
Abstract
Mycobacterium bovis, the classical causative agent of bovine tuberculosis (BTB), infects animals of agricultural importance and other mammals, including humans. Neutrophils are one of the first lines of defense against all microbes and produce a diverse collection of antimicrobial molecules, which play an important role in the early control of tuberculosis progression. An interferon (IFN)-inducible neutrophil-driven blood transcriptional signature that consisted of both IFN-γ and type I IFN-α/β signaling has been identified in human tuberculosis, supporting a role for neutrophils in the pathogenesis of tuberculosis disease. However, it is unknown whether bovine neutrophils play a similar role during M. bovis infection. Thus, we assessed the expression levels of ten IFN-inducible transcriptional genes in neutrophils from healthy cattle stimulated by M. bovis and neutrophils isolated from three groups of cattle of different infection status, and in addition, examined the changes in the expression of myeloperoxidase (MPO) and pentraxin-related protein pentraxin-inducible protein (PTX3) genes during bovine tuberculosis infection. Our results demonstrated a specific expression pattern of IFN-inducible transcriptional genes and MPO and PTX3 genes in neutrophils during bovine tuberculosis infection. The observed expression pattern provides a potential diagnostic tool, which may have implications for vaccine and therapeutic development to combat the bovine tuberculosis epidemic.
Introduction
Tuberculosis can be considered the prototype of chronic infectious diseases in which the most important pathogenic factor is the balance between protection and tissue damage that is mediated by the immune response (Rook and Hernandez-Pando, 1996). The immune response to M. tuberculosis is complex and incompletely characterized, hindering development of new diagnostics, therapies, and vaccines. The early preclinical stages of bovine tuberculosis can be detected in live animals by the use of tests of cellular-mediated immunity, such as the tuberculin skin test and gamma-interferon (IFN) assay. None of the tests currently available for routine diagnosis of tuberculosis in cattle provides a completely accurate determination of infectious status (De la Rua-Domenech et al., 2006).
IFNs are pleiotropic cytokines that can mediate anti-viral responses, inhibit proliferation, and participate in immune surveillance and tumor suppression by inducing the transcription of a number of IFN-stimulated genes. The IFN family includes two main classes of related cytokines, type I IFNs and type II IFN. There are many type I IFNs: IFN-α, IFN-β, and many others. By contrast, there is only one type II IFN, IFN-γ, which is produced by activated T cells and natural killer cells (Platanias, 2005). The absolute requirement for IFN-γ in the immune control of TB is well established in humans (Dupuis et al., 2000). Though IFN-γ-dependent protection is commonly believed to be mediated by increasing the mycobactericidal activity of macrophages, recent work has suggested additional important regulatory roles for IFN-γ during M. tuberculosis infection (Nandi and Behar, 2011). The latest research demonstrated that type I IFNs play a non-redundant protective role during the early stages of TB in vivo, and the effects of type I IFNs are intricately connected to, and ultimately masked by, the influence of IFN-γ, especially after the onset of adaptive immunity (Desvignes et al., 2012).
Neutrophils or polymorphonuclear cells (PMN) form the first line of defense of the animal innate immune system (Borregaard, 2010). Traditionally, it was believed that the main role for neutrophils was in the efficient elimination of extracellular pathogens, but several results also imply the participation of neutrophils in the elimination of intracellular bacterial pathogens, such as Mycobacterium tuberculosis (Silva, 2010). Besides, they are the first inflammatory cells to arrive at sites of infection and present a diverse collection of antimicrobial molecules, which can be used as powerful weapons to defend against and kill all foreign microorganisms, including M. bovis (Cassatella, 1999).
Berry et al. identified a transcriptional signature involving 86 genes in blood neutrophils from patients infected with M. tuberculosis, which consisted of transcripts that are induced by type I and type II IFNs. Their results also support a role for neutrophils in the pathogenesis of tuberculosis disease, which may result from over-activation by IFN-γ and type I IFNs (Berry et al., 2010). Consistent with the notion that neutrophils are specifically involved in the immune response to M. tuberculosis, in a mouse model of M. tuberculosis infection, neutrophils were required for the production of early innate immune-derived IFN-γ (Pedrosa et al., 2000). Thus, mouse and human studies suggest a crucial role for neutrophils in immunity against the prototypical intracellular pathogen M. tuberculosis. However, there is shortage of data regarding the precise role of neutrophils in the immunity or pathogenesis of tuberculosis, and their exact participation in bovine tuberculosis is still poorly understood.
