
Abstract
Butylated hydroxyanisole (BHA) is a synthetic phenolic food additive that is used to prevent oils, fats, and shortenings from oxidative deterioration and rancidity. It was tested for potential cytotoxicity and genotoxicity upon A549 lung cancer cells. MTT assay and flow cytometry analysis were used for cytotoxicity assessment, whereas genotoxicity was evaluated in vitro by DAPI staining, alkaline comet, and DNA fragmentation assays. BHA dose- and time-dependently decreased the growth of A549 cells with an IC50 of ∼0.55, 0.4, and 0.3 mM BHA at 24, 48, and 72 h, respectively. Primary and late apoptosis in the treated cells was determined using the flow cytometry analyses. In addition, single-strand DNA breakage has been observed through the comet assay technique. In addition, the morphology of DAPI-stained cells and DNA fragmentation assay using gel electrophoresis showed clear fragmentation in the chromatin and DNA rings within the nucleus of cells treated with BHA.
Introduction
A number of studies in animals showed that BHA exhibits a wide range of biological activities. Dietary administration of this compound protects animals against radiation and acute toxicity of radiation and numerous xenobiotics and inhibits carcinogenesis in numerous tissues, including the liver, colon, lung, and breast (Yu et al., 2000; Liu et al., 2007). These protective actions of BHA were due to its ability in inducing phase II detoxifying enzymes such as epoxide hydrolases, quinone reductases, glutathione S-transferases, and UDP-glucuronosyl transferases, as well as through the inhibition of cytochrome P-450 monooxygenase (Yu et al., 1997, 2000). In contrast to its beneficial effects, BHA is also found to act as a tumor initiator or a tumor promoter in some animal models (Yu et al., 1997). At high doses, BHA induced a number of disorders as well as papilloma and carcinoma formation in the forestomachs of rats, mice, and hamsters and proliferative effects in the esophagus of pigs and primates when fed constantly (Whysner and Williams, 1996; Yu et al., 2000; Kashanian and Ezzati Nazhad Dolatabadi, 2009). Even though both anticarcinogenic and carcinogenic effects of BHA are well illustrated, the exact mechanisms of BHA effects remain doubtful, but perhaps are dose-, time-, and/or tissue-dependent.
Cyto-/genotoxic effects of BHA and the mechanism on A549 lung cancer cells still remain unidentified. Because of that in the present study, we assessed the cytotoxic effects of BHA on the inhibition of A549 lung cancer cells by the MTT assay and flow cytometry analysis. In addition, for evaluation of the genotoxicity, DAPI staining, comet, and DNA fragmentation assays were used to compare the nature of the chromatin fragmentation/condensation and DNA damage of treated cells by BHA and control cells.
Materials and Methods
Materials
The A549 lung carcinoma cell line was obtained from the cell bank of Iran (Pasteur Institute). Cell culture plates and flasks were purchased from the National and IWAKI, Japan. BHA, RPMI1640 medium, and Trypsin–EDTA (0.02%–0.05%) were obtained from Sigma Aldrich Co. Low- and normal-melting-point agarose and fetal bovine serum (FBS) were bought from Gibco, Invitrogen. The Annexin V-FITC apoptosis detection kit was purchased from Oncogene Research Products. The other chemical materials were bought from Sigma Aldrich and Merck Co.
Cell culture
The A549 human lung cancer cells were maintained in a complete medium containing RPMI 1640 and 10% FBS in a humidified incubator with an atmosphere of 95% air and 5% CO2 at 37°C. A BHA stock solution (1 mM) was prepared by dissolving an appropriate amount of BHA in a filtered RPMI 1640 medium/dimethyl sulfoxide (DMSO) (99.5:0.5%).
Cytotoxicity assay
The cytotoxic effect of BHA was determined by the MTT assay. The A549 cells were plated in 96-well plates. After an overnight culture, cells were treated with a series of concentrations of BHA. For the control group, an equal volume of DMSO (0.5%) was added into the medium. After incubation for the indicated time, 50 μL MTT reagent (2 mg/mL in phosphate-buffered saline [PBS]) was added to each well and incubated for an additional 4 h in a humidified incubator. After 4-h incubation at 37°C in a humidified 5% CO2 atmosphere, the formazan crystals, formed by oxidation of the MTT dye, were dissolved in DMSO, and the absorbance at 570 nm was measured using a spectrophotometric plate reader ELx 800 (Biotek) (Asadi-Khiavi et al., 2011).
