
Abstract
This study addresses the potential application of adiponectin gene therapy and exercise in protection against skeletal muscle dysfunction in type 2 diabetes mellitus (T2DM) while focusing on the role of sarco and endoplasmic reticulum Ca+2 ATPase (SERCA) and Glut4. 50 rats were divided into five groups: control, T2DM, T2DM treated with either adiponectin gene or exercise or a combination of both. Serum glucose, insulin, HOMA index, triglycerides, and cholesterol were measured. Weight gain%, muscle contractile parameters {(peak twitch tension (Pt), peak tetanic tension (PTT), half relaxation time (HRT)}, and gene expression of SERCA, Glut4, and adiponectin were assessed in gastrocnemius muscle. Diabetic rats treated with either adiponectin gene or exercise showed significant reduction in all serum parameters and wt gain%. There was significant elevation in Pt and PTT with shortening in HRT. Furthermore, a significant increase in SERCA, Glut4, and adiponectin gene expression was noticed in both groups. Combination therapy caused marked gene expression of SERCA, GLUT4, and greater improvement in muscle contractility than either of the monotherapies. Skeletal muscle dysfunction in T2DM is mediated via impaired SERCA and Glut 4. Combination therapy offered best protection against muscle dysfunction and provides a novel promising strategy for a complete cure of muscle dysfunction in T2DM.
Introduction
Adiponectin is a 244–amino acid collagen-like protein that is mainly secreted by adipocytes and acts as a hormone with anti-inflammatory and insulin sensitizing properties (Kadowaki et al., 2006). Findings from animal studies and metabolic studies in humans suggest several mechanisms through which adiponectin may decrease the risk of T2DM. These include suppression of hepatic gluconeogenesis, stimulation of fatty acid oxidation in the liver and skeletal muscle, stimulation of glucose uptake in skeletal muscle, and stimulation of insulin secretion (Kadowaki et al., 2006). It was suggested that higher adiponectin levels are associated with a lower risk of T2DM (Li et al., 2009).
Exercise has many well-recognized beneficial effects in healthy as well as in diabetic subjects (Thompson, 2003; Boule et al., 2005; Pedersen, 2006). Regular exercise was recorded to improve insulin sensitivity in healthy individuals (Duncan et al., 2003; Boule et al., 2005). It also improves blood glucose control, and may prevent T2DM in high-risk individuals (Knowler et al., 2002; Sigal et al., 2004). It was shown that even with impaired insulin action, exercise-induced increase in skeletal muscle glucose uptake seemed intact (Martin et al., 1995).
Sarco and endoplasmic reticulum Ca+2 ATPase (SERCAs) are responsible for the maintenance of Ca2+ ion concentrations within the endoplasmic reticulum (MacLennan et al., 1985).Many studies linked insulin resistance in T2DM to impaired SERCA activity. Other studies reported several common dysfunctions of Ca2+ handling and alteration in SERCA activity in response to glucose in NIDDM model. This indicates a role of SERCA in the poor insulin secretion (Marie et al., 2001). Moreover, many novel links between SERCA and exercise were established. In mammalian skeletal muscle, SERCA activity has been shown to be acutely reduced after exhaustive exercise bouts (Byrd et al., 1989; Gollnick et al., 1991). Acute exercise training also has been found to improve Ca2+ sequestering performance of trained-striated muscle (Viru, 1994). In addition, previous studies showed that the impairment of sarcoplasmic reticulum (SR) function in diabetic cardiomyopathy is caused by reduced activity of SERCA2a which is due primarily to a decrease in SERCA2a expression (Wold et al., 2005).
In the present study, we have employed type 2 diabetic rats with skeletal muscle weakness to determine whether adiponectin gene therapy and regular swimming exercise regimen are able to counter the progression of skeletal muscle dysfunction-related T2DM in this commonly employed animal model. We also investigated the role of SERCA and adiponectin in the pathogenesis of skeletal muscle dysfunction in this model. In addition, the links between SERCA, adiponectin, and GLUT4 were studied.
Materials and Methods
The present experiment was reviewed and approved by the Committee of Ethics of Animal Experiments and conducted according to the Guidelines for Animal Experiments, German University in Cairo, Faculty of Pharmacy and Biotechnology.
Experimental animals and protocols
Eight-week-old albino male rats were obtained from the animal house of National Research Center, Cairo, Egypt. The animals were housed in wire mesh cages at room temperature. They were fed their respective diets with free access to water. Animals were randomly divided into five groups, and each group consisted of 8 rats: control group (CG), diabetic group (DG), DG treated by adiponectin gene (DAG), DG treated by swimming exercise (DEG), and DG treated by a combination of both (DAEG). T2DM was induced by the administration of a high fat diet (fat content 42% of energy), based on lard (HF-L), olive oil (HF-O), coconut fat (HF-C), or fish oil (derived from cod liver, HF-F) for a 12 week diet course (Buettner et al., 2006) with a single intraperitoneal injection of 45 mg/kg of streptozotocin (Sigma). Control rats were injected with the same volume of citrate buffer. Diabetes was confirmed on the second day post-STZ administration if serum glucose concentration was at least 250 mg/dL. At the end of the experimental, protocol rats were sedated with an intraperitoneal injection of thiopental Sodium before surgery. The left gastrocnemius was isolated from its distal insertion, keeping nerve and vasculature intact for the assessment of the physiological muscle contraction parameters, which include Peak Twitch Tension (Pt), Peak Tetanic Tension (PTT), and Half Relaxation Time (HRT).
