
Abstract
We previously showed the anticancer property of crocin, a carotenoid isolated and purified from saffron against chemical-induced gastric and breast cancer in rats. In this study, the mechanism of crocin action was investigated in the gastric adenocarcinoma (AGS) cells in comparison with human normal fibroblast skin cells (HFSF-PI3). Crocin revealed a dose- and time-dependent cytotoxic effect against an AGS cell line, as determined by 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Crocin-induced apoptosis was evidenced by flow cytometry and measuring caspase activity. The increased sub-G1 population and activated caspases in the treated AGS cells confirmed its anticancer effect. Expression of both Bax and Bcl-2 was determined using a semiquantitative reverse transcriptase–polymerase chain reaction and Western blot in these cells before and after treatment with crocin. Apoptosis was significantly stimulated as indicated by increasing the Bax/Bcl-2 ratio after crocin treatment. All of the above-mentioned parameters remained normal in HFSF-PI3 treated with crocin. These data are providing insight into the molecular mechanisms underlying the crocin-induced apoptosis in the AGS cells, rendering it as the potential anticancer agent.
Introduction
Recently, our team has investigated the mechanism of crocin action in the term of its interaction with high-molecular-weight DNA, different oligonucleotides (Bathaie et al., 2007; Hoshyar et al., 2008; Hoshyar et al., 2012), histone H1, and H1–DNA complex (as a model of chromatin) (Ashrafi et al., 2005). Our results showed that crocin bound to DNA through the minor groove and induced the B-to-C-DNA transition, which was accompanied by DNA precipitation at higher concentrations of crocin (Bathaie et al., 2007). In addition, the great potential of this ligand to treat the rat models of both chemical-induced breast and gastric cancers was shown in our laboratory [manuscripts under preparation]. However, the exact mechanism of crocin action is not known, yet. Furthermore, according to our literature review, the effect of crocin on human gastric cancer cell lines has not been evaluated, yet, so that the aim of the present study was to investigate the inhibitory effect of crocin on the proliferation and induction of apoptosis in the human adenocarcinoma gastric cell line, AGS, in comparison with the normal human fibroblast cells HFSF-PI3. In addition, the molecular mechanism of crocin action was investigated. Therefore, the impact of crocin on Bax and Bcl-2 expression, at both mRNA and protein levels, and activity of caspases were studied.
Materials and Methods
Crocin preparation
Crocin was separated and purified from the Iranian saffron by using Alumina-90-active column chromatography and eluted with ethyl acetate: ethanol: water (10:3:2, v/v/v). The progress of the purification processes was followed by UV-Vis spectroscopy, TLC, determining the melting temperature and absorbency at 440 nm (Bolhasani et al., 2005).
Cell lines and culture conditions
Human gastric carcinoma (AGS) and human normal fibroblastic cell lines (HFSF-PI3) were purchased from the Iranian Biological Resource Center (Tehran, Iran) and Pasteur Institute (Tehran, Iran), respectively. Cancerous and normal cells were cultured in the Ham's F-12 (Bio west) and Dulbecco's Modified Eagle's media (DMEM; Gibco), respectively. Cells were supplemented with 10% heat-inactivated fetal bovine serum (Invitrogen), 100 units/mL penicillin, and 100 mg/mL streptomycin (Gibco). Both cell lines were grown at 37°C in a humidified atmosphere containing 5% CO2. Before treatment, the cells were grown to 90% confluency and starved by incubation in a basal medium for 24 h. Then, they were treated with different concentrations of crocin (0–3.5 mg/mL) at various time intervals (0–72 h). All treatments were performed in triplicate.
In vitro cytotoxicity assay
The cytotoxic effects of crocin on AGS and HFSF-PI3 cells were measured with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, a tetrazole; Sigma-Aldrich) assay. The cells were seeded onto the 96-well plates for 24 h. Various concentrations of crocin were added to the cells. After incubation at different time courses, the medium was removed, and 200 μL of 1× MTT solution was added to each well for 4 h. The supernatant was removed by aspiration, and 150 μL dimethyl sulfoxide was added to the wells to dissolve any precipitate. The absorbance was then read at 540 nm using an ELISA plate reader (Awareness). For analysis of the cytotoxic efficiency, the IC50 value (drug concentration that reduced the absorbance of treated cells by 50% compared to untreated cells) of crocin was calculated using the dose- and time-dependent curves by linear interpolation. In addition, the cell viability was calculated by dividing the absorbance of treated cells in each well to the mean absorbance of control cells. All experiments were performed in triplicate, independently, and the results were expressed as the mean±SD.