Dual staining with Annexin V-FITC and propidium iodide
After treatment, the cells were collected and resuspended in 100 μL of Annexin V-binding buffer and 8 μL Annexin V-FITC. After 15-min incubation at room temperature in the dark, the cells were centrifuged (1000 RPM, 5 min). Then, the cell pellet was resuspended in 100 mL Annexin V binding buffer, and the cells were counterstained with 8 μL propidium iodide (PI) before analysis. The cells were analyzed using the Becton Dickinson FACSCalibur System with emission filters of 515–545 nm for FITC (green) and 600 nm for PI (red). A total of 10,000 cells per sample were acquired, and the data were analyzed with CellQuest software (Becton Dickinson) (Grosse et al., 2009; Sawai and Domae, 2011).
DAPI staining
For DAPI staining, BHA-treated A549 cells were fixed with 4% formaldehyde for 10 min, and the cells were washed three times with PBS, permeabilized with 0.1% (w/v) Triton X-100 for 5 min, washed again with PBS, and stained with 200 ng/mL dichlorofluorescin diacetate (DAPI) for 20 min, followed by an observation under a fluorescence microscope (Qiao et al., 2013).
Alkaline comet assay
The comet assay is a precise and semiquantitative tool for assessment of DNA-strand breakage. Here, we have used this technique to investigate the BHA effects on the whole chromatin structure and its DNA-breaking properties. Although, there are many lines of evidence that show the antioxidant properties of BHA, here we want to apply semihigh doses of BHA to confirm its oxidant-scavenging properties. As the reactive oxygen species (ROS; outcome of oxidative stress) can interact with macromolecules and induce breakage in their structures, we also want to find the oxidative stress level in BHA-treated cells, indirectly. We performed the alkaline comet assay based on a previous work procedure (Singh et al., 1988; Eskandani et al., 2010). In brief, normal-melting-point agarose- (1.5%) covered slides were used as a surface for the low-melting-point agarose- (0.5%) embedded cells (1×10 4) and subjected to a lysis step [4-h incubation at 4°C in 2.5 M NaCl, 100 mM Na2EDTA, and 1% Triton X-100 (pH 10.5)] in a dim place without a third agarose layer (Vandghanooni and Eskandani, 2011) and then placed in an ice-cold electrophoresis chamber containing the alkaline electrophoresis solution (300 mM NaOH and 1 mM Na2EDTA of pH>13) for 30 min to allow DNA unwinding and histones to be removed, as well. Then, electrophoresis was run for 20 min at 300 mA and 30 V in an ice-cold chamber to avoid further DNA fragmentation. After that, the slides were washed with a neutralization buffer (40 mM Tris–HCl, pH 7.5) and stained with a drop of ethidium bromide. Finally, immediate microscopic analyses (Olympus IX81 fluorescence microscope equipped with an XM10 monochrome camera; wavelength 546 nm; barrier 580 nm) were done (Singh et al., 1988; Eskandani et al., 2010). The images were analyzed by CASP software. DNA single-strand breaks were expressed as the percentage of total fluorescence DNA migrated in the tail for each nucleus (% DNA in tail/% DNA in head).
DNA fragmentation assay
A549 cells were cultured and treated with BHA (1 mM) after 24 h. Cells were washed twice with PBS (pH 7.4), and DNA was extracted based on a previous work procedure. Briefly, treated cells were incubated for 5 min in the lysis buffer containing 50 mM Tris base, 10 mM EDTA, 0.5% sodium dodecyl sulfate, and 5 units RNase at 37°C (the pH of the buffer was adjusted to 7.4). Then, the total proteins were denaturized with 500 μL of chloroform/isoamyl alcohol (24:1), and the total DNA was separated by centrifugation at 12,000 rpm. The eluants containing the DNA pellets were electrophoresed (1 h at 85 V/30 mA) on a 1.2% agarose gel containing 2 μL/100 mL SYBER green. The gel was examined and photographed by an ultraviolet gel documentation system (Baskić et al., 2006; Qiao et al., 2013).
Statistical analysis
The results displayed in the figures represent the mean of at least two independent experiments; bar represents SD. The Student's t-test or one-way analysis of variance (ANOVA) was applied for parametric data. The statistical significance was defined as p<0.05.