Preparation and transfection of adiponectin gene
Adiponectin gene was taken from a muscle tissue to be amplified by RT-PCR and purified by using Qiagen Kit, where the purification solution is slightly acidic, containing guanine chloride, and this enhances DNA negative charge. DNA was washed to remove small pieces of DNA. This was achieved using Ethanol and Tris buffer. This solution solublizes smaller pieces of DNA. Lipofectamine™ 2000 (Invitrogen) was used as transfection kit, and it is a 3: 1 (W/W) liposome formulation of the polycationic lipid 2, 3-dioleyloxy-N-[2(sperminecarboxamido)ethyl]-N, N-dimethyl-1-propanaminium trifluoroacetate (DOSPA) and the neutral lipdioleoyl phosphatidylethanolamine (DOPE) in membrane-filtered water. Transfection activity was enhanced by using Plus™ Reagent (Cat. No 11514-015) to precomplex the DNA (Shih et al, 1997; Hawley-Nelson et al., 1993), where 0.2–0.4 μg DNA was diluted in 25 μL of Opti-MEM® I Reduced Serum medium and mixed gently. Lipofectamine™ was mixed gently before use, and then, 0.5–5 μL was diluted in 25 μL of Opti-MEM® I Reduced Serum medium and mixed gently. The diluted DNA was combined with diluted Lipofectamine™, mixed gently, and incubated for 15–45 min at room temperature. For each transfection, 0.15 mL of Opti-MEM® I Medium was added to the tube containing the complexes and mixed gently. Finally, 0.2 mL of the diluted complexes was added by an intramuscular injection.
Exercise protocol
Exercise training was carried out by long-term swim training, over a period of 9 weeks (Reynolds et al., 1997). Animal training was done in a group swimming method (Ueno et al., 1997) in a half-filled cylindrical plastic container, containing water at room temperature, in a session of 10 min, 5 days/week. The rats were acclimated to swimming by gradually increasing the swimming time and sessions as follows: 1st 2 weeks, 10 min, 1 session, 2nd 2 weeks, 20 min, 2 sessions, 3rd 2 weeks, 30 min, 3 sessions, last 3 weeks, 40 min, and 4 sessions. Sessions were separated by 1 h rest (Ploug et al., 1990; Peijie et al., 2004; de Lemos et al., 2007).
Biochemical analysis
Plasma glucose was assayed by an automated glucose oxidase procedure using a Beckman Glucose Analyzer II (Beckman Instruments) (Gault et al., 2002). Plasma insulin was determined using ELISA kit for the measurement of rat/mouse insulin (Linco Research). Determination of homeostasis model assessment of insulin resistance (HOMA-IR): HOMA-IR was calculated as the product of fasting insulin (microunits/mL) multiplied by fasting glucose (mM) divided by 22.5 (Matthews et al., 1985). Plasma triacylglycerol and cholesterol levels were measured using a Triglyceride Quantification Kit (BioVision Research) (Wahlefeld, 1974) and a Cholesterol Assay Kit (BioAssay System) (Roeschlau et al., 1974). All analyses were carried out according to the manufacturer's instructions.
Assessment of muscle physiological parameters
Gastrocnemius muscle was isolated from its distal insertion, keeping nerve and vasculature intact, and secured to an MLT0210/D force-displacement transducer by a small metal hook fastened to the calcaneal tendon. The leg was secured by screws tightened onto the medial and lateral condyles of the femur. Using a MLA0320 Stimulator, the muscle was stimulated through twin platinum electrodes that were applied directly to the surface of the muscle. Optimal voltage was determined by generating single twitch contractions at increasing voltages until no increase in single-twitch force production was observed. Optimal muscle length was determined in a similar manner. Specifically, muscle length was manipulated, and single-twitch force production was observed. The length and voltage at which a single twitch produced the greatest force were used throughout the stimulation protocol. For Pt assessment, the stimulating voltage was set to produce a maximum contraction using a gradual increase in the square-wave pulses from 0 to 10 V until producing maximum contraction for 1 ms duration and 20 Hz frequency. Recording was displayed at sampling rate 1K/sec. For PTT assessment, the left gastrocnemius muscles were stimulated by 15 supramaximal pulses, 1 ms duration with a gradual increase in frequency (from 20 to 50 Hz) until the muscle was completely tetanized, and HRT was assessed from the maximum to half the maximum peak tension at muscle optimal length during the relaxation phase of the twitch. Muscle temperature was monitored and maintained at 35°C–37°C, while the muscle was kept hydrated with saline. Data were collected through an AD Instruments Bridge Amp and Powerlab 4/25 and were analyzed with Chart5 PowerLab software for Windows. The tissues were immediately collected and either snap frozen or mounted on cork using mounting medium and quick frozen in isopentane cooled by liquid nitrogen. Samples were maintained at −80°C until further analysis for detection of gene expression.