Cell cycle analysis and sub-G1 measurement
Briefly, 2×105 cells were plated on 6-cm dishes for 24 h. The cells were then treated with different concentrations of crocin at various time intervals, followed by collecting the cells by centrifugation. The cells were fixed overnight with cold 70% ethanol and then stained with a propidium iodide (PI) solution (45 mg/mL PI, 10 mg/mL RNase, and 0.1% Triton X-100). After 1-h incubation at room temperature in the dark, fluorescence-activated cells were sorted in a FACScan flow cytometer (equipped with a 488-nm argon laser) using CellQuest 3.0.1 software (Becton Dickinson). In the nonapoptotic populations (as 100% after excluded sub-G1 population), the percentage of cells in each phase of the cell cycle was determined at least in triplicate and expressed as mean±SD.
Annexin V assay
The Annexin-V-FLUOS Staining Kit (Roche) was used for the apoptosis assay. AGS cells were seeded onto 6-cm dishes and then were exposed to different doses of crocin at different time intervals. Treated and control cancer cells were harvested by trypsinization, washed twice with ice-cold phosphate-buffered saline (PBS), and resuspended in 100 μL incubation buffer (HEPES buffer). Cell suspensions were then incubated with 20 μL of Annexin V- FLUOS and 20 μL of PI for 15 min at room temperature in the dark. The cells were evaluated immediately by flow cytometry. The percentage of cells undergoing apoptosis was determined by three independent experiments and expressed as mean±SD.
Caspase activity assay
Activation of caspases appears to be directly responsible for many of the molecular and structural changes in apoptosis. The enzymatic activities of caspase-2, 3, 6, 7, 8, 9, and 10 were detected by the Homogeneous Caspase Assay fluorometric kit (Roche). The assay was measured the free rhodamine 110 (R110) resulting from the cleavage of a common caspase substrate, DEVD, conjugated with R110. The amount of free R110 was determined fluorometrically at an excitation wavelength of 499 nm and an emission wavelength of 521 nm, and was expressed as relative fluorescence units by Gene 5 software with a fluorescence plate reader (Synorgy H4 Hybrid Reader). Caspase activity was expressed as a percentage of the activity from untreated cells.
RNA extraction and semiquantitative reverse transcriptase–polymerase chain reaction analysis
HFSF-PI3 and AGS cells were treated with 2.5 mg/mL crocin at 6–36 h, and the total RNA of collected cells was isolated by using RNx-plus reagent (Cinnagen, Iran). Then, the first-strand cDNA was synthesized according to the manufacturer's instructions of the RevertAid™ H Minus first-strand cDNA synthesis kit (Fermentas). The primer sequences for reverse transcriptase–polymerase chain reaction (RT-PCR) was selected according to the previously published article (Karagozlua et al., 2010), as follows: Bax, forward: 5′-TGC-CAG-CAA-ACT-GGT-GCT-CA-3′, reverse: 5′-GCA-CTC-CCG-CCA-CAA-AGATG-3′; Bcl-2, forward: 5′-CGC-ATC-AGG-AAG-GCT-AGA-GT-3′ reverse: 5′-AGC-TTC-CAG-ACA-TTC-GGA-GA-3′. As the internal control, the GAPDH gene was used. Its primers were as follows, forward: 5′-CAAGGTCATCCATGACAACTTTG-3′ and reverse: 5′-GTCCACCACCCTGTTGCTGTAG-3′ [RevertAid H Minus first strand cDNA synthesis kit (Fermentas)]. Amplification was performed in a MJ Mini™ Personal Thermal Cycler-Bio-Rad (Bio Rad) with cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and extension at 72°C for 30 s. After 30 cycles, the amplified PCR products were separated by electrophoresis on 1% agarose gels for 35 min at 85 V and visualized by ethidium bromide. All bands were semiquantitatively analyzed by densitometric resolution through image-capturing system software. Intensities of the mRNA levels were normalized to those of the GAPDH products as ratios to produce arbitrary units of relative abundance.