To better understand the immune response against M. bovis in cattle and the role that neutrophils may play in the progression of bovine tuberculosis, we investigated the expression of ten IFN-inducible transcriptional genes in neutrophils isolated from peripheral blood of healthy cattle stimulated by the M. bovis Beijing strain and in neutrophils isolated from three groups of cattle of different infection status, respectively. Our results showed an interesting pattern of expression of IFN-inducible transcriptional genes in neutrophils during bovine tuberculosis infection.
Materials and Methods
Bacterial strains and culture
Virulent M. bovis Beijing strain (bovis strain 93006) was purchased from the China Institute of Veterinary Drug Control (CVCC). The bacteria were grown at 37°C under shaking conditions, reaching a logarithmic phase in Middlebrook 7H9 broth (Difco) supplemented with 0.05% Tween80 and 10% oleic acid-albumin-dextrose-catalase enrichment (Difco West Molesey) (Wang et al., 2011). For neutrophil infections, log-phase cultures were centrifuged and washed with phosphate-buffered saline (PBS), and bacterial clumps were dispersed by passing them through a 26-gauge needle.
Animals
The blood samples were from Holstein cow herds in Shandong province, China. On the basis of the skin-fold thickness response to tuberculin pure protein derivative (PPD) and a whole blood IFN-γ enzyme immunoassay, the cattle were divided into three groups, namely a healthy control group, a M. bovis-infected cattle group, and a group with evidence of coinfection (shown in Supplementary Table S1; Supplementary Data are available online at
Tuberculin skin test
Cattle were tested by a delayed-type hypersensitivity reaction after an intradermal injection of PPD in the neck. The thickness of the skin was measured and recorded with calipers before applying the tuberculin and 72 h after the injection. A thickness that was 4 mm greater than the avian reaction was considered a positive reaction for M. bovis.
Whole blood IFN-γ enzyme immunoassay
The blood level of IFN-γ was measured by enzyme immunoassay. Briefly, whole heparinized blood was mixed in a 24-well culture plate in a 1:1 ratio with RPMI 1640 medium (Invitrogen) and was then stimulated with avian PPD or bovine PPD in 100 mL in three replicates for 72 h. The results were obtained by comparing the mean absorbance values of the nil (PBS) antigen, avian, and PPD samples for each animal. An OD value greater than that of avian PPD and nil (PBS) antigen by more than 0.100 indicated the presence of M. bovis infection.
Bovine peripheral blood neutrophils isolation
Peripheral blood neutrophils were isolated from Acid Citrate Dextrose (ACD) anticoagulated blood of cattle by OptiPrep (Asix-Shield) gradient centrifugation according to the manufacturer's protocol. The mononuclear cell supernatant (including monocytes and lymphocytes) was discarded, the PMN-rich supernatant was collected, and the residual RBCs were lysed by hypotonic lysis. The separated neutrophils were stained by Wright–Giemsa staining (shown in Supplementary Fig. S1) and the purity of neutrophils isolated was more than 90%. Then, neutrophils were resuspended in RPMI 1640 (Invitrogen) supplemented with 10% fetal calf serum (Hyclone), 2 mM L-glutamine, and penicillin (100 U/mL) at 37°C under 5% CO2.
Cell infection with M. bovis
Neutrophils were allowed to adhere to six-well flat bottom plates (Costar, Corning) at 2×106 cells per well and challenged with M. bovis (MOI=5:1) for 3 h at 37°C with 5% CO2, then washed to remove bacteria. Samples were collected at different time dots for gene expression analysis. The challenge of M. bovis was replaced by PBS in the control group.
RNA extraction and complementary DNA preparation
Total RNA was extracted from cells using the SV Total RNA Isolation System (Promega), and reverse transcribed into complementary DNA (cDNA) using a commercial kit (cDNA Synthesis Kit; Fermentas) with oligo (dT) 18 primers in accordance with the manufacturer's instructions.
