
Abstract
The Editor-in-Chief of DNA and Cell Biology officially retracts the article entitled, “The E2F1 Oncogene Transcriptionally Regulates NELL2 in Cancer Cells” by Dong Hee Kim, Yun-Gil Roh, Hyun Hee Lee, Sang-Yeop Lee, Seung Il Kim, Byung Ju Lee, and Sun-Hee Leem (DNA Cell Biol. 2013;32(9):517–523; doi: 10.1089/dna.2013.1974) after discovering and investigating a report of duplicated images (in Figure 5) related to the paper on the PubPeer platform.1
The publisher of the journal contacted all authors listed on the paper to inquire about the problems identified on PubPeer. The lead author, Dr. Dong Hee Kim, responded by stating, “There seems to be a problem in the processing of working on the figures,” and supplied a new figure to replace the duplicated image. After reviewing the newly supplied figure and the author's answer, the Editor of the journal found the author's response to be unacceptable and called for a full retraction of the published paper.
DNA and Cell Biology
is committed to upholding the rigors of scientific publishing and the veracity of the literature.
1. Actinopolyspora biskrensis. PubPeer.
Introduction
According to a recent study, NELL2 is repressed by the antitumor agent Genistein in pancreatic cancer Panc1 cells (Bai et al., 2004) and is overexpressed in Burkitt's lymphoma cells and neuronal tumors (Kuroda et al., 1999; Maeda et al., 2001). However, the molecular mechanism of NELL2 overexpression in human cancer cells remains unclear.
The E2F family represents a class of transcription factors, which regulate a broad range of genes involved in major cellular processes, such as DNA replication, apoptosis, differentiation, and the cell cycle (Iaquinta and Lees, 2007; Wu et al., 2009). Eight E2F genes have been identified in mammals that encode proteins classified as transcriptional activators (E2F1–3) or repressors (E2F4–8) (DeGregori and Johnson, 2006). E2F1, the founding member of the E2F family, has been shown to possess oncogenic properties and considerable evidence shows that regulation of the E2F activity plays a key role in tumorigenesis (Tsantoulis et al., 2005; Chen et al., 2009).
In this study, we found potential E2F1 binding consensus sequences in the NELL2 promoter region. Using promoter function assays and chromatin immunoprecipitation (ChIP) assays, we found that E2F1 could directly transactivate NELL2 expression by binding to its promoter. Therefore, our results suggest that frequent and persistent expression of E2F1 is required for cell growth control through transcriptional activation of NELL2 expression in cancer cells.
Materials and Methods
Cell culture, promoter assay, and transfection
Human bladder cancer cell lines (UC5, 5637, EJ, and T24 cells) and breast cancer cell lines (MCF7, MDA-MB231, and MDA-MB435 cells) were cultured in the RPMI medium (Hyclone) and H-DMEM (Hyclone) supplemented with 10% fetal bovine serum (Hyclone), penicillin (100 unit/mL), and streptomycin (100 μg/mL). All cells were incubated at 37°C under 5% CO2 in a humidified incubator. To determine whether E2F1 regulates NELL2 transcription, 5637 and MDA-MB231 cells were transiently cotransfected with NELL2 promoter-luciferase reporter constructs (kindly provided by Dr. Lee, University of Ulsan, Ulsan, South Korea) (Choi et al., 2010) and an E2F1 expression vector (pcDNA6; Invitrogen) containing the human E2F1 gene (NCBI GenBank database accession No. BC050369) using jetPrime reagent (Polyplus-Transfection, Inc.). Lysates of the transfected cells were generated with the Cell Culture Lysis Reagent (Promega) and the chemiluminescence signal was measured in a Wallac Victor 1420 Multilabel Counter (EG&G Wallac). Transfection efficiency of each assay was normalized by cotransfecting a Renilla luciferase control construct.
Small interfering RNA transfection
The 5637 and MDA-MB231 cells were transfected with 100 nM E2F1 small interfering RNA (Dhamarcon) using jetPrime. Cells were harvested 24 h after transfection.