PCR analyses for RT-PCR
30 mg of gastrocnemius muscle was collected at 67 days after vector infusion and snap frozen in liquid nitrogen. Total RNA was extracted using SV Total RNA Isolation System (Promega) and reverse transcripted. Optimal primer and cDNA template concentrations were determined by titration. Amplification efficiencies for each primer set were determined to be similar. Data were represented as differences between threshold cycle values (ΔCT) for the transcripts of interest and the internal standard, β-actin. The expression of the muscle genes encoding for SERCA, GLUT4, and adiponectin was examined using semi-quantitative RT-PCR. Oligonucleotide sequences are available on request.
Statistical analyses
Differences between groups were determined by F test, Anova, and Post hoc test. Data are expressed as means±standard errors (SEM).
Results
Effect of Adiponectin gene transfer therapy and/or exercise on diabetic state
The use of a high fat diet and streptozotocin resulted in an insulin-resistant diabetic model (DG group) that exhibited hyperglycemia, hyperinsulinemia, and increased levels of triglycerides and cholesterol in comparison to control rats. Treatment with either adiponectin gene or exercise or both resulted in a significant decrease in all these parameters compared with the DG group. However, the treatment with either therapy did not improve the values of these parameters to the control values. With regard to HOMA-IR, serum triglycerides, and cholesterol levels, the combination between adiponectin gene and swimming exercise resulted in a significant reduction in these parameters compared with DEG and in only serum cholesterol level when compared with DAG. (Table 1).
Data are means±SEM.
(p<0.05) versus CG.
(p<0.05) versus DG.
(p<0.05) versus DAG.
(p<0.05) versus DEG.
Control, nondiabetic male albino rats; CG, control group; DG, type 2 diabetic male albino rats; DAG, type 2 diabetic male albino rats treated by adiponectin gene therapy; DEG; type 2 diabetic male albino rats following a regular swimming exercise regimen; DAEG, type 2 diabetic male albino rats treated by adiponectin gene therapy and following a regular swimming exercise regimen.
Effect of Adiponectin gene transfer therapy and/or exercise on wt gain%
Induction of T2DM led to a significant increase in wt gain% compared with CG. Adiponectin gene therapy or exercise resulted in a significant reduction in wt gain% compared with DG but still not reaching the control values. Meanwhile, the combination therapy resulted in a significant decrease in wt gain% compared with the control. (Table 2).
Data are means±SEM.
(p<0.05) versus CG.
(p<0.05) versus DG.
(p<0.05) versus DAG.
(p<0.05) versus DEG.
Effect of Adiponectin gene transfer therapy and/or exercise on muscle contraction parameters
Pt and PTT were significantly decreased in DG with a prolongation of HRT compared with CG. Treatment with either adiponectin gene or performing swimming exercise elevated both Pt and PTT with shortening of HRT compared with DG. However, the present results elucidated that adiponectin gene therapy exerted a better protective role than exercise. Furthermore, the cotreatment using both therapies not only caused a significant increase in Pt and shortening of HRT compared with DG but also resulted in a greater significant increase in Pt and more shortening in HRT than CG. Therefore, the current data showed that the best protective effect was offered by the combination therapy. (Figs. 1 –3).
