
Abstract
Serum paraoxonase (PON1) is an esterase that is involved in the detoxification of organophosphate insecticides. Emerging lines of evidence have shown that functional polymorphisms in the PON1 gene might play a critical role in increasing susceptibility to organophosphate toxicity, but individually published studies showed inconclusive results. This meta-analysis aimed to derive a more precise estimation of the associations between the PON1 polymorphisms and organophosphate toxicity risk. Nine case–control studies were assessed with a total 1,042 patients with organophosphate toxicity and 1014 healthy controls. The meta-analysis results showed that the PON1 192Q and 55L polymorphisms may increase the risk of organophosphate toxicity. Further subgroup analyses by ethnicity showed significant associations of the PON1 192Q and 55L polymorphisms with increased risk of organophosphate toxicity among the Caucasian populations. However, similar associations were not observed among the Asian populations. In conclusion, the current meta-analysis indicates that the PON1 192Q and 55LM polymorphisms may increase the risk of organophosphate toxicity, especially among the Caucasian populations.
Introduction
Paraoxonase 1 (PON1) is a high-density lipoprotein-associated serum enzyme that involved in the detoxification of organophosphate insecticides such as parathion, diazinon, and chlorpyrifos (Mackness and Mackness, 2011). PON1 catalyzes the hydrolysis of toxic organophosphates, and is widely distributed among tissues such as the liver, kidney, intestine, and serum (Li et al., 2003). The PON1 gene is located on chromosome 7q21-q22 and consists of ∼26.2 kbps (Ichikawa et al., 2009). The human PON1 gene displays two polymorphisms in the coding region (Q192R and L55M) and several polymorphisms in the promoter and the 3′-UTR regions (Costa et al., 2012). Genetic alterations in the PON1 may alter organophosphate metabolism and particular detoxification pathways, which may also contribute to the risk of organophosphate toxicity (Povey, 2010). Therefore, it was hypothesized that single-nucleotide polymorphisms (SNPs) in the PON1 gene could be functional and were associated with the organophosphate toxicity risk (Furlong et al., 2010).
Recently, the molecular basis of the PON1 genetic polymorphisms and their role in organophosphate toxicity have been extensively investigated (Costa et al., 2012). Several studies have indicated that two common polymorphisms (Q192R and L55M) in the PON1 gene might play a critical role in increasing the susceptibility to organophosphate toxicity (Catano et al., 2006; Povey et al., 2007; Wang et al., 2007; Androutsopoulos et al., 2011). However, some other studies exist that suggest that the PON1 genetic polymorphisms are not associated with the risk of organophosphate toxicity (Brophy et al., 2001; Hernandez et al., 2005; Searles Nielsen et al., 2005). In view of the conflicting results from previous studies, we performed a meta-analysis of published data evaluating the associations of the PON1 Q192R (rs662 A>G) and L55M (rs854560 T>A) polymorphisms with organophosphate toxicity risk.
Materials and Methods
Literature search strategy
Relevant articles published before October 1, 2012, were identified through a search on Pubmed, Embase, Web of Science, and Chinese BioMedical databases using the following terms: (“genetic polymorphism” or “polymorphism” or “single nucleotide polymorphism” or “SNP” or “gene mutation” or “genetic variants”) and (“pesticides” or “insecticides” or “insect repellents” or “organophosphorus pesticide”) and (“paraoxonase 1” or “aryldialkyl phosphatase” or “PON1” or “OPH enzyme” or “organophosphorus hydrolase”). The references from the eligible articles or textbooks were also reviewed manually to check for other potential studies. Disagreements were resolved through discussions between the authors.
Inclusion and exclusion criteria
Studies included in our meta-analysis have to meet the following criteria: (1) case–control studies or cohort studies focused on the associations of the PON1 Q192R and L55M polymorphisms with susceptibility to organophosphate toxicity; (2) all patients with organophosphate toxicity should be confirmed by a history of organophosphate pesticide expose and essential clinical criteria; (3) published data about the frequencies of alleles or genotypes must be sufficient. Studies were excluded when they were (1) not a case–control study or a cohort study; (2) duplicates of previous publications; (3) based on incomplete data; (4) meta-analysis, letters, reviews, or editorial articles. If more than one study by the same author using the same case series was published, either the study with the largest sample size or the most recently published study was included.
