
Abstract
The aim of this study was to evaluate the influence of the γ-aminobutyric acid receptor type A β-3 subunit (GABRB3) polymorphisms in patients with nonsyndromic cleft lip and/or palate (NSCL/P). We carried out a structured case–control analysis of three GABRB3 polymorphisms (rs4477673, rs6576618, and rs981778) in 229 patients with nonsyndromic cleft lip with or without cleft palate (CL±P) and in 314 unaffected controls from Brazil. The polymorphisms were genotyped by the TaqMan 5′-exonuclease allelic discrimination assay, and each sample was independently typed for 40 biallelic short insertion/deletion markers (INDELs) to characterize the genomic ancestry. The genotype distributions of the three polymorphisms were as expected by the Hardy–Weinberg equilibrium test. After adjustment to ancestry contribution, the minor A allele of rs981778 was associated with NSCL/P, but significant results did not persist after Bonferroni correction for multiple tests. Similarly, the haplotype analysis revealed that the CCA haplotype (C allele of rs4477673, C allele of rs6576618, and A allele of rs981778) was correlated with NSCL/P, but this association did not remain statistically significant after Bonferroni correction. With a weak association, our data do not support the hypothesis that the GABRB3 variants are a cause of NSCL/P, but further studies are warranted.
Introduction
γ-Aminobutyric acid receptor type A β-3 subunit (GABRB3), one of the four members of the GABAA receptor family, is the primary mediator of the fast inhibitory synaptic transmission in the central nervous system (Hagiwara et al., 2003). GABAA receptors comprise 2 α-subunits, 2 β-subunits, and 1 γ- or δ-subunit (Bormann, 2000). Mice that lack the β-3 subunit of the GABRB3 gene exhibited severe neurological impairment characterized by hyperactivity, lack of coordination, and seizures (Homanics et al., 1997). In addition, these animals showed oral clefts, which were identified as the principal cause of death that occurs in ∼90% of the newborns within 24 h (Homanics et al., 1997). As mutations in GABRB3 were described in human epilepsy syndromes and genetic polymorphic variants were associated with isolated forms of epilepsy (Lachance-Touchette et al., 2010; Macdonald et al., 2010), it is plausible that the polymorphisms in GABRB3 are associated with the pathogenesis of NSCL/P. Indeed, previous studies provided bodies of evidence of an association between the GABRB3 variants and human oral clefts (Scapoli et al., 2002; Inoue et al., 2008; Vieira et al., 2008). In a large case–control study, including samples from the Philippines and Iowa, Vieira et al. (2008) evaluated nine SNPs in the GABRB3 gene, revealing a possible association of three of them (rs4477673, rs6576618, and rs981778) with NSCL/P. Herein, we have performed a case–control study to investigate whether those genetic variations of the GABRB3 gene are associated with NSCL/P in the Brazilian population. However, as the Brazilian population displays very high levels of genomic diversity, the result of five centuries of mating between Amerindians, Europeans, and sub-Saharan Africans (Parra et al., 2003) that may have important implications on NSCL/P susceptibility, we performed a structured genetic analysis in which the genomic ancestry variation of each individual was taken into account.
Materials and Methods
Samples
A total of 229 patients with NSCL/P and 314 healthy controls were included in this study. Patients with NSCL/P were recruited at the Center for Rehabilitation of Craniofacial Anomalies, Dental School of Alfenas, Minas Gerais, State, Brazil. The controls were chosen among healthy subjects admitted as in-patients for regular dental treatment in the Dental School of the same University with conditions unrelated to clefting disorders (volunteers with congenital malformations or with a history of familial oral clefting were not included). Participants were recruited between 2008 and 2011, and all subjects were born in the study area, which involves 156 cities in the South of Minas Gerais State, Brazil. To confirm the NSCL/P diagnosis, all patients were carefully examined and screened for the presence of associated anomalies or syndromes by the team of the Center for Rehabilitation of Craniofacial Anomalies. The nonsyndromic clefts were classified with the incisive foramen as a reference, and 82 patients had cleft lip (CL), and 147 had cleft lip and palate (CLP). Written informed consents were obtained, and the study was carried out with approval of the Human Research Ethics Committee of the University.
Genotyping
To test the association between GABRB3 and NSCL/P, genomic DNA was extracted from oral mucosa cells and genotyped for three SNPs (Table 1). These polymorphisms were selected based on their minor allele frequencies close to 0.3 (
Reference:
GABRB3, γ-aminobutyric acid receptor type A β-3 subunit.
Assessment to genomic ancestry
Each sample was independently genotyped for 40 biallelic short insertion–deletion polymorphisms (INDELs) after the methods of Bastos-Rodrigues et al. (2006).