Protein preparation and Western blot analysis
AGS and HFSF-PI3 cells were treated with 2.5 mg/mL crocin for various durations, harvested, washed with ice-cold PBS, and lysed in an ice-cold lysis buffer (50 mM Tris, pH 7.4, 100 mM NaCl, 1 mM EDTA, 5% glycerol, 0.05% SDS, 1 mM phenyl methyl sulfonyl fluoride (PMSF), 0.1 mM NaF, 0.66 mM Na4P2O7, 0.5% Triton X-100, and protease inhibitor cocktail purchased from Sigma). After 30 min of incubation on ice, lysates were centrifuged at 12,000 g for 10 min at 4°C, and supernatants were collected. Protein content was determined by the BCA assay (Pierce), and samples containing 50 μg of proteins were loaded and separated by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Protein bands were transferred to polyvinylidene difluoride (PVDF) membranes (Roche). After blocking in the PBS solution containing 5% nonfat milk and 0.05% Tween-20 at room temperature overnight, the membranes were probed with specified primary antibodies (1: 500) at 4°C for 2 h. After washing in PBS containing 0.05% Tween-20, membranes were incubated with horse reddish peroxidase-conjugated goat anti-mouse secondary antibody (1:1000). Finally, immunoblot signals were visualized using an enhanced chemiluminescence advance Western blotting detection kit (Amersham). All the antibodies were purchased from Santa Cruz Biotechnology, Inc. The level of β-actin for each sample was used as a loading control.
Statistical analysis
Results were expressed as mean±SD of the indicated number of independent experiments. Student's t-test was used to compare the mean of each group with that of the control group, and p-values<0.05 were considered as significant.
Results
Cytotoxicity of crocin
Figure 1 shows the significant inhibitory effect of crocin on the AGS cell growth in a dose- and time-dependent manner (p<0.001). Parallel treatment of the normal cells with this component of saffron indicted a much less inhibitory effect on the viability of HFSF-PI3 cells. Analyses of the cell survival by MTT assay showed that the IC50 values of crocin on the AGS cells were 3, 2.7, and 2.5 mg/mL after 24, 48, and 72 h, respectively.

Effect of crocin on the viability of human adenocarcinoma gastric cell, AGS, and HFSF-PI3 cells. Cells were treated with different concentrations of crocin for 24, 48, and 72 h. Results are reported as the mean±SEM. ***Indicates the significance of data at the level of p<0.001 at different times and doses compared to the findings in normal cells.
Effect of crocin on cell cycle distribution of AGS cells
To elucidate the mechanism of its inhibitory effect on cellular proliferation, the capacity of crocin to inhibit the cell cycle progression was determined by flow cytometry. Figure 2A shows the effect of crocin at various durations on the cell cycle-phase distribution of the AGS cells. A sub-G1 apoptotic fraction that is a reliable biochemical indicator of cell death was markedly increased when crocin (3 mg/mL) was added to the cell culture medium at different time intervals (Fig. 2B). Moreover, after 24 h, crocin treatments with doses of 2.2, 2.5, 3, and 3.5 mg/mL resulted in an increase in the percentage of cells in the sub-G1 phase from 1.65% to 3.64%, 21.28%, 43.91%, and 60.4%, respectively, with no change on other phases of the cell cycle (Fig. 2C).