Data extraction
Using a standardized form, data from the included studies were extracted independently by two authors. For each study, the following characteristics and numbers were collected: the first author, year of publication, country, language, study design, numbers of subjects, ethnicity of subjects, source of cases and controls, type of insecticides, detecting sample, genotype method, allele and genotype frequencies, and evidence of the Hardy–Weinberg equilibrium (HWE) in controls. In cases of conflicting evaluations, disagreements were resolved through discussion between the authors.
Quality assessment
Two authors independently assessed the quality of included studies according to the modified STROBE quality score systems (da Costa et al., 2011). Forty assessment items related to quality appraisal were used in this meta-analysis with scores ranging from 0 to 40. Scores of 0–20, 20–30, and 30–40 were defined as low, moderate, and high quality, respectively. Disagreements were resolved through discussion between the authors.
Statistical analysis
Crude odds ratio (OR) with 95% confidence intervals (CI) was calculated under five genetic models: the allele model, the dominant model, the recessive model, the homozygous model, and the heterozygous model. The statistical significance of the pooled OR was examined by the Z test. Between-study variations and heterogeneities were estimated using Cochran's Q-statistic with a ph< 0.05 as statistically significant heterogeneity (Jackson et al., 2012). We also quantified the effect of heterogeneity by using the I2 test (ranges from 0 to 100%), which represents the proportion of interstudy variability that can be contributed to heterogeneity rather than to chance (Zintzaras and Ioannidis, 2005). When a significant Q-test with ph<0.05 or I2>50% indicated that heterogeneity among studies existed, the random-effects model (DerSimonian Laird method) was conducted for the meta-analysis. Otherwise, the fixed-effects model (Mantel-Haenszel method) was used. To explore the sources of heterogeneity, we also performed a subgroup analysis by the ethnicity and genotype method. We tested whether the genotype frequencies of controls were in the HWE using the χ2 test. Sensitivity analysis was performed by omitting each study in turn to assess the stability of results. Begger's funnel plots were used to detect publication bias. Egger's linear regression test was also used to evaluate the publication bias (Peters et al., 2006). All the p values were two-sided. All analyses were calculated using STATA Version 12.0 software (Stata Corp., College Station, TX).
Results
Study characteristics
In accordance with the inclusion criteria, 9 case–control studies (Padungtod et al., 1999; Hernandez et al., 2003; Mackness et al., 2003; Hernandez et al., 2005; Searles Nielsen et al., 2005; Povey et al., 2007; Wang et al., 2007; da Silva et al., 2008; Singh et al., 2011) were included in this meta-analysis, and 102 were excluded. The flow chart of the study selection process is shown in Figure 1. A total of 2056 subjects were involved in this meta-analysis, including 1042 organophosphate toxicity patients and 1014 healthy controls. The publication year of involved studies ranged from 1999 to 2011. Six of the eight studies were conducted in the Caucasian populations, and the other three studies in the Asian populations. All included studies used blood samples for genotyping. A classical polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) method was performed in six studies. The other three studies used the TaqMan assay. The HWE test was conducted on the genotype distribution of the controls in all 12 studies. None of the studies deviated from the HWE (all p>0.05). All quality scores of included studies were higher than 20 (moderate–high quality). The characteristics and methodological quality of the included studies are summarized in Table 1. A summary of the meta-analysis findings of the associations between the PON1 polymorphisms and organophosphate toxicity risk is provided in Table 2.