Results
Cytotoxic effects of BHA on A549 cells
The potential cytotoxic effect of BHA on A549 cells was investigated by the MTT assay. The results showed that BHA was able to induce cytotoxicity in A549 cells in a dose- and time-dependent manner with an IC50 of ∼0.55, 0.4, and 0.3 mM BHA at 24, 48, and 72 h, respectively (Fig. 1). In addition, light-microscopic observations illustrated that the treated and untreated cells have distinct morphologic differences in the normal and dead cell numbers and appearance.
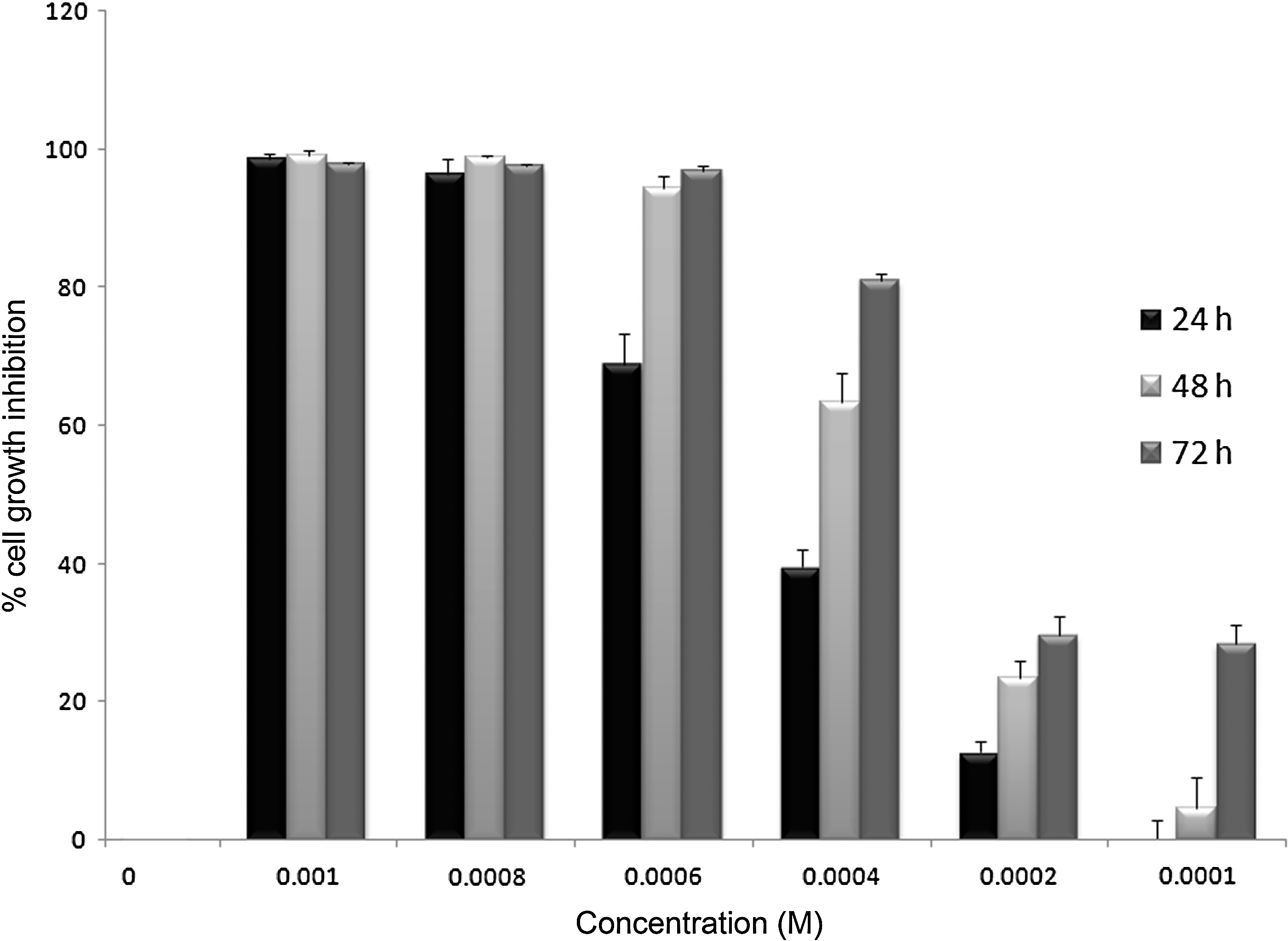
Dose- and time-dependent inhibition properties of butylated hydroxyanisole (BHA) after 24, 48, and 72 h on A549 cells. The figure shows that BHA was able to induce cytotoxicity in A549 cells in a dose- and time-dependent manner (we repeated the experiments three times, and received the same results).