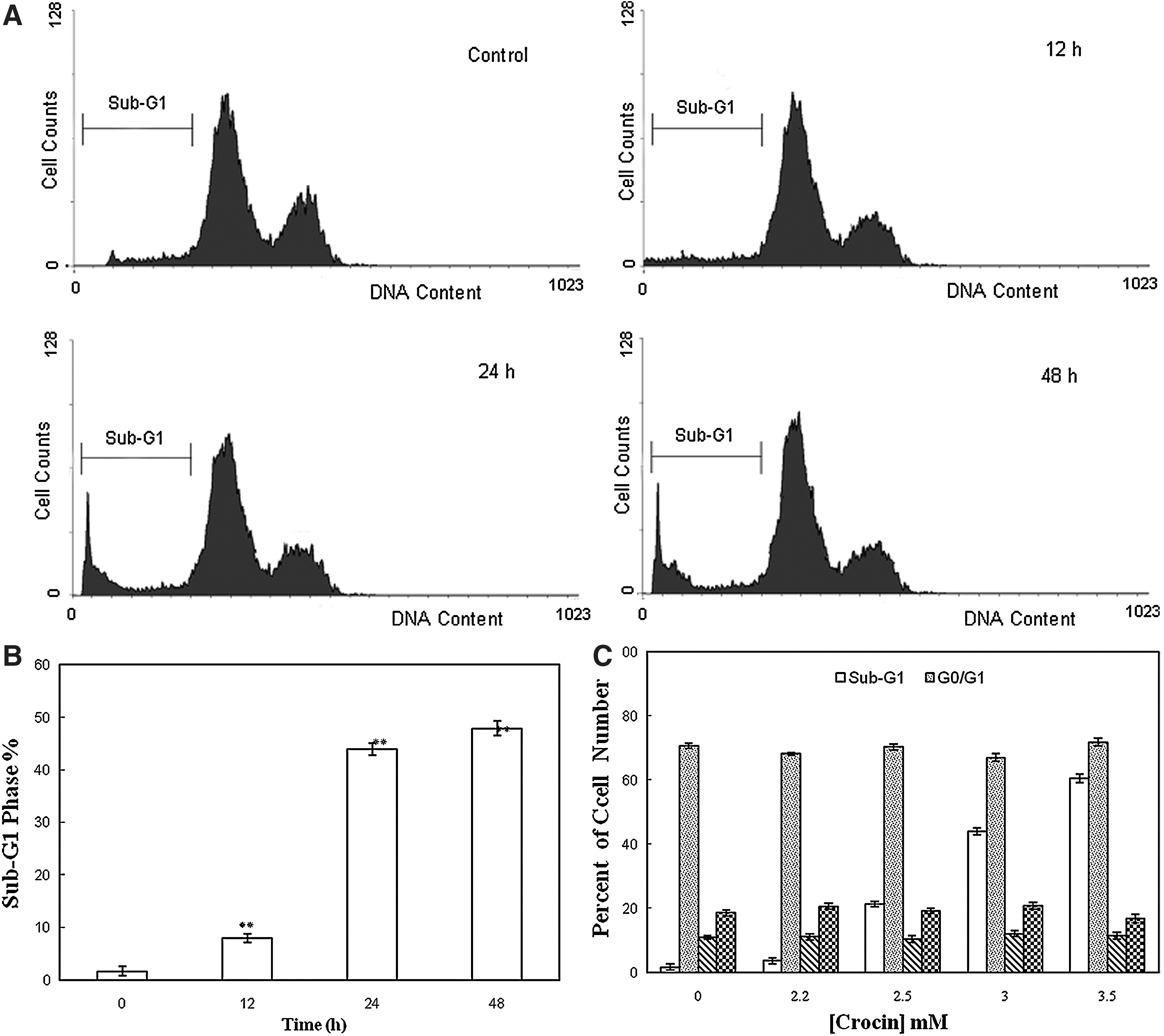
Flow cytometric analysis of cell cycle progression after crocin treatment. (
Apoptosis assay
Simultaneous staining of cells with Annexin V-FLOUS and PI distinguished between intact cells, early apoptosis, late apoptosis, or cell death. As shown in Figure 3A, the percentages of early apoptotic death (annexin V+/PI− and lower right quadrant) or late apoptotic and necrotic death (annexin V+/PI+ and upper right quadrant) increased in the time-dependent manner in the AGS cells treated with 3 mg/mL of crocin in comparison with the untreated AGS cells. Moreover, after the AGS cell exposition to different dosages of crocin (2–3.5 mg/mL) for 12, 24 and 48 h, the apoptotic cells were significantly increased (Fig. 3B).