Evaluation of the mRNA expression of tested genes by real-time RT-PCR
Expression levels of IFN-inducible transcriptional genes were tested with real-time PCR. The H3 histone family 3A gene (H3F3A) was used as a control gene to normalize expression data for target genes. All samples were analyzed in triplicate. Quantitative PCR (q-PCR) was performed using a commercial mix (SYBR Green Master Mix; Bio-Rad) and a thermal cycler (DNA Engine Opticon™ 2 system; MJ Research) with the specific gene primer shown in Table 1. The total volume for q-PCR was 20 μL, comprising 8 μL of water, 0.5 μL of each primer (10 μM), 10 μL Master Mix, and 30 ng of cDNA. Real-time q-PCR data were analyzed using the 2-ΔΔCt Method. The expression levels were normalized to H3F3A, and differences between groups were analyzed by ANOVA and subsequent post-test analysis by SPSS software.
MPO, myeloperoxidase; PTX3, pentraxin-inducible protein.
Results
Expression of IFN-inducible transcriptional genes in neutrophils from cattle with bovine tuberculosis
Based on the skin-fold thickness response to tuberculin PPD and a whole blood IFN-γ enzyme immunoassay test (shown in Supplementary Table S1), the animals were divided into three groups: healthy cattle group, cattle infected with M. bovis group, and cattle with coinfection group. IFN-inducible transcriptional genes, consisting of both IFN-γ (six genes: CXCL10, IFI44, STAT1, TAP2, CD274, and PD-1) and type I IFN-α/β (five genes: IFI6, IRF7, STAT1, STAT2, and OAS1) signaling in neutrophils were examined by real-time PCR among the three groups (n=5 in each group). There were statistically significant expression differences for all the ten examined genes. Eight genes were up-regulated, while two (TAP2 and PD-1) were down-regulated in infected cattle (Fig. 1A, B).

Quantitative RT-PCR assay of mRNA expression of IFN-inducible genes.
Cattle coinfected with M. avium showed an expression pattern similar to that of cattle infected with M. bovis. Among the 10 genes examined, the expression of two genes, namely PD-1 and TAP2, decreased significantly compared with the control cattle, while eight genes showed increasing expression, of which three genes (IFI44, IRF7, and OAS1) were significantly up-regulated. Compared with the group with bovine tuberculosis, cattle with coinfection, infected with M. avium and M. bovis, might not have a similar strong activation effect. However, it is not clear whether M. avium weakened the effect of M. bovis to a certain extent or M. avium itself, as a less virulent Mycobacterium strain has a less potent stimulating activity in comparison with M. bovis.
The up-regulation of the expression of CD274, IFI6, STAT1, STAT2, and OAS1 was much more pronounced in cattle from the BTB group than in cattle from the coinfection group, while the up-regulation of CXCL10 was much more pronounced in the coinfection group. For the other genes, the expression levels in the BTB and coinfection groups were similar.
IFN-inducible transcriptional genes expression increased with persistent M. bovis infection in vitro
It is generally accepted that neutrophils are recruited to the site of mybacteial infection; in order to further explore whether the effect of expression of IFN-inducible transcriptional genes could be induced by the direct interaction between M. bovis infection and neutrophils to mimic the initial stage when M. bovis invading animal, we isolated neutrophils from healthy cattle (n=4) and then examined the levels of expression of IFN-inducible transcriptional genes in response to M. bovis challenge at 6 and 18 h postinfection (Fig. 2A, B). As control, expression levels of IFN-inducible transcriptional genes stimulated by Mycobacterium smegmatis were also tested (shown in Supplementary Fig. S2).

Quantitative RT-PCR assay of mRNA expression of IFN-inducible genes in neutrophils in response to M. bovis infection (
Of the 10 genes tested, expressions of nine increased as infection prolonged, while only one gene, STAT2, decreased significantly, indicating that the induction effect of STAT2 expression was transient. It is interesting to note that genes in neutrophils which had significant changes were included in IFN-γ-inducible genes. Among the IFN-α/β-inducible genes, only STAT2 and the cotranscription factor of type I IFNs and type II IFN STAT1 was significant. The control group of infection with M. smegmatis showed no significant changes in any IFN-inducible genes.