Annexin V-FITC/PI staining
To distinguish apoptotic and necrotic cell death, the cells were stained with Annexin V-FITC/PI, and the time course was investigated using flow cytometry. Annexin V is a calcium-dependent phospholipid-binding protein with a high affinity for phosphatidylserine (PS), which identifies the externalization of PS on the external leaflet of the plasma membrane, an early characteristic of apoptotic cells, when conjugated to a fluorescent probe. Therefore, we could discriminate necrotic cells from apoptotic and living cells (Hao et al., 2007). More than 54% of BHA-treated cells showed the late stage of apoptosis (Annexin V-positive/PI-positive staining), and ∼33.40% of the cells were in the early stage of apoptosis (Annexin V-positive/PI-negative) after 24 h (Fig. 2). Also compared to the positive control, BHA shows a high cytotoxicity effect.
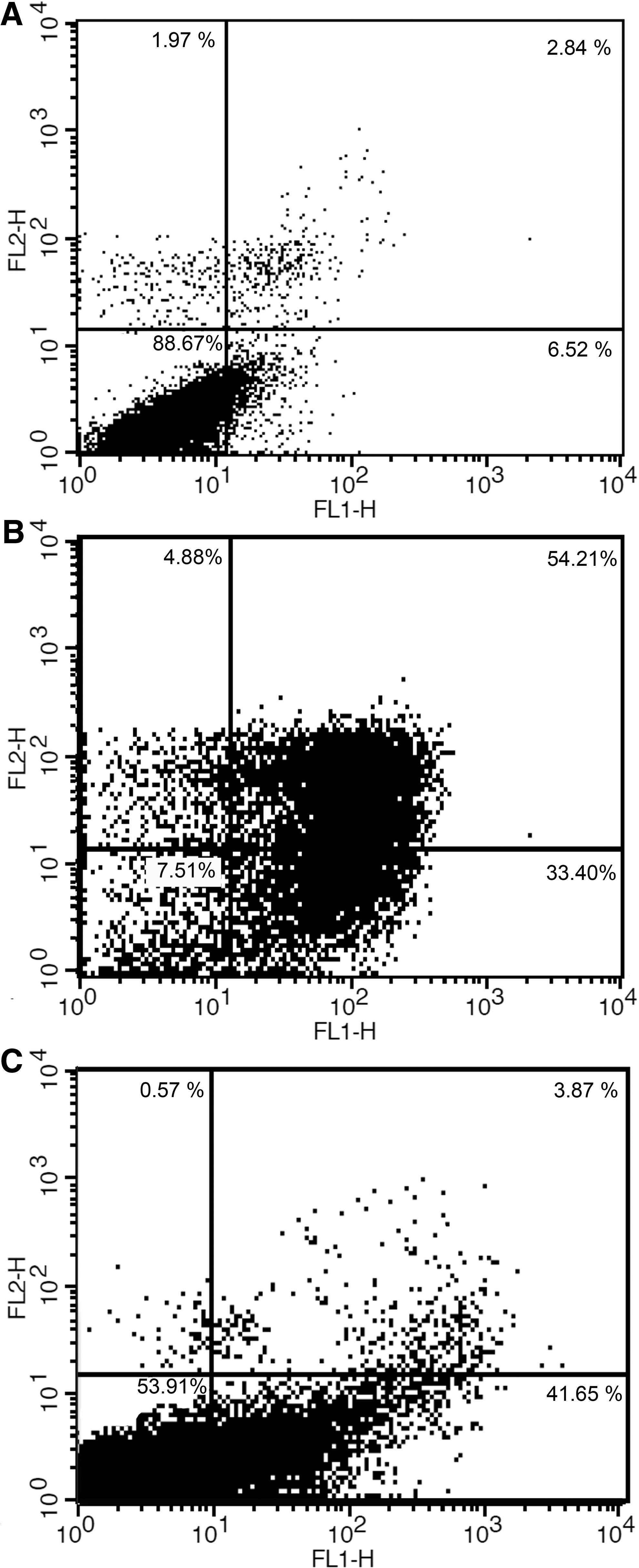
FITC-labeled Annexin V flow cytometric detection of apoptosis in A549 cells.