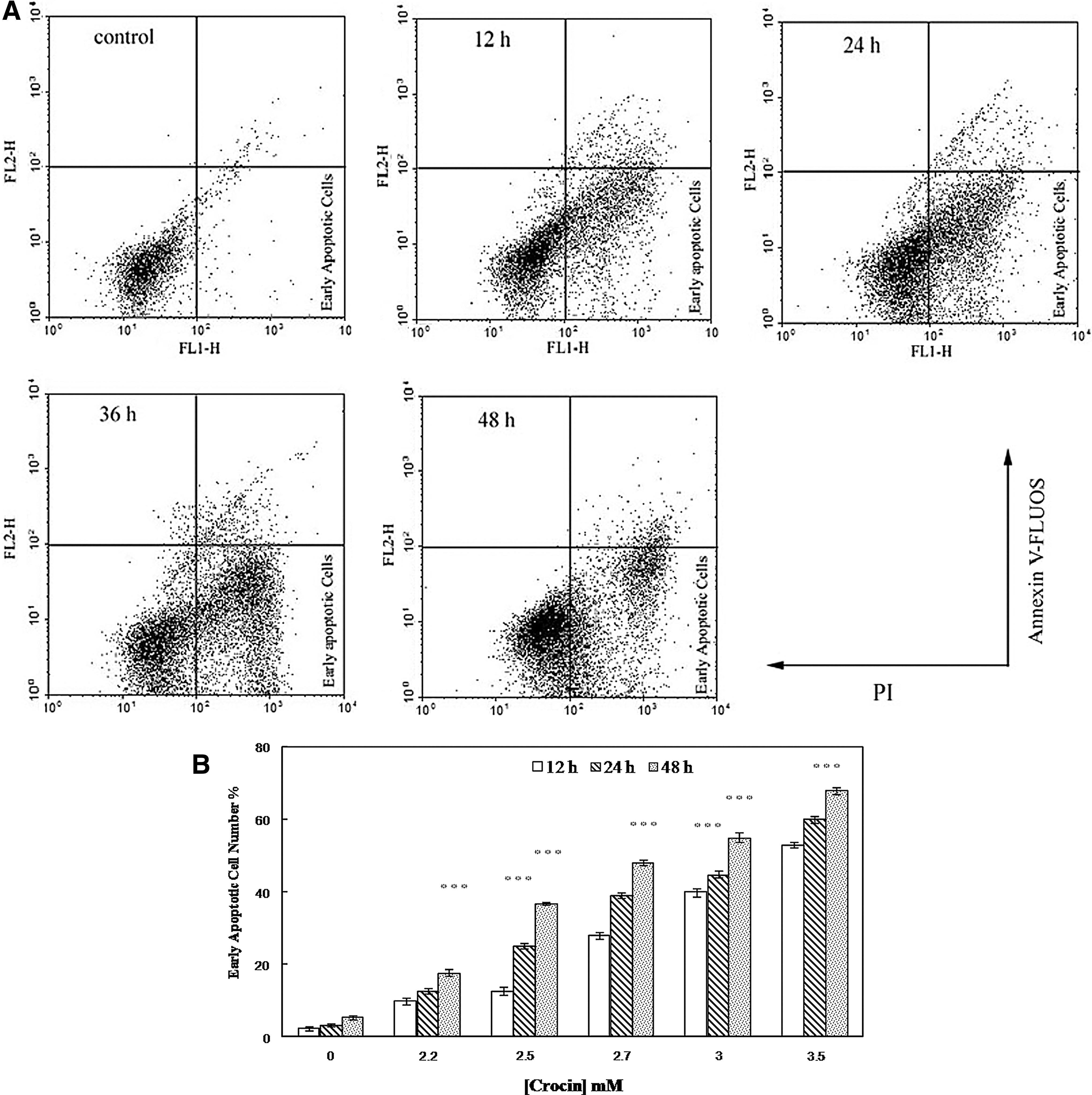
Fluorescence-activated sorting analysis of Annexin V-FLOUS and PI for quantification of crocin-induced apoptosis in AGS cells. (
Determination of the activation of caspases
The enzymatic activities of caspases were detected to confirm the crocin induced-apoptosis in the AGS cells. As shown in Figure 4A, after 6, 12, 24, and 48 h of treatment of AGS cells with crocin, the caspase activities were increased by 1.42±0.29-fold, 1.65±0.19- fold, 2.21±0.82- fold, and 4.13±0.8-fold, respectively.