Furthermore, we tested the expression of IFN-α/β and IFN-γ in mRNA level in infected neutrophils (shown in Supplementary Fig. S3A, S3C). The results showed that IFN-γ went up notably as the infection extended, while IFN-α/β remained relatively low. We also tested the IFN-γ in that conditioned medium from infected neutrophils (using the whole blood IFN-γ enzyme immunoassay ELISA kits, shown in Supplementary Fig. S3B), and the result of optical density also showed that the IFN-γ increased as the infection prolonged. The two results might help explain the reason why IFN-γ-inducible transcriptional genes in neutrophils were up-regulated instead of IFN-α/β-inducible transcriptional genes during M. bovis infection in vitro independent of the crosstalk with any other cells.
Compared with the results from cattle with bovine tuberculosis, expression of most of the genes, with the exception of TAP2 and STAT2, demonstrated consistent changes suggesting existence for a different mechanism that might be utilized by neutrophils in the early stage of M. bovis infection in vitro.
MPO and PTX3 expression was regulated notably during M. bovis infection
In addition to the IFN-inducible transcriptional genes, we examined the expression of myeloperoxidase (MPO) and pentraxin-inducible protein (PTX3), two antimicrobial molecules used by neutrophils to kill bacteria and other pathogens. The expression pattern of the two genes showed consistency between experiments in vivo (Fig. 3) and in vitro (Fig. 4) to a limited extent. In neutrophils isolated from cattle infected with M. bovis, MPO expression increased as infection prolonged (Fig. 4A); while PTX3 expression reached its peak in the first hours of contact with M. bovis and then decreased as infection extended (Fig. 4B); however, it was still significantly higher than the control at 18 h postchallenge. PTX3 expression at 6 h postchallenge was extremely high in vitro infection, which implied that PTX3 played an important role during the first interaction between M. bovis and neutrophils.

Quantitative RT-PCR assay of mRNA expression of MPO and PTX3 in neutrophils among three groups (control group: healthy cattle; BTB group: cattle with M. bovis infection; coinfection group: cattle with M. bovis and M. avium infection). Neutrophils were isolated from bovine peripheral blood of each sample and then tested for mRNA expression by quantitative RT-PCR. Asterisks (*) indicate a statistical significance between groups. Comparisons of the tuberculosis-infected group (n=5), the healthy cattle group (n=5), and the coinfection group (n=5) are shown in different bars. *p<0.05. MPO, myeloperoxidase; PTX3, pentraxin-inducible protein.