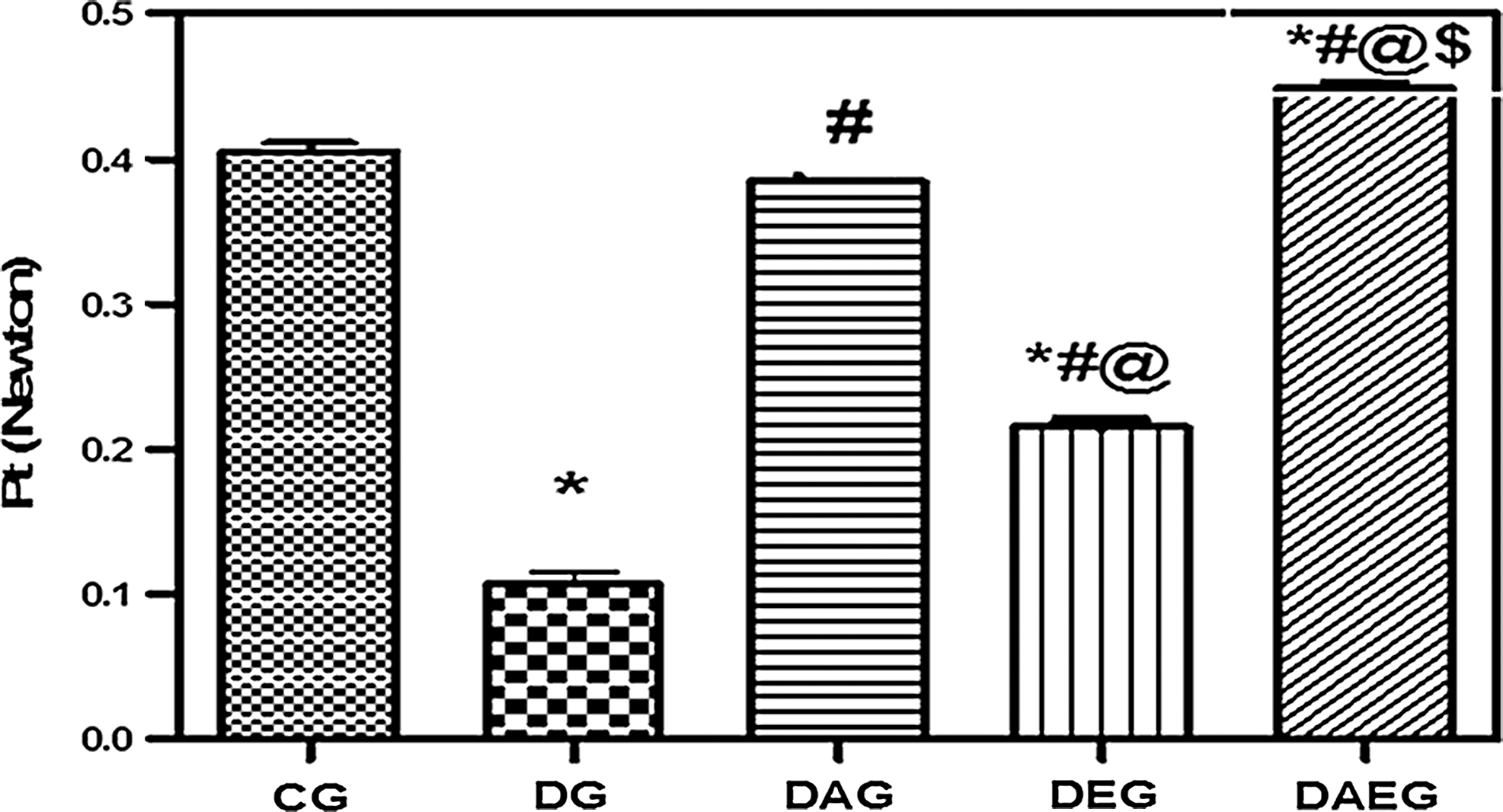
Peak Twitch Tension (Pt) in five studied rat groups. Pt in five studied rat groups. DG was impaired in Pt compared with the control. Both Adiponectin gene therapy and regular swimming exercise regimen restored Pt in diabetic rats. However, adiponectin gene therapy provided a better Pt compared with regular swimming exercise regimen. Combined therapy restored Pt to the control values. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, ‖(p<0.05) versus DAG, $(p<0.05) versus DEG.
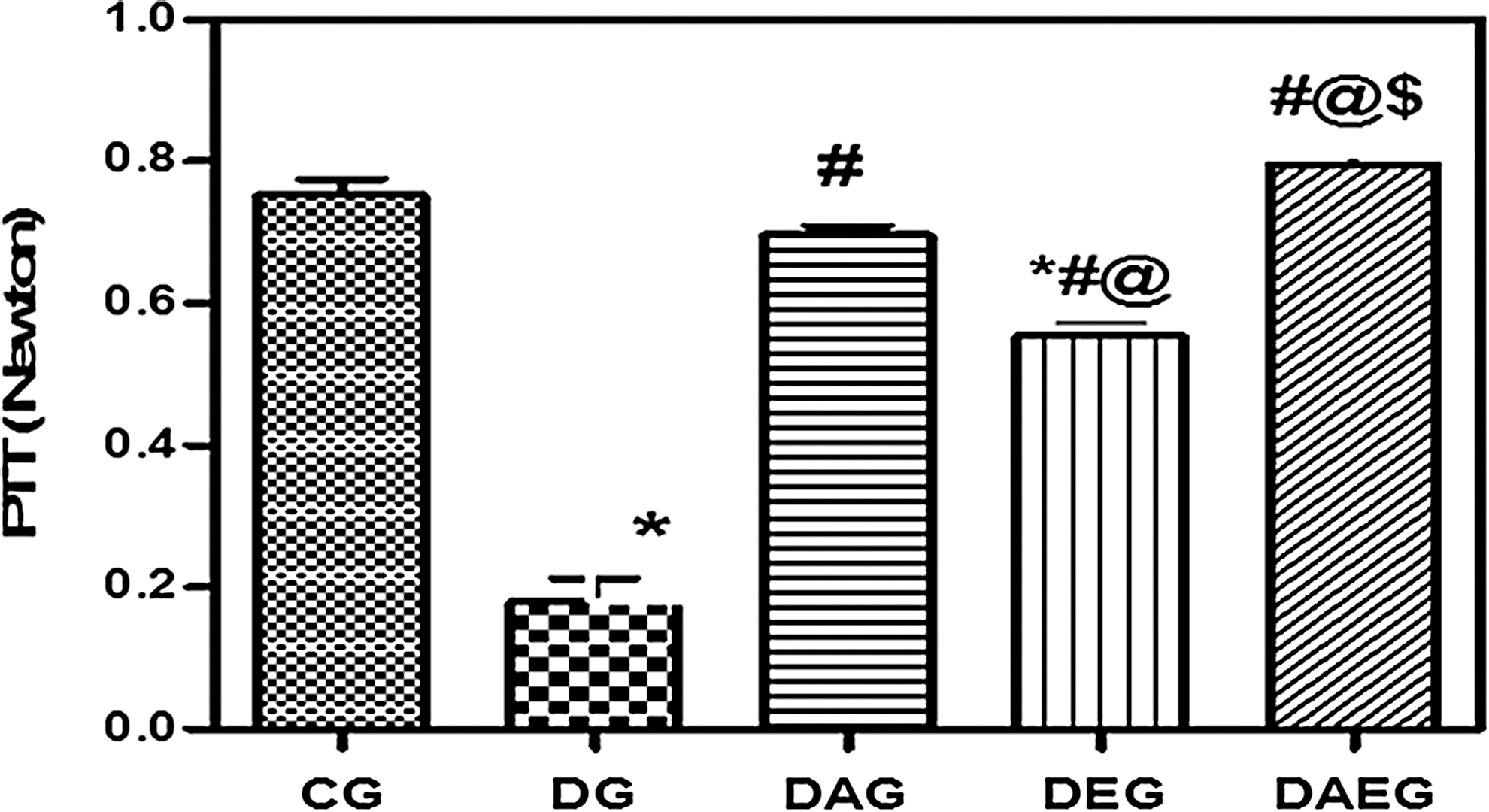
Peak Tetanic Tension (PTT) in five studied rat groups. DG had impaired in PTT compared with the control. Both Adiponectin gene therapy and regular swimming exercise regimen restored PTT in diabetic rats. However, adiponectin gene therapy provided a better PTT compared with regular swimming exercise regimen. Combined therapy restored PTT to the control values. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, ‖(p<0.05) versus DAG, $(p<0.05) versus DEG.
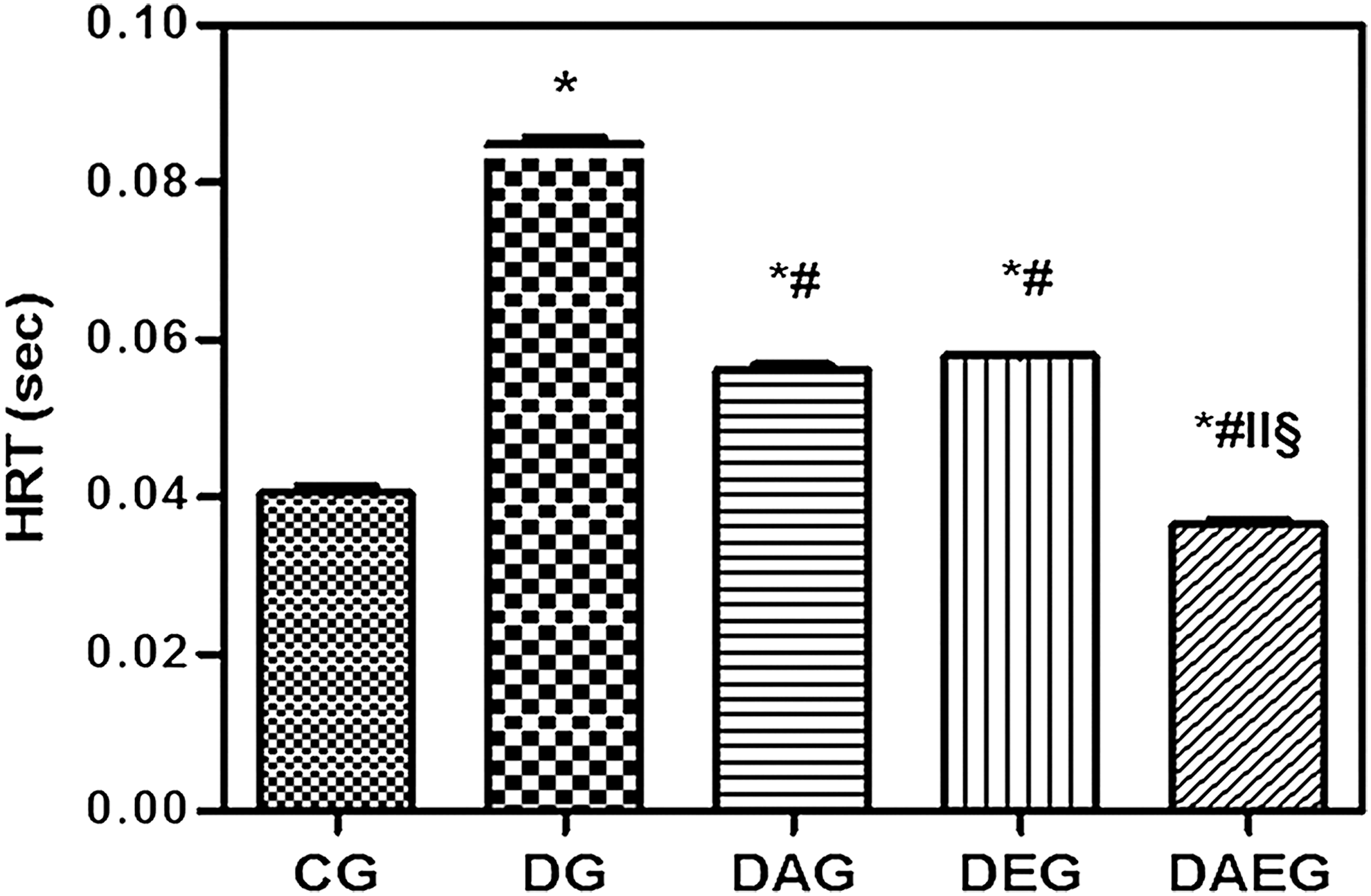
Half Relaxation Time (HRT) in five studied rat groups. HRT in five studied rat groups. DG had prolonged in HRT compared with the control. Both Adiponectin gene therapy and regular swimming exercise regimen shortened HRT in diabetic rats. Combined therapy shortened HRT to the control values. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, ‖(p<0.05) versus DAG, §(p<0.05) versus DEG.