DAPI staining assay
Induction of apoptosis upon treatment of A549 cells with BHA was also studied by a microscopic analysis of DAPI-stained cells. Representative microscopic images of the DAPI-stained cells after 24-h exposure to 1 mM of BHA and 5% DMSO (positive control) are shown in Figure 3. As it is clear, the apoptotic cells are mainly observed in both positive control and BHA-treated cells. BHA treatment caused a statistically significant nuclear shrinkage, fragmentation in the chromatin and DNA rings within the nucleus of treated cells, but their morphology did not change in untreated normal cells (Baskić et al., 2006; Bommareddy et al., 2012).
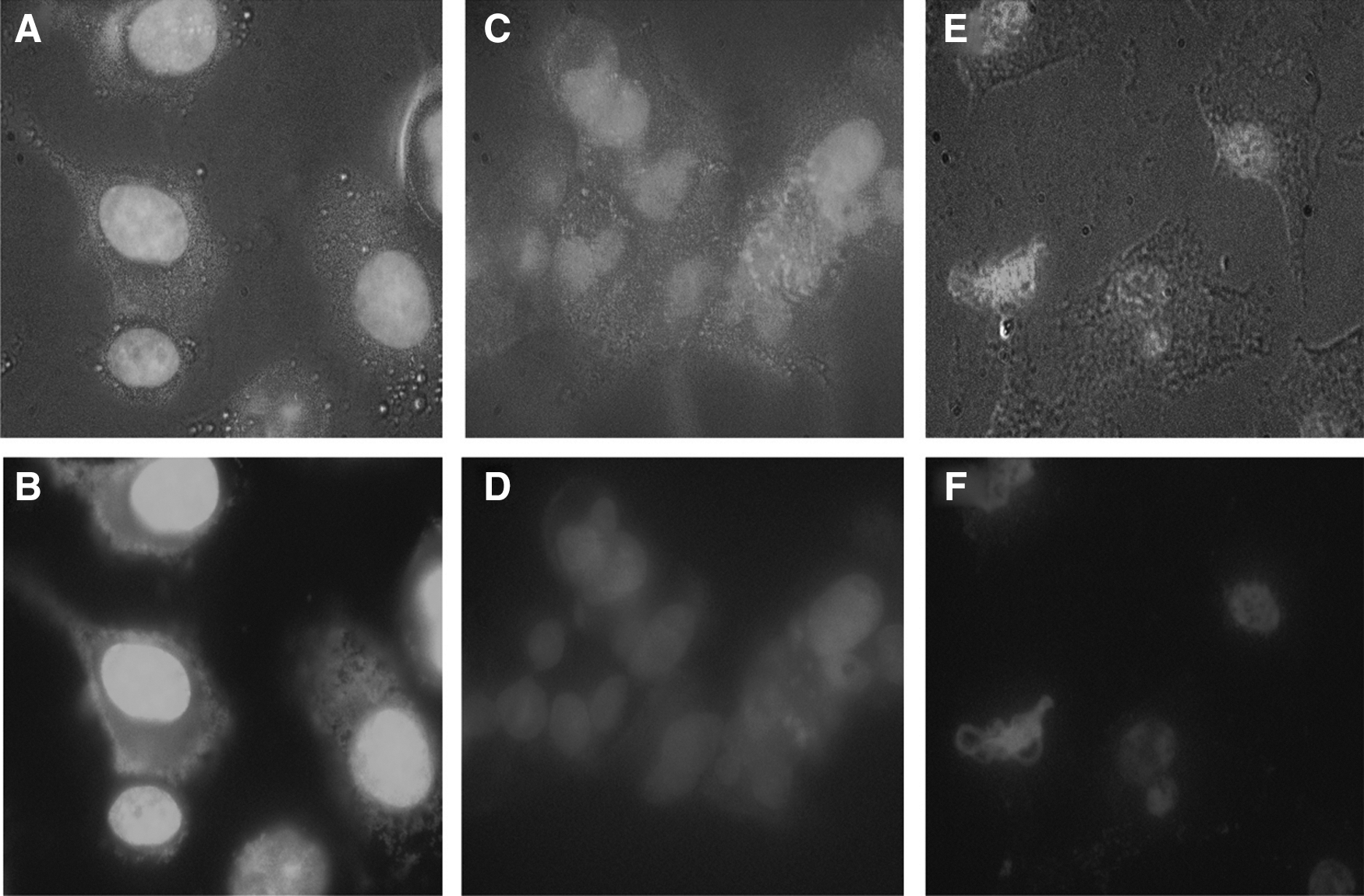
Light and fluorescent microscopy images of A549 cells stained with DAPI;
Alkaline comet assay and DNA damages
The migrated nucleus photos were further analyzed by software and statistical analyses as% head DNA/tail DNA (Fig. 4). One-way ANOVA showed significant and relatively more DNA damages in the BHA-treated cells compared to the negative control cells [Fig. 5 (p<0.001)] (Singh et al., 1988; Eskandani et al., 2010).
