
Abstract
Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels are expressed in dorsal root ganglion (DRG) neurons, which are involved in diverse mechanisms that regulate DRG functions. Protein kinase A (PKA) is an essential kinase that plays a key role in almost all types of cells; it regulates the ion channel activity, the intracellular Ca2+ concentration, as well as modulates cellular signals transduction. Nevertheless, the effect of PKA inhibition on the HCN channel activity in DRG neuron remains to be elucidated. Here we investigated the impact of PKA inhibition on the HCN channel activity and DRG neurons excitability. Our patch-clamp experiments both under whole-cell and single-channel conditions demonstrated that PKA inhibition with KT5720, a cell membrane permeable PKA-specific inhibitor, significantly attenuated HCN channel currents. Current clamp recording on freshly isolated DRG neurons showed KT5720 reduced overshoot amplitude and enhanced the threshold of the action potential. Moreover, our live-cell Ca2+ imaging experiments illustrated KT5720 markedly reduced the intracellular Ca2+ level. Collectively, this is the first report that addresses KT5720 attenuates the HCN channel activity and intracellular Ca2+, thus reducing DRG neurons excitability. Therefore, our data strongly suggest that PKA is a potential target for curing HCN and DRG neuron relevant diseases.
Introduction
HCN channels were extensively expressed in the heart, brain, and nerve systems (Brown et al., 1979; Shi et al., 1999; Demontis et al., 2002; Bender et al., 2003; Holderith et al., 2003). Its expression patterns are roughly changed between the different parts of the body and species (Moosmang et al., 1999; Monteggia et al., 2000; Santoro et al., 2000), this expression variety is probably implicated with different physiological functions. HCN channel encoded current Ih is involved in regulating a variety of neuron biophysical functions, including setting the resting membrane potential (Gauss and Seifert, 2000), controlling neurons excitability (Kase and Imoto, 2012), mediating action potentials (APs) (Hogan and Poroli, 2008), governing motor learning (Lewis et al., 2011), regulating synaptic transmission, and dendritic integration as well (Atkinson and Williams, 2009). Thus, it is not surprising that Ih plays an important role in both physiological and pathophysiological conditions. For example, the HCN channel plays a vital role in developing neocortex after birth (Battefeld et al., 2012); In a hypertrophic heart, increased Ih prolongs the ventricular AP repolarization (Hofmann et al., 2012); The HCN channel is a potential drug target for the treatment of epilepsy (Shah et al., 2013); HCN2 channels play a central role in inflammatory and neuropathic pains, its mutation is potentially associated with generalized epilepsy with recessive inheritance as well (DiFrancesco et al., 2011; Emery et al., 2011).
To date, emerging experimental evidences indicate all four HCN channel subunits are expressed in murine dorsal root ganglion (DRG) neurons (Kouranova et al., 2008), and previous studies have well documented that the HCN channel activity is regulated by intracellular cAMP (Almanza et al., 2012), The cAMP/protein kinase A (PKA) is a critical pathway that controls a serial of cellular processes, recent reports showed regulating PKA has feedback on intracellular cAMP levels (Vandamme et al., 2012). Hence, it is legitimate to postulate that PKA plays a role in regulating the HCN channel activity. However, currently available literatures provide controversial results. PKA inhibitor H-8 demonstrated significantly attenuated Ih in bull-frog sympathetic neurons (Tokimasa and Akasu, 1990), but the same compound was identified to have no effect on HCN channel currents in canine purkinje fibers (Chang et al., 1991). To address this issue, here, on freshly isolated rat DRG neurons, we chose KT5720, a membrane permeable selective PKA inhibitor, combined with cutting edge technologies, including whole-cell and single-channel patch-clamp recording, and live-cell calcium imaging. Our results demonstrated that KT5720 dramatically inhibited Ih in both whole-cell and single-channel conditions. Furthermore, we provided evidences that KT5720 decreased the intracellular calcium concentration in freshly isolated rat DRG neurons. Our data unveiled that PKA plays a key role in regulating the HCN channel activity and DRG neuron excitability, so it may represents a potential target in the treatment of HCN channel-related diseases.
Materials and Methods
Thirty-eight adult male Sprague-Dawley Rats with an average weight of 248.6±13.9 g were used in this study. All the animal operation procedures were reviewed and approved by the Animal Care Committee of Huazhong University of Science & Technology; it also abided by the NIH guidelines.
DRG neurons isolation
Rats were euthanized with CO2, followed by neck dislocation. The back skin was incised, and then the backbone was quickly removed and put into a dish with oxygen saturated Dulbecco's Modified Eagle's Medium (DMEM) solution. After that, the backbone was cut into two parts along the midline, DRG was carefully taken out with a hairspring tweezer, the attached tissue was removed with an iris scissor, and the supernatant was removed by a clean straw. Then, the DRG tissue was completely cut into pieces with a spring scissor. The tissue was then put into a glass tube that contained enzymes, followed by gently mixing and percussing with a pipette. The tube was then put into a 37°C shaking bath for 15 min, wind and percussion several times every 5 min, and then phenylmethanesulfonyl fluoride was added into the suspension to terminate digestion. The suspension was then centrifugalized in 1000 r/min for 5 min, and then the supernatant was removed. Next, a 5 mL fresh DMEM was added into the tube and single DRG neurons were mixed well with a Pasteur pipette. The DRG neuron suspension was then dropped onto coverslips, which had been coated with poly-l-lysine. After that, these coverslips were placed in a 37°C CO2 incubator for 30 min to let the cells adhere tightly, 2.5 mL of the DMEM was then added into each dish plus waiting for 1 h. DRG neurons obtained from these procedures were used for experiments.