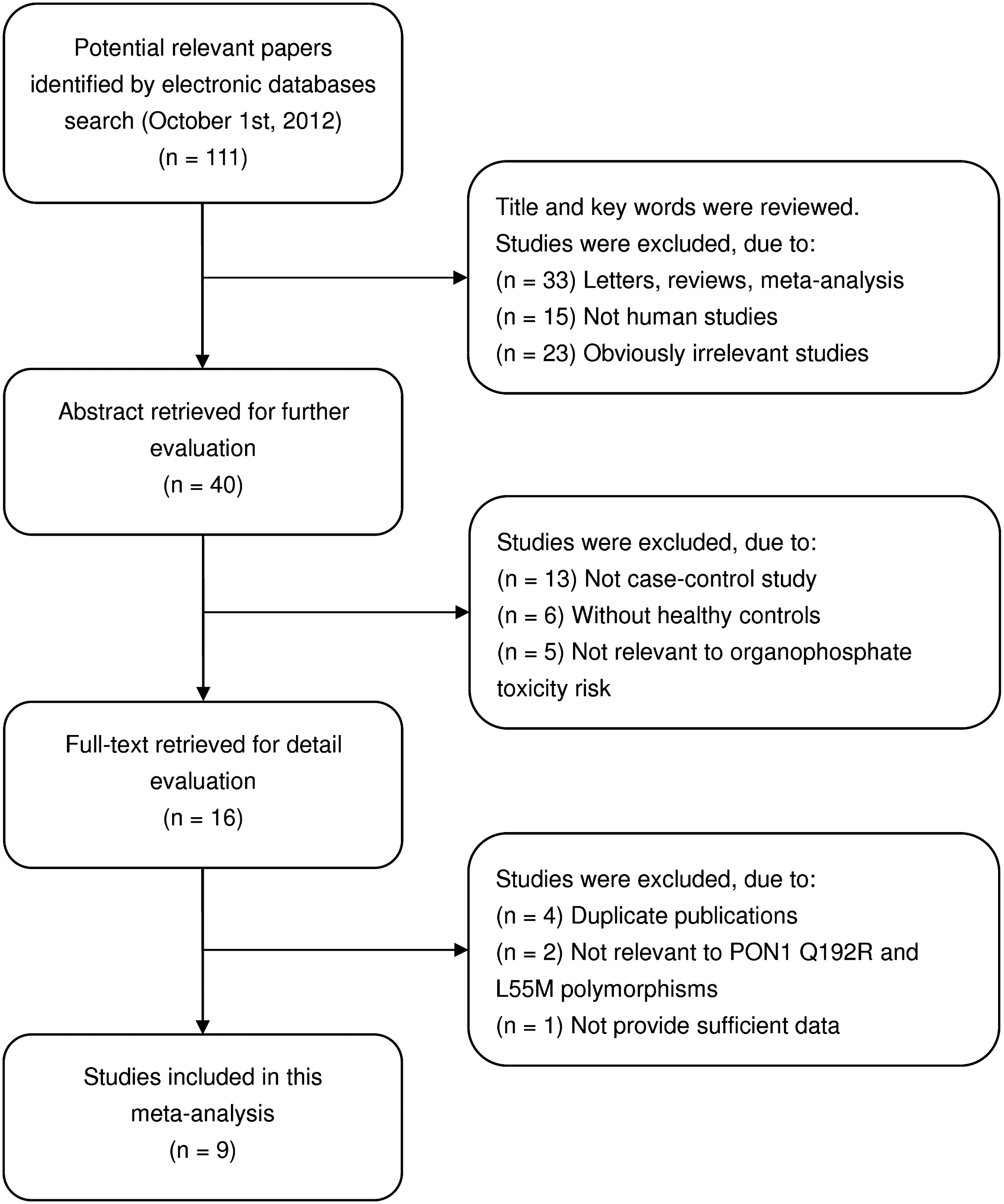
Flowchart shows study selection procedure.
PCR-RFLP, polymerase chain reaction–restriction fragment length polymorphism; PB, population-based; PON1, paraoxonase 1; SNP, single-nucleotide polymorphism.
Estimates for random-effects model.
OR, odds ratios; 95%CI, 95% confidence interval; ph , p value of heterogeneity test.