Effect of crocin on AGS cell caspase activity. Caspase activity was determined after crocin (3 mg/mL) treatment of AGS cells from 6 to 48 h using the DEVD-R110 cleavage assay.
Detection of apoptotic-related gene expression in the AGS cells
Semiquantitative RT-PCR results demonstrated that crocin upregulated the expression of Bax and downregulated the expression of Bcl-2 in the AGS cells (p<0.05), in a time-dependent manner compared to the control cells (Fig. 5A, B). Notably, as shown in Figure 5C, elevation of the Bax/Bcl-2 ratio was observed only in the AGS-treated cells by 2.5 mg/mL of crocin and not in the HFSF-PI3-treated cells.

Effect of crocin (2.5 mg/mL) on the induction of apoptosis-signaling genes. (
Apoptosis-related protein expression after crocin treatment
Western blot analysis indicated that crocin (2.5 mg/mL) significantly increased Bax protein expression; however, it inhibited Bcl-2 protein expression, leading eventually to a time-dependent increase in the Bax/Bcl-2 ratio. In contrast, the Bax and Bcl-2 expression levels remained constant in normal cells treated with crocin at different times of incubation (Fig. 6).

Crocin effect on the expression of proteins involved in apoptosis. Western blot results of the cells were probed with antibodies specific to Bax and Bcl-2. β-actin was used as a loading control. Lanes from left to right are related to the increasing concentrations of crocin.
Discussion
Various pharmacological effects of saffron and its important constituents, crocin, have been considered in the two last decades (Bathaie and Mousavi, 2010); among them, the anticancer property of crocin has been more attended. Suppression of cell growth and induction of cell death are two major means for inhibition of tumor growth (Huang et al., 2000; Zhang et al., 2012). It was previously shown that crocin exerts cytotoxic activities in various human tumors, including acute promyelocytic leukemia cell line (HL-60), hepatocellular carcinoma (KIM-1), HeLa cells, lymphoid leukemia (MOLT-4B) cells, and K562 cells by inducing programmed cell death, and therefore arresting proliferation (Thatte et al., 2000). To evaluate the effect of this herbal ligand on cell proliferation, we investigated the effects of crocin on the growth of human gastric cancer AGS cells and human normal fibroblast skin HFSF-PI3 cells. Cells were exposed to increasing doses of crocin for 24, 48, and 72 h, and cell viability was determined by the MTT assay. As shown in Figure 1, cell viability in the AGS cell line was markedly decreased after exposure to crocin in a dose- and time-dependent manner. The nontoxic effect of crocin on the normal cell line, HFSF-PI3, was also indicated. It has been shown previously that crocin (3 mg/mL) is a nontoxic agent in normal cell lines (Aung et al., 2007; Noureini and Wink, 2012).
Apoptosis or programmed cell death is a gene-regulated phenomenon that was induced by many chemotherapeutic agents (Poncet and Kroemer, 2002; Fulda, 2010). Apoptogenic effects of saffron and its constituents have been shown in a few studies. For example, it has been reported that the saffron extract could induce apoptosis in HepG2 and HeLa cell lines (Escribano et al., 1996; Tavakkol-Afshari et al., 2008). Escribano et al. (1996) showed the crocin-induced morphological changes, including cell shrinkage and piknotic nuclei in the HeLa cells, suggesting that apoptosis could mediate the cytotoxic effect of crocin. In this study, to investigate the mechanisms responsible for this action of crocin, as the main secondary-active metabolite of saffron, on apoptosis induction in the cancerous cells in comparison with the normal cells, some additional experiments were done.