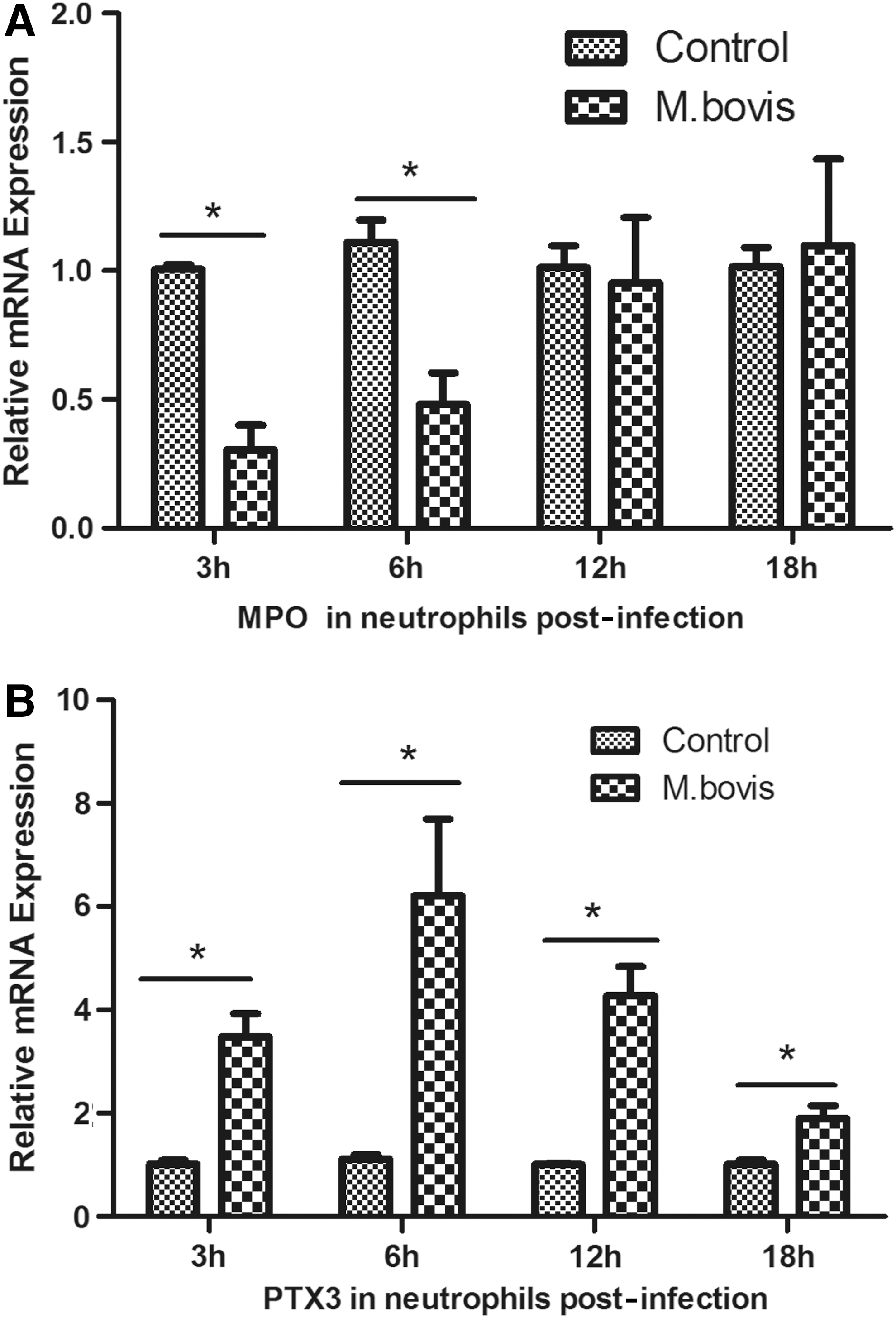
Quantitative RT-PCR assay of mRNA expression of MPO
Discussion
Bovine tuberculosis in animals is primarily associated with cattle and other bovidae, the major causative agent being M. bovis, a member of the M. tuberculosis complex (Smith et al., 2006). The M. bovis Beijing strains causes a substantial proportion of bovine tuberculosis cases worldwide (Kremer et al., 2009). The fact that it also causes human tuberculosis has raised renewed concerns regarding the zoonotic risk for humans, especially those living at the animal–human interface (Michel and van Helden, 2010).
It is well established that neutrophils are one of the first lines of defense when microbes invade and contribute to early defense against mycobacteria. The results of our study showed an up-regulation of IFN-γ-inducible transcriptional genes in neutrophils during M. bovis acute infection. In addition, a difference between IFN-γ and type I IFN signaling gene expressions occurred between infected and control animals. According to the results cited earlier, it may be concluded that type I IFNs contributed to the host defense mechanism during the latent stage of bovine tuberculosis in vivo. Thus, our result also supports a key role of type I IFNs-inducible genes in the host defense mechanism during the long battle between host and bovine tuberculosis in vivo. However, the exact participation and precise role in anti-BTB effect still requires further investigations.
Neutrophils can induce IFN-γ-inducible transcriptional genes that are activated except for direct killing for M. bovis in the early infection stage, when neutrophils were first recruited to and arrived at the infection site. However, in the latent periods, the crosstalk among neutrophils and other types of host immune cells (probably with the lymphocytes that are well known to produce IFN during bovine tuberculosis) can further affect the outcome. Thus, we may say that neutrophils can play a role in the pathogenesis of tuberculosis disease by regulation of IFN-inducible transcriptional genes expression in different infection status and periods.