Data analysis
Deviation from the Hardy–Weinberg equilibrium in the control group was assessed through the chi-squared test. To determine the genomic ancestry of each individual, Structure software (Falush et al., 2007) was utilized in a model assuming that K=3 parental populations based on the trihybrid origin of the Brazilian population. To assist the software in the estimation of the ancestry, the reference samples of the European, African, and Amerindian ancestry from Marshfield Clinic collection were included. After ancestry assessment, STRAT was used to test the association, taking into account the individual ancestry proportions (Pritchard et al., 2000). The odds ratio and associated 95% confidence intervals (95% CI) were also calculated. The genotypes were analyzed under unrestricted, dominant, and recessive genetic models. The haplotype frequencies and pairwise linkage disequilibrium (D′ and r 2) were estimated using HaploView software (Barrett et al., 2005). The Bonferroni correction for multiple comparisons was applied, and the corrected p-value of ≤0.016 (0.05/3) was considered statistically significant.
Results
Figure 1 depicts the proportions of the Amerindian, European, and African ancestry of each sample. The average ancestry contributions were estimated at 84.4% of European, 11.4% of African, and 4.2% of Amerindian in the control group, and in the NSCL/P group was 82.5% of European, 15% of African, and 2.5% of Amerindian.
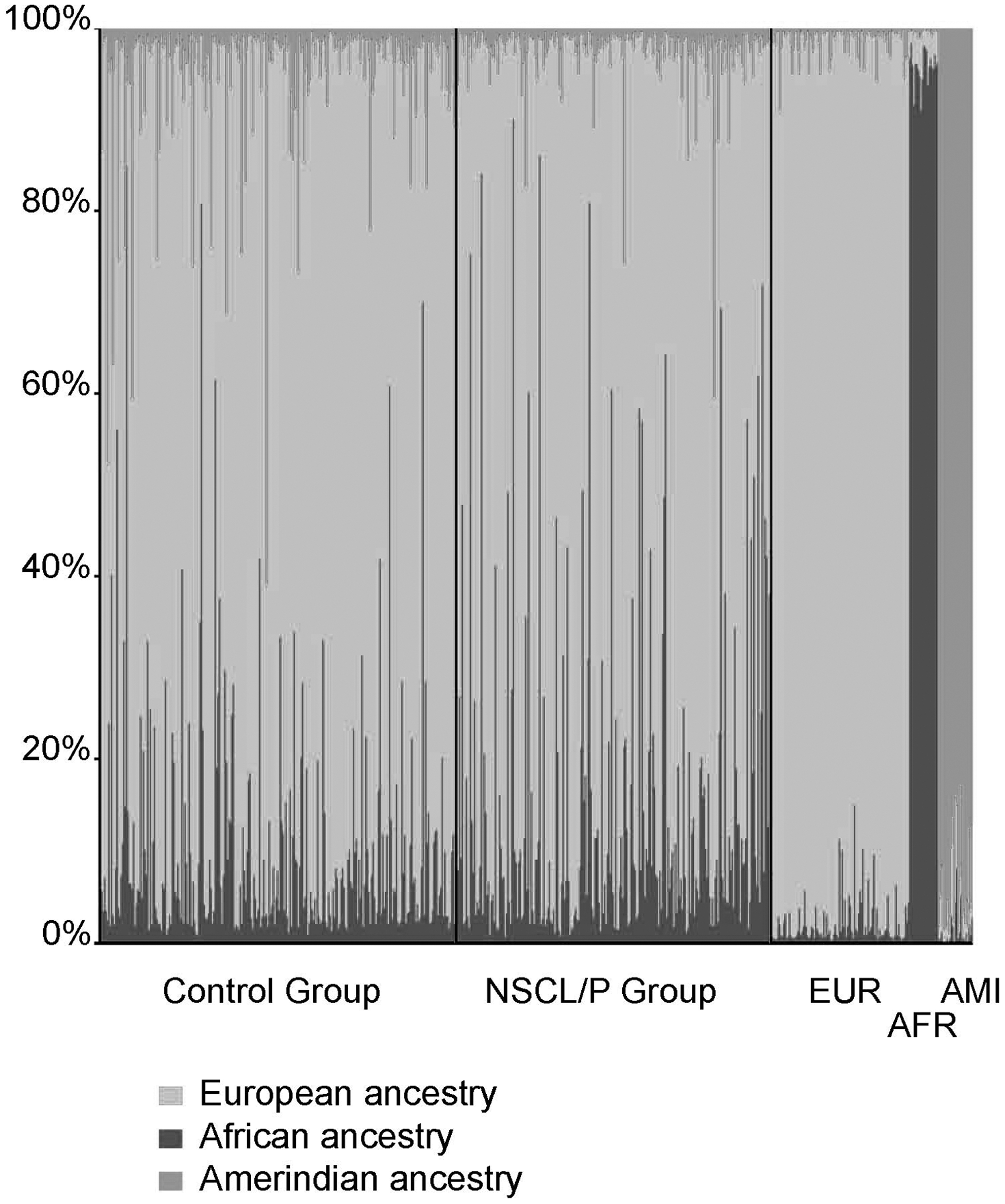
Genomic proportions of the European, African, and Amerindian ancestry in the control and nonsyndromic cleft lip and/or palate (NSCL/P) groups. Each individual is represented by a single column, and the columns identified as EUR (European), AFR (African), and AMI (Amerindian) represent the parental populations used to assist the structure in estimating ancestry of the admixed individuals.