Real-time polymerase chain reaction
To determine the effect of E2F1 on endogenous NELL2 expression, RNA (2 μg) was isolated from the breast and bladder cancer cell lines using the Tri-reagent (Ambion). The isolated RNA samples were reverse transcribed and amplified using quantitative real-time polymerase chain reaction (PCR) with the following primer sets: E2F1 sense primer, 5′-TGC CCT GAG GAG ACC GTA G-3′; antisense primer, 3′-GGT GAC ACT ATG GTG GCA GAG-5′; NELL2 sense primer, 5′-GTC ATG CCC CAG GGA TTT-3′; antisense primer, 3′-CGA GAC AGC TTG GCT GAT GT-5′. For quantitative real-time PCR, 50 ng of cDNA were added to SYBR Green dye (Qiagen) and run in a CFX96™ Optics Module (Bio-Rad, Inc.).
Western blot analysis
Proteins from the 5637 and MDA-MB231 cells were homogenized in the RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 1% NP40, 0.5% sodium deoxycholate, 0.1% SDS) containing a protease inhibitor (Roche). The extracted protein (15 μg) was separated by SDS-PAGE and transferred to a membrane by electrophoretic transfer. The membrane was incubated with a rabbit anti-E2F1 (Cell Signaling Technology, Inc.) and rabbit anti-NELL2 antibody (Santa Cruz Biotechnology). Immunoreactivity was detected using the ECL detection system (GE Healthcare Bio-Sciences Corp.). Films were exposed for multiple time points to ensure that images were not saturated.
ChIP assay
Breast and bladder cancer cells transfected with expression vectors for E2F1 were lysed and fixed by addition of 1% formaldehyde to the medium for 10 min. Free formaldehyde was quenched by addition of 125 mM glycine for 5 min at room temperature. The cells were washed twice with cold PBS, and then cold PBS containing Protease Inhibitor Cocktail II (Roche) was added, followed by harvesting of the cells by scraping and centrifugation. Nuclei were extracted and resuspended with a nuclear lysis buffer (50 mM Tris, pH 8.1, 10 mM EDTA, 1% SDS, and protease inhibitors). Chromatin was sheared by sonication and diluted fivefold in the ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris, pH 8.1, 167 mM NaCl, and protease inhibitors). The reactions were incubated with 5 μg of antibodies against E2F1 (Cell Signaling) at 4°C overnight. Immune complexes were collected by reacting with 60 μL of salmon sperm DNA/protein A-agarose for 1 h at 4°C and then washed consecutively for 5 min each with buffers (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris, pH 8.1) containing different concentration of salts (150–500 mM) and 0.25 M LiCl. DNA from the protein-DNA crosslinks was extracted by incubating the reactions with a solution of 1% SDS, 0.1 M NaHCO3, 10 μg RNase, and 0.3 M NaCl at 65°C for 4 h and was further purified with phenol/chloroform. PCR amplification was performed using 30 cycles of 94°C for 30 s, 53°C for 30 s, and 72°C for 30 s, preceded by 94°C for 5 min, and followed by 72°C for 10 min. Primer sets used for the PCR amplification were a primer set for the E2F1 site at position −73 (sense primer, 5′-GCT CGG TCT TAG GTG TCT G-3′; antisense primer, 3′-AAA AAC CAA GAG GTG CTG GG-5′) and a primer set for the other E2F1 site at −789 (sense 5′-CCC CTG CTC ACT ACC CAT CT-3′; antisense, 3′-AAA AGA GGG AAG CCC TCA C-5′).
MTT assay
An MTT assay was used to analyze the proliferation of UC5, 5637, MCF7, and MDA-MB231 cells in E2F1 and NELL2 expression. For plasmid transfection experiments, cells (5000 cells/well), E2F1, NELL2, and the combined expression vector were plated in 96-well culture plates for 24 h. Briefly, 20 μL of the MTT reagent (5 mg/mL) was added to each well for 2 h at 37°C. After addition of 100 μL DMSO, the absorbance of each well was then measured using a Wallac Victor 1420 Multilabel Counter (EG&G Wallac) at a wavelength of 540 nm.
Invasion assay
The in vitro invasive properties of cell lines UC5, 5637, MCF7, and MDA-MB231 were studied using Boyden chambers (NeuroProbe) precoated with Matrigel (25 μL/mL; BD Biosciences) and incubated for 1 h at 37°C. Bottom wells were filled with 27 μL of media with a 5% serum medium (MDA-MB231 cells), 0.1% serum medium (5637 cells), and 10% serum medium (UC5, MDA-MB231 cells). A total of 1×104 cells/56 μL were seeded into the upper compartment and incubated for 24 h at 37°C and 5% CO2. After incubation for 24 h, cells in the upper surface of the filter were removed by using a cotton swab and those attached to the lower surface of the filters were stained using Diff-Quik reagents (Sysmex Co.) and counted (five fields/well). The invasion percentage was expressed as the percentage of invading cells through the Matrigel. A representative graph of six independent experiments is reported.