Quantitative data synthesis
The association between the PON1 Q192R polymorphism and organophosphate toxicity risk is investigated in eight studies. No heterogeneity was observed under all genetic models (all ph>0.05), so the fixed-effects model was used. The meta-analysis results indicated that the PON1 192Q polymorphism may increase the risk of organophosphate toxicity (Q allele vs. R allele: OR=1.18, 95%CI=1.02–1.36, p=0.028; QQ+QR vs. RR: OR=1.74, 95%CI=1.31–2.32, p<0.001; QQ vs. RR: OR=1.73, 95%CI: 1.27–2.34, p<0.001). Further subgroup analyses by ethnicity showed significant associations between the PON1 192Q polymorphism and increased risk of organophosphate toxicity among the Caucasian populations (Q allele vs. R allele: OR=1.33, 95%CI=1.03–1.71, p=0.029; QQ+QR vs. RR: OR=1.96, 95%CI=1.37–2.82, p<0.001; QQ vs. RR: OR=1.71, 95%CI: 1.18–2.50, p=0.005), but similar associations were not observed among the Asian populations (Fig. 2).
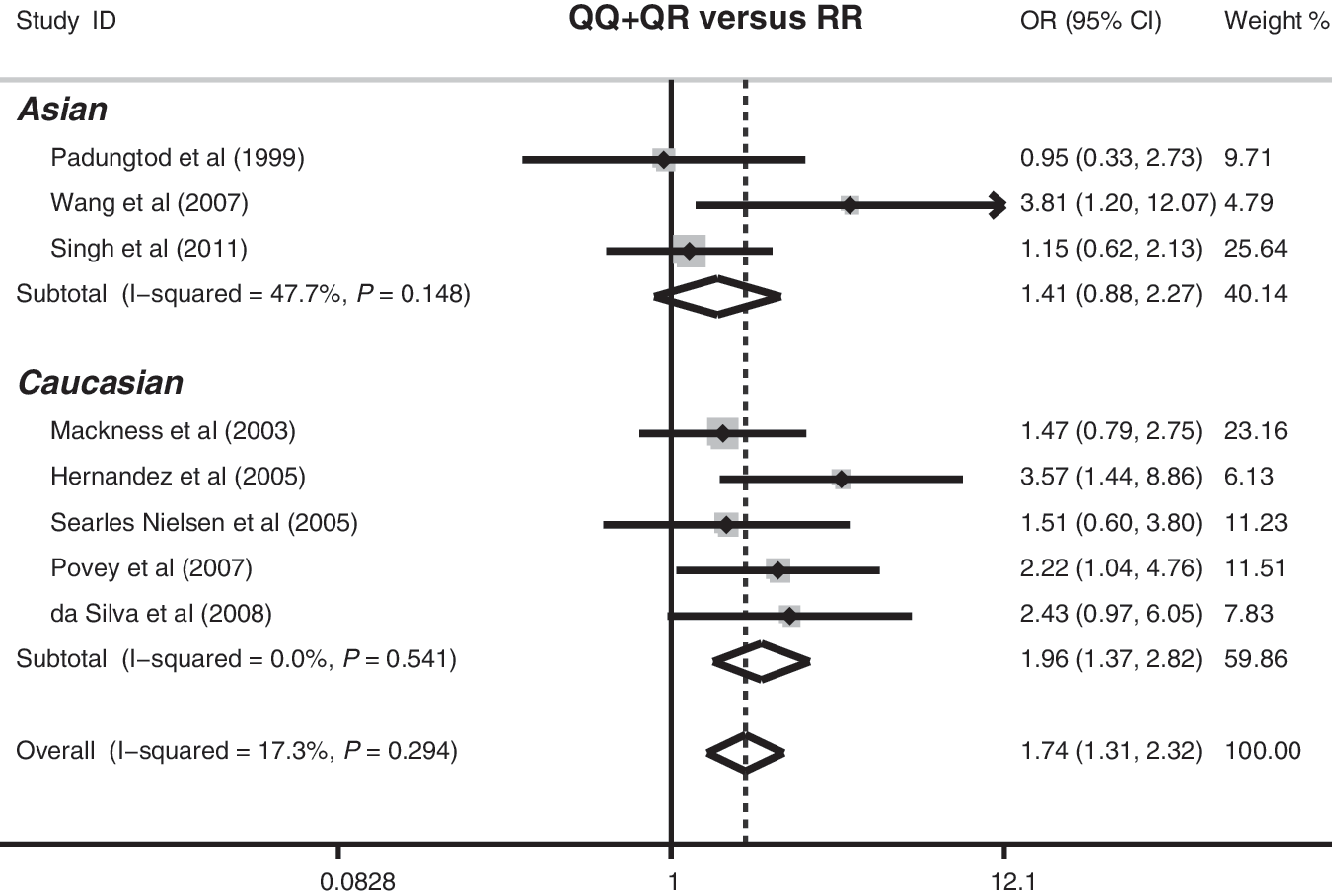
Forest plot of the association between the PON1 Q192R polymorphism and organophosphate toxicity risk under the dominant model (QQ+QR versus RR).