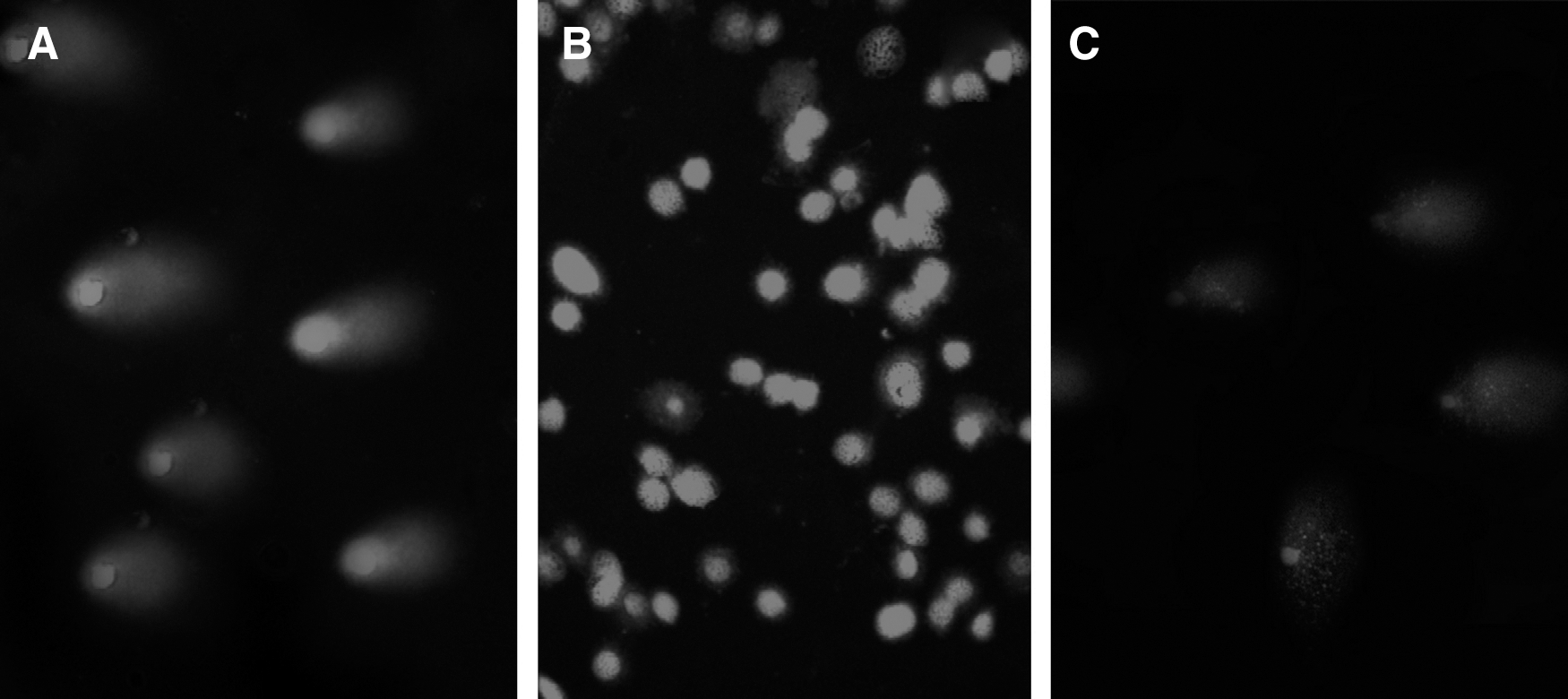
Photographic illustrations of the comet assay after 24-h incubation;
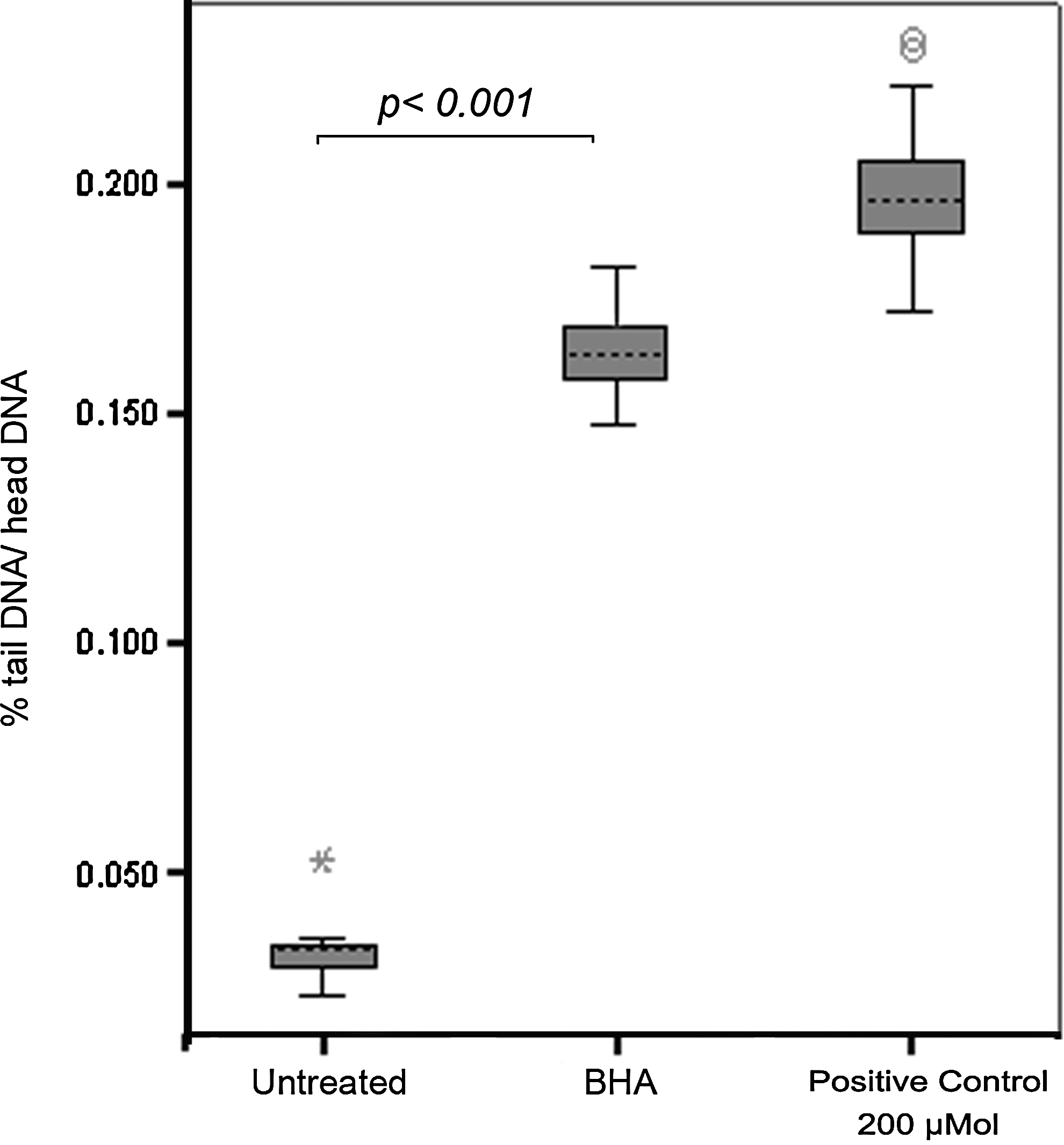
Box-plot graph of (DNA tail/DNA head %); the DNA cleavage level in BHA-treated cells is not as high as treated cells with 200 mM H2O2 but is significantly (p<0.001) higher than untreated cells.