Statistical analysis
The results were analyzed with a one-way analysis of variance followed by the Student–Neuman–Keuls multiple comparison test for unequal replications. The Student's t-test was used to compare two groups.
Results
Overexpression of NELL2 and E2F1 in human invasive cancer cell lines
To find potential regulators of NELL2 expression, we analyzed the sequence of the promoter region of NELL2 and found two putative E2F1 binding sites. For these E2F1 binding sites, we used TTTSSCGC as a consensus sequence (Xu et al., 2011; Zou et al., 2012), where S represents C or G. Because of the E2F1 activity in tumorigenesis (Tsantoulis et al., 2005; Chen et al., 2009; Lee et al., 2010), we determined NELL2 expression in various breast (MCF7, MDA-MB231, and MDA-MB435) and bladder (UC5, EJ, 5637, and T24) cancer cell lines. The 5637 and MDA-MB231 cells showed more invasive activity than UC5 and MCF7 cancer cells by an invasion assay with Boyden chambers (Materials and Methods section).
Higher expression of E2F1 and NELL2 was found in various invasive breast cancer cell lines (MDA-MB231 and MDA-MB435) and bladder cancer cell lines (EJ, 5637, T24) than in the other two noninvasive cell lines (UC5 and MCF7) by quantitative real-time PCR (Fig. 1A, B). The relative protein levels of E2F1 and NELL2 in these cell lines were confirmed by Western blot analysis (Fig. 1C, D). These results suggest that upregulation of E2F1 and NELL2 mRNA may be involved in the invasion activity seen in tumor progression. Therefore, we used these two cell lines (5637 and MDA-MB231) in further functional studies.
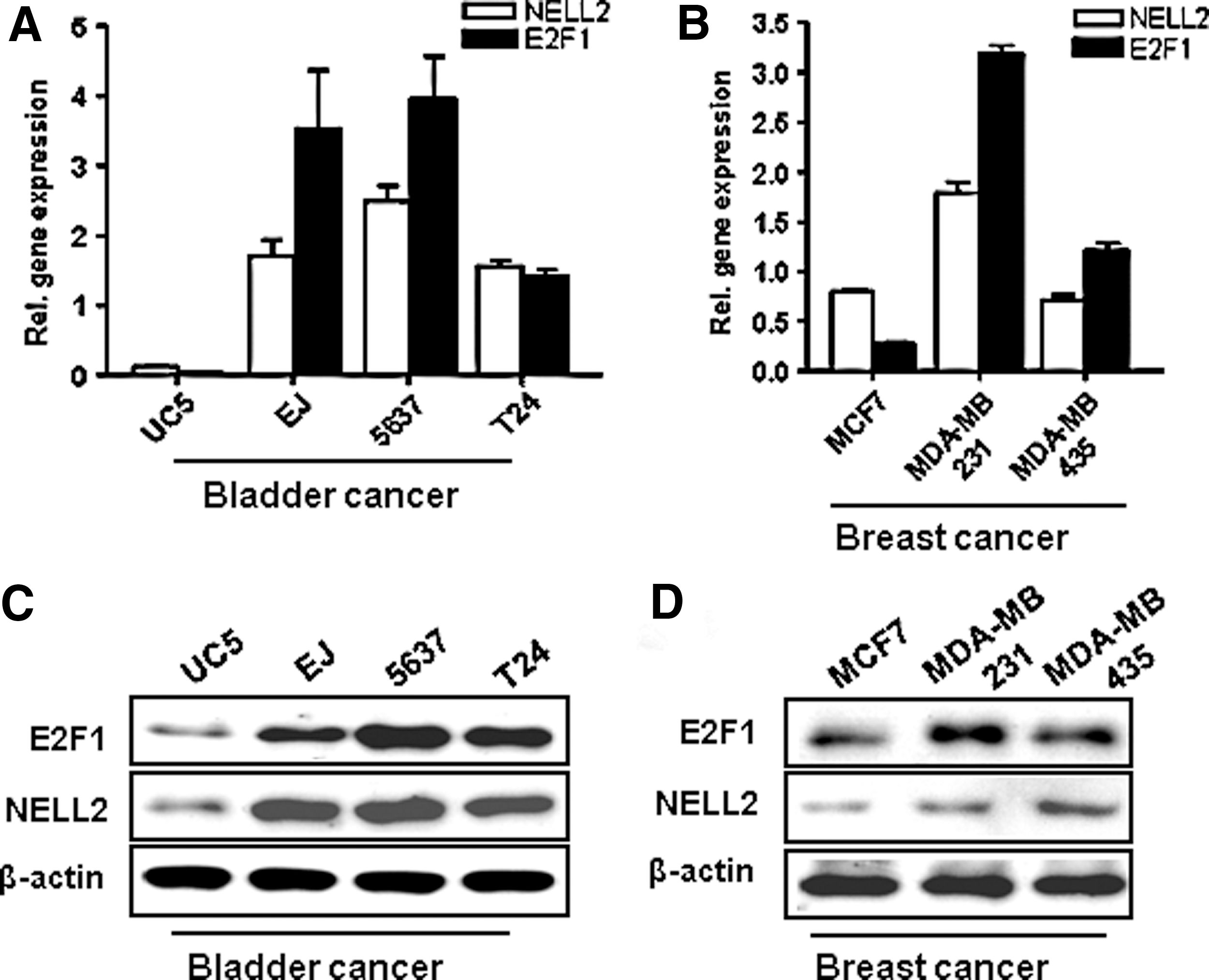
Expression of E2F1 and NELL2 mRNA and protein in bladder and breast cancer cell lines. The expression of mRNA and protein of E2F1
Upregulation of NELL2 in an E2F1-dependent manner in cancer cells
To determine whether E2F1 directly regulates transcription of the NELL2 gene, we used the NELL2 promoter to drive a luciferase reporter gene in transient cotransfections with an E2F1 expression plasmid in 5637 and MDA-MB231 cells. Ectopic E2F1 strongly upregulated transcription from the NELL2 promoter in both cell lines (Fig. 2A, B). For further determination