Four studies referred to the association between the PON1 L55M polymorphism and organophosphate toxicity risk. The heterogeneity was significantly observed under the recessive model (p=0.049), which might result from the difference in the ethnicity, country, and genotype method, so the random-effects model was used. Meta-analysis of these studies showed that there were significant associations between the PON1 55L polymorphism and increased risk of organophosphate toxicity (L allele vs. M allele: OR=1.35, 95%CI=1.12–1.61, p=0.001; LL+LM vs. MM: OR=1.58, 95%CI=1.09–2.30, p=0.016; LL vs. MM: OR=1.82, 95%CI: 1.23–2.71, p=0.003). Results from the subgroup analysis based on ethnicity indicated that the PON1 55L polymorphism may increase the risk of organophosphate toxicity among the Caucasian populations (L allele vs. M allele: OR=1.53, 95%CI=1.23–1.89, p<0.001; LL+LM vs. MM: OR=1.94, 95%CI=1.23–3.06, p=0.004; LL vs. MM: OR=2.48, 95%CI: 1.53–4.04, p<0.001), but not in the Asian populations (Fig. 3).
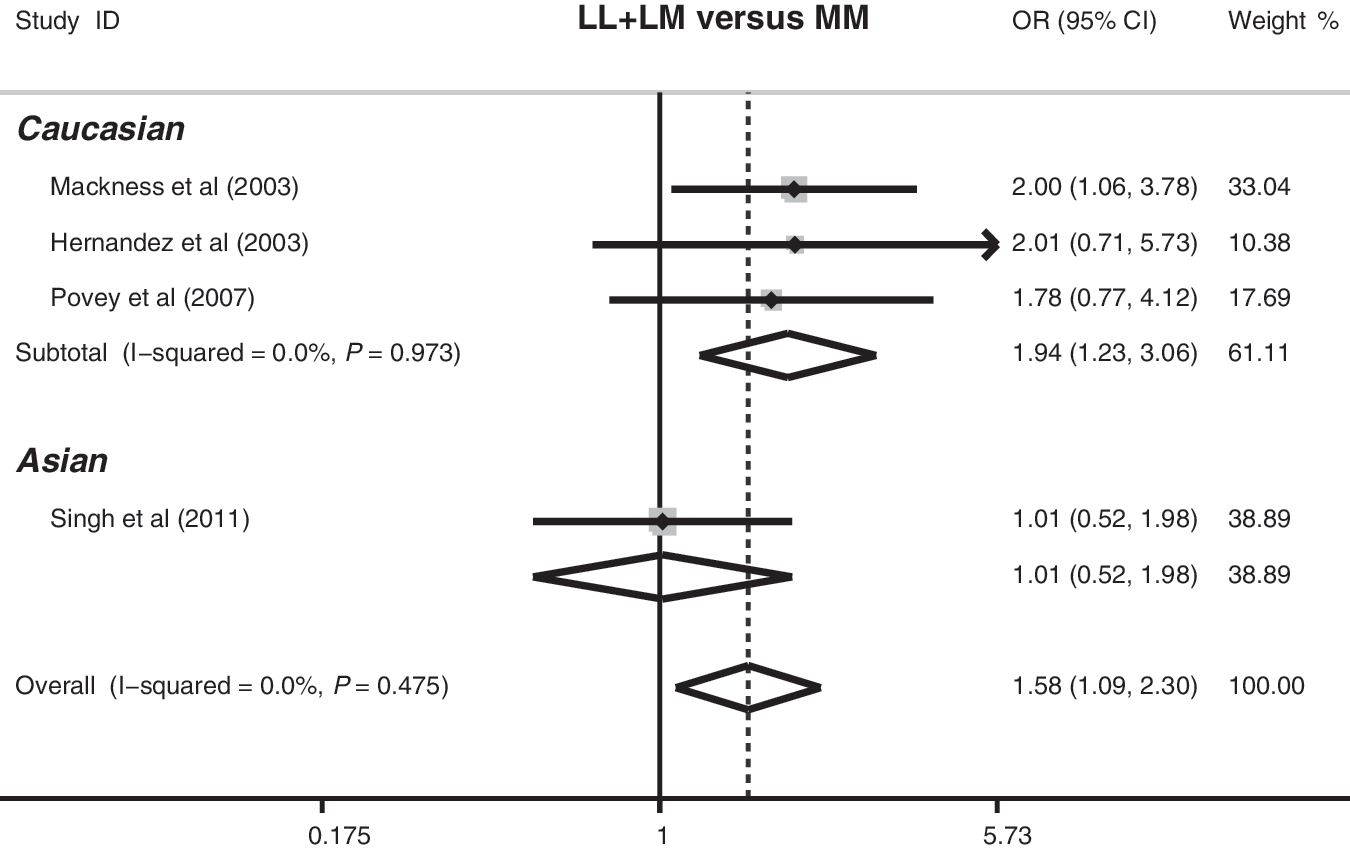
Forest plot of the association between the PON1 L55M polymorphism and organophosphate toxicity risk under the dominant model (LL+LM versus MM).
Sensitivity analysis
Sensitivity analysis was performed to assess the influence of each individual study on the pooled ORs through omitting of individual studies. The analysis results suggested that no individual studies significantly affected the pooled ORs under the dominant model of the PON1 Q192R and L55M polymorphisms (Fig. 4), indicating a statistically robust result.

Sensitivity analysis of the associations of the PON1 Q192R and L55M polymorphisms with organophosphate toxicity risk under the dominant model. Results were computed by omitting each study in turn. Meta-analysis random-effects estimates (exponential form) were used. The two ends of the dotted lines represent the 95% CI. CI, confidence interval.
Publication bias
Publication biases within the available research results might not be representative of all research results. Begger's funnel plot and Egger's linear regression test were performed to assess the publication biases of the included studies. The shapes of the funnel plots did not reveal any evidence of obvious asymmetry under the dominant model of the PON1 Q192R and L55M polymorphisms (Fig. 5). Egger's test also showed that there was no strong evidence of publication bias (Q192R: t=1.38, p=0.216; L55M: t=0.46, p=0.692).

Begger's funnel plot of publication bias for the PON1 Q192R and L55M polymorphisms with organophosphate toxicity risk under the dominant model. Each point represents a separate study for the indicated association. Log[OR], natural logarithm of OR. Horizontal line, mean magnitude of the effect. Funnel plot with pseudo-95% confidence limits was used. OR, odds ratio.