CD274, also known as programmed death ligand1 (PD-L1), is an immune-modulatory molecule that may also be involved in the IFN signaling pathway (Boasso et al., 2008). The pathway of PD-L1/PD-1 interactions can play a role in a diverse array of settings, including infectious disease. Recent studies implied that this pathway participated in the pathogenesis of chronic infections, including TB. The latest research showed that the expression of PD-L1 increased in whole blood from active TB patients compared with whole blood from healthy controls or M. tuberculosis-exposed individuals, and that expression by neutrophils is largely responsible for this increase (McNab et al., 2011). Our results were in accordance with these recent findings, which suggest that the over-expression of CD274 has a close connection with the defense mechanism against bovine tuberculosis infection. In a recent study, PD-1 KO mice infected with M. tuberculosis succumbed to disease much earlier than wild-type mice, and had severe lung pathology that was associated with a dramatic increase in neutrophils in the lungs (Lázár-Molnár et al., 2010), suggesting that PD-L1 expression acts by limiting neutrophil-mediated immunopathology.
MPO is a peroxidase enzyme that is critical in the oxidative burst, most abundantly expressed and first formed in granulocytes during neutrophil maturation, and stored in azurophilic granules of the neutrophil (Klebanoff, 2005). Previous data indicated that purified MPO, in the presence of hydrogen peroxide, exerts a consistent killing activity against M. tuberculosis H37Rv and also against a clinical isolate in a time- and dose-dependent manner (Borelli et al., 1999). Our results showed that the progression of the infection induced an increase in MPO expression, confirming reports of a possible role of MPO in host defense strategy against the intracellular bacterial infection.
Pentraxin-related protein PTX3 is produced and released by several cell types, including mononuclear phagocytes, dendritic cells, fibroblasts, and endothelial cells in response to primary inflammatory signals (Bottazzi et al., 2009). Our results revealed that at the early stage of bovine tuberculosis infection, PTX3 expression could reach a relatively high level in a short time, but fell considerably as infection prolonged. After neutrophils are activated, a preformed amount of PTX3 is released into the extracellular space, representing a ready-to-use reservoir of PTX3 to guarantee its early release and early activity in acute inflammation caused by stimulus such as M. bovis, but excessive inflammation where increased numbers of activated neutrophils is gathered is harmful to the host (Kunes et al., 2012).
In acute infection with M. bovis, neutrophils have been activated and preformed PTX3 is released into the extracellular space. When inflammation is evoked, those stimuli that account for early PTX3 release concomitantly delay neutrophils' apoptotic death. A prolonged neutrophil lifespan is needed to support the host protection against infection. However, there is a natural feedback mechanism that can inhibit excessive inflammation to a certain required extent. In chronic infection, the role of PTX3 appears to be primarily protective, which can dampen the inappropriate, exaggerated inflammatory response. MPO has been believed to be critical in the oxidative burst, which can be used as a powerful weapon for killing M. bovis. In both acute and chronic infection, the neutrophils can function as the first line of defense for recognition and killing M. bovis, displaying enhanced activity against invading organisms.
To our knowledge, the best-known diagnostic test for tuberculosis diagnosis is the tuberculin skin test. However, exposure to environmental mycobacteria and operator errors can lead to false results. M. avium is a common environmental microbe that can colonize many different hosts, including cattle (Primm et al., 2004). Due to a degree of cross reactivity with M. bovis, it is not easy to differentiate between M. bovis and M. avium infections only by the skin test and misdiagnosis can occur. Compared with the tuberculin skin test, the IFN-γ test can also be used to differentiate between infected and vaccinated individuals. Our study demonstrated that several IFN-inducible transcriptional genes such as CD274, IFI6 and OAS1, and STAT1 in peripheral blood neutrophils were significantly differentially expressed compared with the control groups, which may have implications for vaccine and therapeutic development, particularly through the use of MPO and PTX3 as assistant genetic biomarkers with potential as diagnostic tools.
Footnotes
Acknowledgments
This work was supported by the key project of Ministry of Agriculture, China (Project No. 30972164, No. 31001048, No. 31172293, and No. 31272532), the special scientific fund for non-profit public industry (Agriculture), China (Project No.200903027), and the Beijing Science Foundation of China (Project No.6101002). The authors would also like to thank the China Agricultural University, which provided them with BSL-3 Laboratories for experiments using M. bovis culture and infection.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.