of in vivo interaction between E2F1 and two potential E2F1 binding sites in the NELL2 promoter region, ChIP assays were performed using the E2F1 antibody. The precipitated DNA was amplified using the PCR primer sets specific to the promoter regions containing the E2F1 binding site (Fig. 2C, D); one primer set amplified a 98 bp fragment of the NELL2 promoter region from −147 to −49 encompassing the E2F1 site at −73 (−73 to −65); the other primer set amplified a 159 bp fragment from −879 to −720 of the NELL2 promoter with the E2F1 site at −789 (−789 to −781). As shown in Figure 2C and D, the appropriate NELL2 promoter region was immunoprecipitated with the E2F1 antibody. The results revealed that PCR fragments containing the E2F1 sequences at −73 and −789 were markedly increased in DNA samples from E2F1 transfected cells compared with DNA from pcDNA transfected cells. No detectable band was observed in the control IgG precipitations.

Interaction of the NELL2 promoter with E2F1. Determination of the promoter region of NELL2, which can be activated by E2F1.
To observe whether E2F1 affects the expression of endogenous NELL2, the mRNA and protein of NELL2 were analyzed in E2F1 overexpressing 5637 (Fig. 3A, E) and MDA-MB231 cells (Fig. 3B, F) by quantitative real-time PCR and Western blot analysis. Equal amounts of total RNA were used to perform quantitative real-time PCR with primer pairs specific to NELL2 and actin (control). Transient expression of E2F1 enhanced NELL2 mRNA expression compared to vector control (Fig. 3A, B). And the result was confirmed by the relative expression of E2F1 and NELL2 in these cell lines as seen by Western blot analysis (Fig. 3C, D). As shown in Figure 3C and D, the NELL2 protein level increased in E2F1 overexpressing cells. These results demonstrate that E2F1 can upregulate the expression of NELL2 mRNA and protein. In addition, to test whether downregulation of E2F1 affects NELL2 expression, we used siRNA against E2F1 to reduce the expression level of E2F1 in 5637 and MDA-MB231 cells. Downregulation of E2F1 by transfected siRNA (Fig. 3C, D, G, H) significantly decreased the expression of NELL2 RNA (Fig. 3C, D) and protein (Fig. 3G, H).