Discussion
PON1 is one of the three members of human serum paraoxonase family, which is one of the key drug-metabolizing enzymes involved in organophosphate compound metabolism by hydrolyzing organophosphate esters (Furlong et al., 2010). SNPs within PON1 confer differences to PON1 activity and expression levels that may render a subset of the population susceptible to organophosphate toxicity (Huen et al., 2009). Mutations in the PON1 gene are significant because they may alter organophosphate metabolism and particular detoxification pathways, which may not allow adequate time for detoxification before enzyme inactivation and the activity of other metabolizing enzymes (Povey, 2010). Many previous genetic studies have suggested that the PON1 Q192R and L55M polymorphisms contribute the risk-enhancing effects for the development of organophosphate toxicity (Catano et al., 2006; Povey et al., 2007; Wang et al., 2007; Androutsopoulos et al., 2011), while other studies found no convincing evidence of these polymorphisms in increasing susceptibility to organophosphate toxicity (Brophy et al., 2001; Hernandez et al., 2003; Searles Nielsen et al., 2005). These disparate results may be due to a number of methodological limitations, including small sample size, use of poor or inaccurate exposure estimates, referent groups that may have differed from the exposure group on characteristics other than exposure (e.g., sheep dippers versus ceramic workers), and inadequate control of potential confounding variables, such as previous pesticide poisoning (Starks et al., 2012). Therefore, we conducted this meta-analysis to provide a profound understanding of correlations between the two common polymorphisms (Q192R and L55M) in the PON1 gene and the risk of organophosphate toxicity.
In this meta-analysis, nine independent case–control studies were included with a total of 1,042 patients with organophosphate toxicity and 1,014 healthy controls. When all the eligible studies were pooled into the meta-analysis, the results showed that the PON1 192Q and 55L polymorphisms were associated with an increased risk of organophosphate toxicity, especially among patients Caucasian populations, while similar associations were not observed among patients Asian populations. There are two potential explanations for this finding. First, the lack of association between the PON1 genetic polymorphisms and organophosphate toxicity risk among patients Asian populations may be due to a lack of a sufficient number of eligible studies. Another potential explanation may be the interactions between the genetic and environmental factors. More research is needed to determine the associations between the PON1 genetic polymorphisms and organophosphate toxicity risk among the Asian populations. Although the exact function of the PON1 genetic polymorphisms in the development of organophosphate toxicity among different populations is not yet clear, a possible reason could be that inherited mutations in PON1 might be associated with changes in the organophosphate metabolites and thereby could possibly explain the interindividual differences in susceptibility to organophosphate toxicity (Costa et al., 2012). In addition, we also conducted subgroup analysis based on the genotype method for the Q192R and L55M polymorphisms, and it came out that there existed significant correlations of the PON1 192Q and 55L polymorphisms with the risk of organophosphate toxicity in the PCR-RFLP subgroup, but not in the TaqMan subgroup. These findings are consistent with the previous hypothesis that variability in the PON1 gene may increase the risks of organophosphate toxicity, suggesting that the PON1 192Q and 55L polymorphisms may be useful as biomarkers in predicting an individual's susceptibility to organophosphate toxicity (Furlong et al., 2010).
Some limitations of this meta-analysis should be acknowledged. First, there were only nine articles included in the present meta-analysis, so the sample size was relatively small and may not provide sufficient statistical power. Therefore, more studies with larger sample size are needed to accurately provide a more representative statistical analysis. Second, as a type of a retrospective study, a meta-analysis may encounter recall or selection bias, possibly influencing the reliability of our study results. Third, our lack of access to the original data from the studies limited further evaluation of potential interactions between other factors and organophosphate toxicity risk, such as gene–environment and gene–gene interactions. In spite of these limitations, however, this is the first meta-analysis of the relationship between the PON1 genetic polymorphisms and susceptibility to organophosphate toxicity.
In conclusion, this meta-analysis provides strong evidence that the PON1 192Q and 55L polymorphisms may be associated with an increased risk of organophosphate toxicity, especially among the Caucasian populations. These relationships have the potential to provide functional profiling of the PON1 gene involved in organophosphate metabolism and to help us understand the biological processes associated with the development of organophosphate toxicity. However, detailed studies are still needed to confirm our findings. Further studies investigating the effect of the gene–environment interactions on organophosphate toxicity are also essential.
Footnotes
Disclosure Statement
No competing financial interests exist.