It has been reported that DNA fragmentation creates small fragments of DNA that can be eluted after incubation of cells in a hypotonic phosphate–citrate buffer. When stained with a quantitative DNA-binding dye such as PI, cells that have lost DNA will take-up less stain and will appear to the left of the G1 peak (Gorlach et al., 2011). Therefore, increased sub-G1 apoptotic fraction up to 43.91% in the AGS cells due to their treatment with 2.5 mg/mL crocin after 24 h was possibly the major cause of the inhibited proliferation (Fig. 2). Moreover, quantitative analysis of phosphatidylserine externalization by Annexin V- FLOUS and PI staining indicated that the percentage of Annexin V-positive and PI-negative cells was increased after their treatment with crocin, confirming the crocin-induced apoptosis in the AGS cells (Fig. 3). A similar mechanism of crocin action has been revealed previously against HT-29 and DHD/K12-PROb cells as the human and animal adenocarcinoma cells (Garcia-Olmo et al., 1999). Classical apoptosis can be initiated via two major pathways: the intrinsic or mitochondrion-mediated and the extrinsic or death receptor-mediated pathways, both of which result in the activation of different caspases (Fesik, 2005). Figure 4 indicated that after treatment with crocin, AGS caspase activities increased more than fourfold by time. However, there were no significant changes in the activity of these enzymes in the normal fibroblast cells. Therefore, caspase-dependent apoptosis also played an important role in crocin-induced AGS growth inhibition.
Genetic alternations may result in the disturbance of apoptosis-signaling pathways, which are critical components of carcinogenesis (Schulte-Hermann et al., 1994). Apoptosis is modulated by antiapoptotic and proapoptotic effectors, which involve a large number of proteins. The proapoptotic and antiapoptotic members of the Bcl-2 family act as a rheostat in regulating programmed cell death; therefore, they can be a target for anticancer therapy (Baell and Huang, 2002). Antiapoptotic Bcl-2 appears to function, at least in part, by interacting with and antagonizing proapoptotic Bax (Chan and Yu, 2004). During early apoptosis, Bax is inserted into the mitochondrial membrane and increases membrane permeability (Chan and Yu, 2004), leading to release of cytochrome C, activation of a variety of caspases, and cleavage of downstream death effector proteins, which ultimately result in apoptotic cell death (Chan and Yu, 2004; Baell and Huang, 2002), whereas antiapoptotic protein Bcl-2 prevents this process by preserving mitochondrial integrity (Kirkin et al., 2004). Actually, the balance between Bax and Bcl-2 is crucial for the induction of apoptosis. Therefore, deciphering of the mechanism of their expression modulation may be crucial for identifying therapeutic strategies for cancer. As shown in Figures 5 and 6, a time-dependent suppression of Bcl-2 expression and increment of Bax expression, in both mRNA and protein levels, were observed in the crocin-treated AGS cells. These results are compatible with those reported by Mousavi et al. (2009). They have showed that the saffron extract inhibited the growth of MCF-7 cells by increasing the Bax protein expression level and caspase activity (Mousavi et al., 2009).
Regarding to the relatively high doses of crocin used to induce the toxicity against cancer cells, two parameters should be considered: its solubility and safety for normal cells. Crocin can readily inter the cell and put its desirable effects. However, working with some herb components such as curcumin (Babaei et al., 2011) is restricted because of their limited solubility in the aqueous medium, and some additional process is needed for their stability and functionality. In addition, it has previously been shown that saffron carotenoids are not the precursors of vitamin A. Thus, the possibility of its long-term and high-dose application was recommended and introduced as one of the important advantages of crocin (Tarantilis et al., 1994).
In summary, our results demonstrated the anticancer effect of crocin that is a combination of its effects to inhibit the tumor cell growth and induce the tumor cell apoptosis by increasing the Bax/Bcl-2 ratio. The great interest is that normal cells were relatively insensitive to this herbal ingredient. With regard to its solubility and stability in the medium, it is suggested as an anticancer agent against stomach cancer in humans.
Footnotes
Acknowledgments
The authors would like to thank the Research Affairs of Tarbiat Modares University for financially supporting this project.
Disclosure Statement
The authors declare that there are no conflicts of interest.