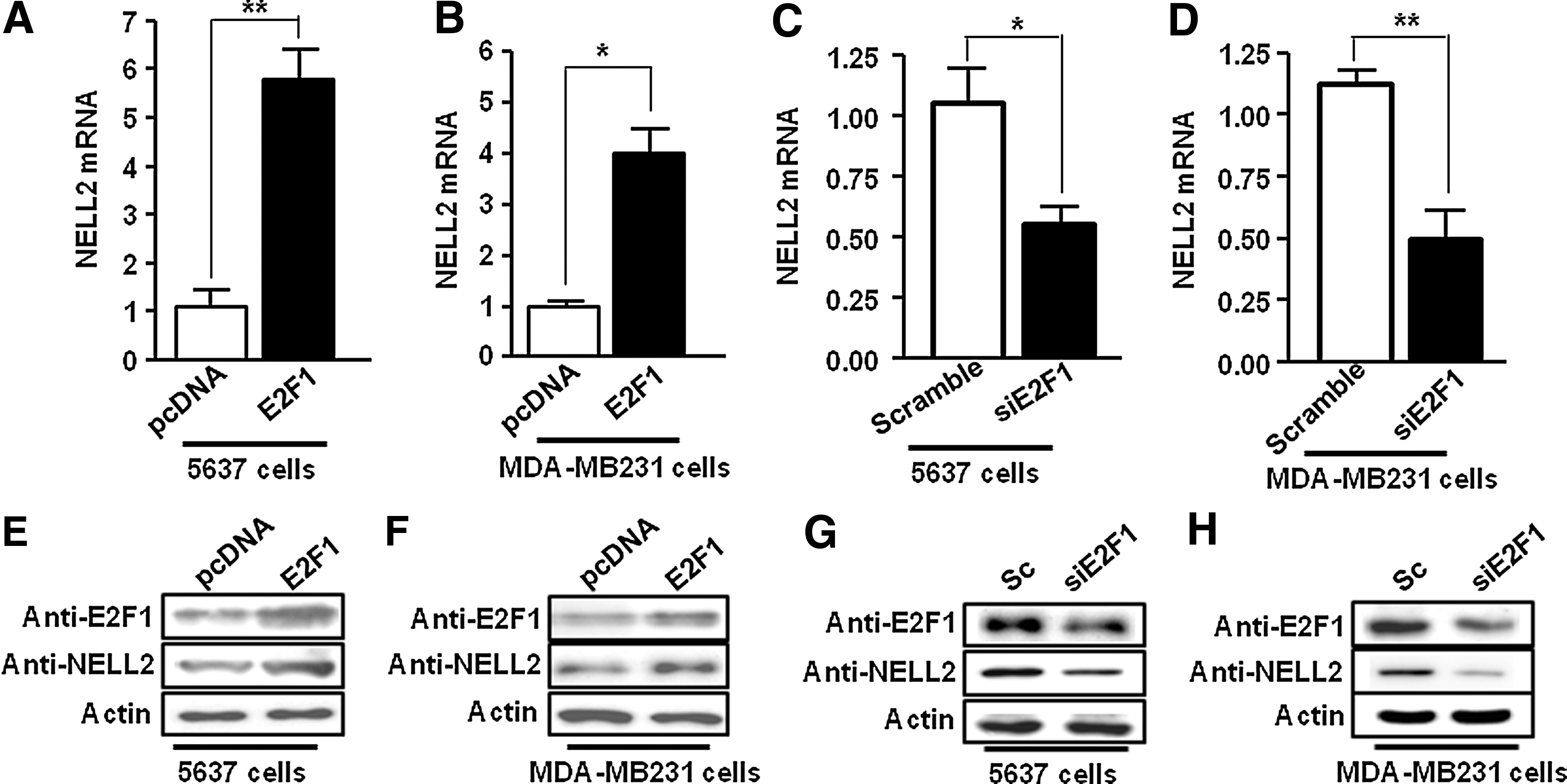
Regulation of NELL2 by E2F1. Overexpression of E2F1 leads to increased expression of NELL2.