Effect of Adiponectin gene transfer therapy and/or exercise on muscle gene expression
SERCA, GLUT4, and adiponectin muscle gene expressions were significantly lower in DG than in CG. Either adiponectin gene therapy and/or exercise significantly increased expression of these genes compared with DG. Nevertheless, combination therapy proved to have a better protective role concerning SERCA and GLUT4 than the use of adiponectin gene as monotherapy. (Figs. 4 –6).
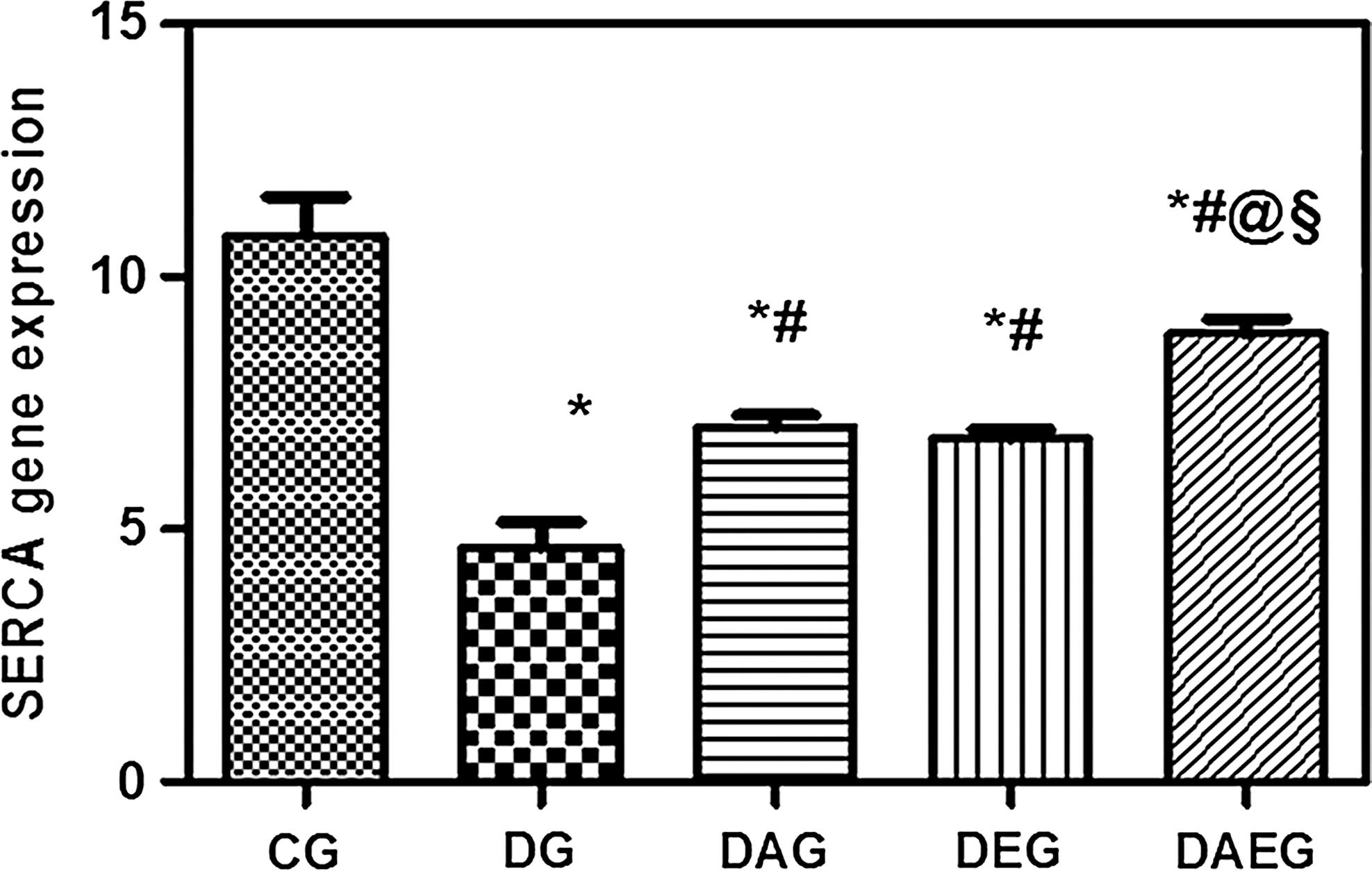
SERCA muscle gene expression in five studied rat groups. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, ‖(p<0.05) versus DAG, §(p<0.05) versus DEG.

GLUT4 muscle gene expression in five studied rat groups. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, ‖(p<0.05) versus DAG, §(p<0.05) versus DEG.

Adiponectin muscle gene expression in five studied rat groups. Data are means±SEM, *(p<0.05) versus CG, #(p<0.05) versus DG, §(p<0.05) versus DEG.
Discussion
T2DM is one of the most common metabolic disorders, representing a major health issue worldwide (Narayan et al., 2000). Since skeletal muscles are the primary sites of insulin-dependent glucose disposal (Ferrannini et al., 1999), thus, resistance of the skeletal muscles to insulin-dependent glucose uptake may be an early step in the development of T2DM (Cline et al., 1999).
