMP/PKA/CREB might be involved in OPIDN via inhibition of NTE needs to be further studied.
In conclusion, we provided direct evidence for the first time that CREB can bind to the promoter region of NTE gene and upregulate NTE expression in HeLa cells. We also showed that cAMP/PKA signaling is involved in the upregulation of NTE expression by CREB.
Footnotes
Acknowledgments
This work was supported in part by the grants from the National Natural Science Foundation of China (No. 30870537, 31071919) and the National Basic Research Program of China (No. 2012CB114100). We thank Ms. H. Huang for polishing the English of the article.
Disclosure Statement
No competing financial interests exist.