The allele and genotype distributions of the polymorphisms are presented in Table 2, and their derived haplotypes are presented in Table 3. The genotype frequencies observed for all studied polymorphisms in controls did not reveal statistically significant differences compared to those expected under the Hardy–Weinberg equilibrium. A structured association test showed that the minor allele frequencies and the genotype distributions of rs4477673 and rs6576618 were similar in the control and NSCL/P groups. The minor allele frequency of rs981778 (allele A) was more frequent in the NSCL/P group, and reached a significant level in the CL group (p=0.03; Table 2). However, this significance did not remain after Bonferroni correction for multiple comparisons. Further, the analysis in the dominant and recessive genetic models also revealed no differences in the genotype distribution between groups (data not shown).
HWE, Hardy–Weinberg equilibrium; CL±P, cleft lip with or without palate; CL, cleft lip; CLP, cleft lip and palate;
Order of the polymorphisms: rs4477673, rs6576618, and rs981778.
The GABRB3 polymorphisms were in the linkage disequilibrium (D′=0.68 and r 2=0.51). Eight haplotypes were observed in our cohort, and carriers of the CCA haplotype (C allele of rs4477673, C allele of rs6576618, and A allele of rs981778) were found to be more prevalent among all types of cleft as compared to controls, but this difference was not kept after Bonferroni correction (Table 3).
Discussion
NSCL/P is a common congenital anomaly with significant medical, psychological, and social impacts. Substantial evidence indicates that this anomaly represents a complex disease in which clinical and genetic heterogeneities are observed (Rahimov et al., 2012). The genetic basis of oral clefting remains an open question, and the identification of risk factors for NSCL/P has been the subject of intensive research. On these grounds, we investigated the association between the GABRB3 genetic variants and NSCL/P, since the members of the GABBA system are promising candidates for the pathogenesis of oral clefting (Homanics et al., 1997; Hagiwara et al., 2003; Ding et al., 2004; Oh et al., 2005). Although we have identified an association of the rs981778 A allele with CL and of the CCA haplotype with both CL and CLP, those associations disappeared after controlling for multiple comparisons.
Previous studies have investigated the role of the GABRB3 variants in orofacial clefts, but the results have been inconsistent. The first study was performed by Tanabe et al. (2000) with one group of Japanese patients affected by NSCL/P. Those authors evaluated the CA dinucleotide-repeat polymorphism in GABRB3 and concluded that the GABRB3 gene is not involved with the pathogenesis of oral clefts in Japanese patients, but it should be taken into account that their results were not submitted to any statistical analysis. Later, Scapoli et al. (2002) investigated the same CA dinucleotide-repeat marker in 38 Italian families with NSCL/P and showed significant evidence of the linkage disequilibrium. Individually, SNPs in the GABRB3 gene were not associated with NSCL/P in a second study with Japanese patients, but one 6-locus haplotype of the gene was significantly more frequent in the patients with NSCL/P than in the controls, revealing the involvement of the GABRB3 gene in the pathogenesis of NSCL/P in the Japanese population (Inoue et al., 2008). Vieira et al. (2008) analyzed nine SNPs in the GABRB3 gene in the populations from Iowa and Philippines and demonstrated an association between three of them (the same used in the current study) and NSCL/P. As in the present study, a weak association between the GABRB3 polymorphisms and NSCL/P was reported with different polymorphic markers in the populations from Lithuania and Ireland (Morkuniene et al., 2007; Carter et al., 2010).
These inconsistent results that characterize the role of GABRB3 in NSCL/P pathogenesis may be related to the intrinsic genetic heterogeneity of the disease or to the low penetrance of the gene. In support to the later, recent genome-wide association studies (GWAS), which are based on comparison of multiple common polymorphisms between cases and controls, have not identified GABRB3 as a candidate gene in association with an increased or decreased risk to NSCL/P development (Grant et al., 2009; Marazita et al., 2009; Beaty et al., 2010; Mangold et al., 2010; Ludwig et al., 2012). Other possible reason to the lack of association in the current study includes the number of available samples, particularly with regard to the CL number; therefore, modest associations of polymorphisms and the oral cleft risk may have been missed. In addition, Bonferroni correction is too stringent, and led to a loss of the significant findings, but in light of not having replication in a second cohort, this approach appeared to be the most efficacious.
Conclusion
Although we identified an association of the GABRB3 polymorphisms with NSCL/P, no statistically significant differences resisted to Bonferroni correction. Considering the modest association in a cohort with more than 80% of European ancestry, it is likely that GABRB3 might track a functionally relevant role in the European haplotypic block as a low-penetrant gene. Further analysis of larger groups of controls and NSCL/Ps is needed to clarify the underlying role of the GABRB3 polymorphisms in NSCL/P, which may overcome the limitations of the present study.
Footnotes
Acknowledgments
This work was supported by the grants from São Paulo Research Foundation-FAPESP, São Paulo, Brazil, National Council for Scientific Technological Development-CNPq, Brasília, Brazil, and Fundação de Amparo a Pesquisa do Estado de Minas Gerais-FAPEMIG, Belo Horizonte, Brazil. MRF was supported by PIBIC-CNPq.
Disclosure Statement
No competing financial interests exist.