DNA fragmentation assay
The chromatin was degraded into 200-bp fragments generally due to apoptosis in many cell types, and it is among the most reliable methods for detection of apoptotic cells (Qiao et al., 2013). Therefore, we had investigated the DNA fragment patterns and DNA ladder formation with an ordered pattern of multiple bands in agarose gels. Gel electrophoresis indicated the formation of the DNA ladder, which was induced by BHA in the concentration of 1 mM (Fig. 6). Here, clear apoptotic DNA ladders of 200-bp DNA ladder were used as the control (Nath et al., 2013).

DNA ladder formation through gel electrophoresis; M, DNA marker; 1, control; 2, DNA fragmentation due to treatment of the cells with 1 mM BHA.
Discussion
Among various food additives, BHA has been extensively used for preventing oxidation or rancidity of food products. Because of its widespread usage in food products, it is ingested by virtually every individual who consumes packaged foodstuffs. The maximum acceptable concentration of BHA in food products has been set at 0.02% of the fat and oil content of foods by the World Health Organization (Leslie et al., 1978; Williams et al., 1990).
A growing number of studies showed that oral administration of BHA causes cytotoxicity and enhances the development of preneoplastic and neoplastic lesions in the mouse forestomach and urinary bladder and induces forestomach squamous cell carcinomas in rodents at high dosage (Whysner and Williams, 1996; Yu et al., 1997). Also, the BHA interaction with mitochondria and triggering of mitochondrial permeability transition can result in the release of cytochrome c and induce apoptosis as well (Yu et al., 2000). Although the cytotoxicity of BHA has been well documented, the mechanistic aspect by which BHA induces cell death is not clear. Therefore, in the present study, we focused on evaluating the effects of BHA on the growth and death of A549 cells.
In the current study, we examined the cytotoxic effects of BHA on A549 cells. It was shown that BHA was able to induce cytotoxicity in A549 cells in a dose- and time-dependent manner. Using FITC-labeled Annexin V flow cytometry, we observed the happening of early/late apoptosis and even necrosis within A549 cells treated with BHA (Fig. 2). The results arisen from flow cytometry revealed that BHA led cell death mainly through an apoptotic pathway not a necrotic pathway. To pursue the mechanism behind such cytotoxicity using the DAPI stain, comet, and DNA fragmentation assay, significant fragmentation of DNA was observed in A549 cells treated with BHA (Figs. 3, 4, and 6). The direct interaction of BHA with DNA in vitro and generation of reactive intermediates in various cell lines have been reported (Yu et al., 1997, 2000; Kashanian and Ezzati Nazhad Dolatabadi, 2009). In this study, the alkaline comet assay results confirmed that the direct interaction of BHA with DNA and increasing free radicals such as ROS can be two probable reasons for apoptosis induction. Also, the formation of DNA ladder through the DNA fragmentation assay validated the DNA breakage and fragmentation. Therefore, we speculate that BHA may have prooxidant properties at high dosage, and cell death induced by BHA may be associated with oxidative stresses derived from ROS. For that reason, we concluded that there is a possible correlation between cytotoxicity and DNA damage in the treated cells with the BHA.
Conclusively, the present study results showed that BHA as a food additive inhibited the growth of A549 cells in a dose- and time-dependent manner and stimulated apoptosis and inherent material toxicity. It was shown that BHA inhibits the growth rate of cells by inducing apoptosis via chromatin and DNA fragmentation. We showed the occurrence of early/late stages of apoptosis within cells treated with BHA using FITC-labeled Annexin V flow cytometry. In addition, using the comet and DNA fragmentation assays, significant fragmentation of DNA was illustrated in treated cells. Finally, all experiment data supported the fact that it is worthwhile to make thorough analyses on the extensive usage of BHA in food industries, and it should be noted that the concentrations of BHA used in this study were lower than the concentration expected from human exposure to antioxidants in food products.
Footnotes
Acknowledgment
The authors are grateful for the financial support from the Research Center for Pharmaceutical Nanotechnology, Tabriz University of Medical Sciences.
Disclosure Statement
No competing financial interests exist.