In the present study, the administration of a high fat diet and STZ injection in DG produced a significant increase in serum glucose, insulin, HOMA-IR, serum triglycerides, and cholesterol levels compared with CG, indicating the development of insulin resistance.
In the present study, the muscle mechanical contractile performance was assessed. Pt and PTT were recorded. Pt reflects the strength of the force and the contractile activity of the muscle, whereas PTT signifies that impairment in force generation in fibers on removal of external Ca2+ is due to impaired SR Ca2+ release (Payne et al., 2004). DG showed a significant decrease in Pt and PTT to control values. The reduction in Pt and PTT is in agreement with several studies, including that of Paulus and Grossie (1983), who recorded a marked decrease in twitch and tetanic forces in uncontrolled diabetic rats (Paulus and Grossie, 1983). Several mechanisms were involved to explain this deterioration. The polyol pathway was implicated and was driven by both hypoinsulinemia and hyperglycemia (Cameron et al., 1989, 1990). In addition, the hypoinsulinemia was considered a secondary factor causing atrophy, particularly of fast muscles (Cameron et al., 1990).
In the present work, HRT was also measured. HRT depends on the uptake of Ca2+ by the SR (Gollnick et al., 1991), the rate of dissociation of cross-bridges (Crow and Kushmerick, 1983; de Haan et al., 1989), and the free energy of hydrolysis of ATP (Dawson et al., 1980; Russell et al., 1984). The current results showed that HRT was prolonged in the untreated diabetic rats. Cotter and his co-workers (1993) reported similar results. They attributed the prolongation of HRT to the polyol accumulation that causes swelling of SR tubules and SR damage (Cotter et al., 1993).Current results revealed a significant decrease in the gastrocnemius muscle SERCA gene expression level in DG. Therefore, such an alteration in SERCA gene expression and the resulting decrease in Ca2+ uptake by SR could explain the prolongation of HRT in DG in this study.
Depletion of GLUT4 in either adipose tissue or skeletal muscle causes insulin resistance (Zisman et al., 2000; Abel et al., 2001). Therefore, in the present work, the reduction in the muscle GLUT4 gene expression in DG confirmed the insulin resistance and maybe one of the causes of the reduction in Pt and PTT, as it is associated with a decrease in glucose uptake, a reduction in the energy and fuel in the muscle, resulting in the weak contraction obtained.