NELL2 and E2F1 enhances the proliferation of cancer cells
We examined whether NELL2 was involved in the promotion of proliferation of breast and bladder cancer cells in a manner mediated by E2F1. An MTT assay was performed to assess cell viability following E2F1 and NELL2 overexpression. As shown in Figure 4, the absorbance values increased in cells transfected with E2F1 and NELL2 expression constructs. UC5 cells (Fig. 4A), 5637 cells (Fig. 4B), MCF7 cells (Fig. 4C), and MDA-MB231 (Fig. 4D) cells were transfected with a control vector, a NELL2 expression vector, a E2F1 expression vector or cotransfected with NELL2+E2F1 expression vectors, incubated for 24 h, and subjected to the MTT assay. We found that cancer cell viability in the overexpression group was significantly higher than in the control group. These results suggested that overexpression of E2F1 and NELL2 might be related to the increase in breast and bladder cancer cell proliferation.

Effect of NELL2 and E2F1 on proliferation of cancer cells. Cells were seed at 5×103 cells per well in 96-well plates. Cells were transfected with E2F1, NELL2, and E2F1+NELL2 expression vectors for 24 h. Cell viability was tested at the indicated times using an MTT cell proliferation assay. UC5 cells
Overexpression of NELL2 and E2F1 increased invasiveness of cancer cells
An invasion assay was carried out to assess whether NELL2 plays a role in E2F1-mediated invasion by UC5, 5637, MCF7, and MDA-MB231 cancer cells. The four cancer cells were transfected with a control vector, a NELL2 expression vector, an E2F1 expression vector, and NELL2+E2F1 expression vectors, incubated for 24 h, and then were plated in Boyden chambers. Invading cells could be visualized using the Diff Quik solution staining of membranes (Fig. 5A). The overexpression of NELL2 or E2F1 showed an increase of invasiveness in four cell lines. Moreover, an enhanced effect from cotransformation with E2F1 and NELL2 was detected in transfected cells (Fig. 5). These results are consistent with the high expression levels in invasive cancer cell lines (Fig. 1). Therefore, NELL2 is a possible mediator of the E2F1 activity and may be involved in promoting proliferation as well as invasion by bladder and breast cancer cells.
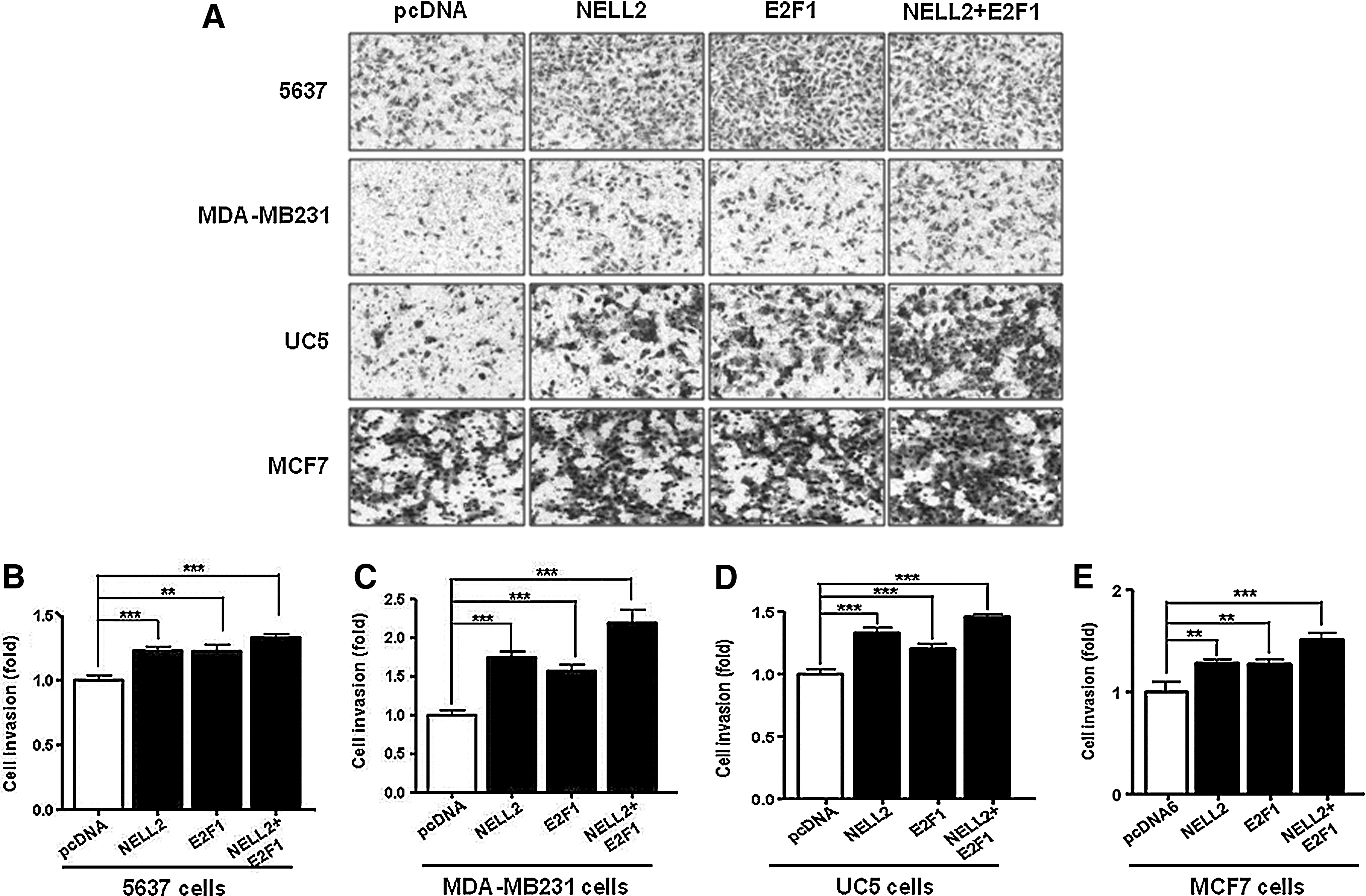
Effect on cancer cell invasion by NELL2 and E2F1. NELL2 was able to potently promote invasion by bladder and breast cancer cells after transfection of E2F1 as seen in a Boyden chamber assay. Invading cells could be visualized using the Diff Quik solution staining of the membranes
Discussion
NELL2 was initially characterized as a gene required for proper development of the nervous system and it can alter the behavior of both central nervous system and peripheral nervous system progenitor cells (Oyasu et al., 2000; Kim et al., 2002; Matsuyama, 2004; Matsuyama, 2005; Nelson et al., 2004). We have previously reported that NELL2 is transactivated by E2F1 and that NELL2 mediates the survival promoting effect of E2F1 in neuroprogenitor cells (Maeda et al., 2001). Thus, NELL2 regulates various factors that assist neuronal cell survival in the central nervous system and peripheral nervous system. Moreover, NELL2 proteins also participate in the growth and proliferation of cancer cell lines (DeGregori et al., 1995; Kuroda et al., 1999). Despite the extensive studies on the biological properties of NELL2, very little is known about the mechanisms involved in the transcriptional regulation of NELL2.
In this study, we demonstrated the key role of E2F1 in activating NELL2 expression. We first identified two E2F1 binding sites (5′-TTTCCCGC-3′) (Xu et al., 2011; Zou et al., 2012), one located between −73 to −65 and the other located between −789 to −781 in the NELL2 promoter. Because a putative SP1 site, which may be involved in mediating E2F1-dependent transcriptional regulation (Xu et al., 2011), is also found in this region, we checked the binding activity of E2F1 at the SP1 site. The results show that E2F1 could regulate NELL2 expression by directly binding to its promoter, but the SP1 site at −536 on the NELL2 promoter was not detected by ChIP assay (Fig. 2). Furthermore, overexpression of E2F1 could increase the NELL2 promoter activity, suggesting the relevance of E2F1-mediated regulation of NELL2 in vivo (Fig. 2).
The transcription factor E2F1 has a role in promoting cell proliferation by regulating genes controlling cell cycle progression and DNA synthesis (DeGregori et al., 1995; Lee et al., 2010; Li et al., 2012). Here we described a new function of NELL2 expression in human cancer cell lines (Fig. 1B, D), with implications for understanding the mechanisms of tumor-associated growth regulation. E2F1 activated NELL2 and promoted the invasive activity of cancer cells (Fig. 4). Therefore, we suggest that E2F1 is a transcriptional regulator of NELL2 and this interaction could be amplified in invasive tumors. The detailed cellular mechanism underlying this effect remains to be further identified.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Korean government (MEST; 2012-0000481). S.I.K. was partially supported by a grant (T33414) from the Korea Basic Science Institute.
Disclosure Statement
The authors declare no competing financial interests.