Treatment with adiponectin gene therapy resulted in an improvement in the diabetic state. However, these group values did not reach the control values, suggesting partial improvement in these parameters. Another study showed similar results to our own. Adiponectin administration in both wild-type (WT) and diabetic animal models revealed an enhancement in insulin sensitivity and corrected metabolic abnormalities (Yoon et al., 2006). Yamauchi et al. (2002) proved that these adiponectin effects are mediated through AMPK activation. Activation of PPAR-α, a further important factor, that regulates the expression of genes involved in glucose and lipid metabolism was also identified to mediate the insulin sensitizing action of adiponectin (Kadowaki and Yamauchi, 2005).
With regard to the gastrocnemius muscle contractile performance, DG treated with the adiponectin gene showed a significant increase in Pt and PTT compared with the untreated DG, with no significant difference with the control values, suggesting complete improvement in the muscle force after gene therapy. The present study is in agreement with the work of Krause and his co-workers (2008), who assessed the muscle contractile properties, where PTT was 50% lower in adiponectin-KO (Ad-KO) mice compared with WT mice. Furthermore, this study revealed a shortening in HRT in DG on gene therapy, suggesting an increase in Ca2+ uptake by the SR (Krause et al., 2008) (Table 3).
Data are means±SEM.
(p<0.05) versus CG.
(p<0.05) versus DG.
(p<0.05) versus DAG.
(p<0.05) versus DEG.
Pt, peak twitch tension; PTT, peak tetanic tension; HRT, half relaxation time.
In addition, the present study proved an increase in the muscle SERCA, GLUT4, and adiponectin gene expressions in the treated DG with the adiponectin gene. Thus, the shortening of HRT can be explained by the significant increase in the SERCA gene expression and, consequently, the increase in the Ca2+ uptake by SR. It has been postulated that adiponectin may attribute to the enhanced glucose uptake in skeletal muscle cells (Berg et al., 2001) via increasing GLUT4 expression and translocation (Ceddia et al., 2005) (Table 4).
Data are means±SEM.
(p<0.05) versus CG.
(p<0.05) versus DG.
(p<0.05) versus DAG.
(p<0.05) versus DEG.
SERCA, sarco and endoplasmic reticulum Ca+2 ATPase.
In the present study, we examined the effects of a 9 weeks daily swimming exercise regimen on T2DM rats. DG following the swimming exercise regimen showed a significant reduction in the measured serum parameters compared with DG. However, this same group showed a significant elevation in all these measured parameters compared with CG, suggesting a partial protection.
Pt and PTT showed a significant increase in the trained DG compared with the untrained DG and a significant decrease compared with CG. Moreover, this same group showed a shortening in HRT compared with DG (Table 3).
Moreover, the trained DG revealed a significant increase in the muscle SERCA, GLUT4, and adiponectin gene expressions. This coincides with other studies which indicated that both moderate- and high-intensity exercise significantly increase the SERCA2a expression in the gastrocnemius (Kubo et al., 2003). Exercise training has profound effects on PPAR-γ activation in rats (Petridou et al., 2007). Activation of PPAR-γ affects GLUT4 (Berger and Moller, 2002), activates AMPK (Saha et al., 2004), and improves insulin action (Saltiel and Olefsky, 1996). Since PPAR-γ activation is also associated with increased adiponectin production (Tsuchida et al., 2005), activation of PPAR-γ with exercise represents a possible mechanism that links the insulin sensitizing effects of exercise with adiponectin (Table 4).
Furthermore, the combination of adiponectin gene therapy and exercise resulted in a significant reduction in all the serum parameters.
The present work also revealed a significant elevation in Pt and PTT with a shortening in HRT in DG in the combined therapies compared with both DG and CG, suggesting full improvement in the muscle contractile performance by the combination therapy (Table 3).
Finally, the combination therapy group showed a significant increase in the gastrocnemius muscle SERCA, GLUT4, and adiponectin gene expressions. An interesting observation in the previous studies was that both exercise and adiponectin shared somehow similar insulin sensitizing effects. Both promoted skeletal muscle glucose uptake, increased fatty acid oxidation, and modulated GLUT4 translocation and AMPK activity (Ren et al., 1994; Tomas et al., 2002; Wright et al., 2004; Ceddia et al., 2005) (Table 4).
Conclusions
To our knowledge, this is the first study linking SERCA and skeletal muscle contractile performance in a T2DM insulin resistant animal model. There was alteration in SERCA, GLUT4, and adiponectin genes expression in the gastrocnemius muscle associated with contractile dysfunction. Adiponectin gene therapy or swimming exercise, by increasing the expression of these genes, exerted a partial protective effect on the insulin resistance state and the skeletal muscle contractile performance. However, the use of a combination therapy resulted in the best protection against the skeletal muscle deterioration. Taken together, these data support the potential use of such a combination as a promising strategy in the treatment of T2DM muscle dysfunction.
Recommendations
Further investigations on the role of adiponectin gene therapy and exercise training in other pathophysiologic pathways involved in diabetic skeletal muscle dysfunction are recommended. In addition, the trial of various types of exercise protocols is recommended in order to select the one that provides the best protection available.
Footnotes
Disclosure Statement
No competing financial interests exist.