Electrophysiological whole-cell patch-clamp recording
Whole-cell currents were recorded with Axopatch 200B, Digidata 1440A, and pClamp 10.2 software (Molecular Devices, Union city, CA), signals have been filtered with an eight-pole Bessel filter 900CT/9L8L (Frequency Devices, Ottawa, IL). Ten millimeter length borosilicate glasses (Sutter Instruments, Novato, CA) were used to prepare for the patch pipettes. The pipettes were pulled with a Narishige PP-830 Two-step puller, and then polished with a Micro Forge MF-830 fire polisher (Narishige Group, Tokyo, Japan) to obtain a final resistance of 3–5 MΩ.
Electrophysiological single-channel patch-clamp recording
Single-channel recording was implemented in a cell-attached mode. Fire-polished borosilicate glass pipettes with a final resistance of 8–10 MΩ were chosen for recording, those seal resistance ≥10 GΩ were considered for data analyses, 10 μM ZD7288 was added to ensure that the recorded single-channel currents were Ih (data did not show). Only those rounds, shinny looking DRG neurons were chosen for experiments. All single-channel experiments were implemented at room temperature 25°C–26°C.
Live-cell calcium imaging
For intracellular Ca2+ imaging experiments, 250 μL poly-l-lysine was added to each 35-mm-diameter glass bottom dish, incubated at 37°C, poly-l-lysine was put on the coverslip and incubated for 1 h, then dishes were aspirated and washed with 2 mL sterile water three times. Cell suspension was added into the dishes, kept at room temperature for 30 min to let the cells adhere to the bottom of the dishes. The Fura 2-AM solution was diluted with a bath solution to a final concentration of 2 μM. The supernatant was then removed from the dishes, 250 μL of the Fura 2-AM working solution was added into the dishes, and stored in darkness at room temperature for 30 min. The Fura 2-AM working solution was then removed and cells were washed with a bath solution 3 times. Ca2+ imaging at ×40 oil objective was acquired with an OLYMPUS IX81 motorized inverted research microscope, the corresponding software was MetaFluor 7.7.2.0.
Solutions and drugs
The DRG tissue-containing solution has the following compositions (in mg/mL): DMEM 10 pH 7.4 with NaOH. The DRG digestion solution contains (in mg/mL) DMEM 10, type I collagenase 1, trypsin 0.5 pH 7.4 with NaOH. The whole-cell patch-clamp recording bath solution contains (in mM) NaCl 80, KCl 5, BaCl2 2, NaH2CO3 26, CoCl2 2, TEAC 40, CaCl2 1, 4-AP 4, CdCl2 0.2, D-Glucose 10 pH 7.4 with NaOH, 1 μM TTX was added to block irrelevant sodium channels. The whole-cell patch-clamp recording pipette solution contains (in mM) NaCl 5, K+-gluconate 135, ATP-Mg2+ 2, GTP-Na+ 0.4, EGTA 0.5, HEPES 10 pH 7.3 with KOH. The single-channel patch-clamp recording bath solution contains (in mM) potassium gluconate 100, KCl 50, NaCl 4, CaCl2 1, EGTA 5, HEPES 10, and glucose 10 pH 7.25 with KOH. The single-channel patch-clamp recording pipette solution contains (in mM) NaCl 140, KCl 5, CaCl2 2, MgCl2 1, HEPES 10 pH 7.4 with NaOH, plus 1 μM TTX, 10 mM TEA, 0.2 mM CdCl2, 10 μM picrotoxin was supplemented to block other voltage- and ligand-gated channels. All the compounds and chemicals were purchased from Sigma.
Data analysis and statistics
Clampfit 10.2 software (Molecular Devices, LLC, Sunnyvale, CA) was used for the patch-clamp electrophysiological data analyses. SigmaPlot 10.0 software (Systat software, Inc., Chicago, IL) and Igor Pro 5.0.1 (WaveMetrics, Inc., Lake Oswego, OR) were used for statistical analysis and figure illustration. τ was fitted with a monoexponential equation. In step protocol, the currents were taken from the averaged last 100 ms recording. The single-channel current amplitudes were calculated from all points histograms, and the mean single-channel conductance values were calculated from the current–voltage curve. The results were summarized as means±SEM from cell numbers n and animal numbers N. Data were compared with two-way ANOVA or the paired two-tailed student's t-test. p-value less than 0.05 was considered statistically significant.
Results
The selective PKA inhibitor KT5720 attenuates Ih in freshly isolated rat DRG neurons
The effect of PKA inhibition on DRG neuron Ih was investigated by the conventional whole-cell patch-clamp technique. At the holding potential of −40 mV, whole-cell Ih was elicited with a protocol that hyperpolarized from −60 to −140 mV in −10 mV interval, the hyperpolarize stimulation maintained for 2 s, then returned to the holding potential for 1 s. The cell capacitance was kept unchanged during the recording procedure. In the control condition, Ih was recorded without adding any compound, followed with perfusion with a 5 mL 3 μM KT5720 solution into the recording chamber, which dramatically attenuated the whole-cell Ih . At the end of recording, 10 μM ZD7288 (a selective HCN channel inhibitor) was added into the bath solution, in the presence of ZD7288, the remained inward current was totally inhibited (Fig. 1A). To confirm that the effect of KT5720 on Ih being attributed to PKA inhibition, the same experiments were carried out with H89, it had the same effect as KT5720 (Fig. 1B), which confirmed KT5720 attenuates Ih via PKA inhibition. Furthermore, to test whether KT5720 blocks Ih through HCN channel inhibition, ZD7288 was applied to DRG neurons in the absence of KT5720. The result was consistent with our hypothesis, ZD7288 blocked whole-cell Ih .
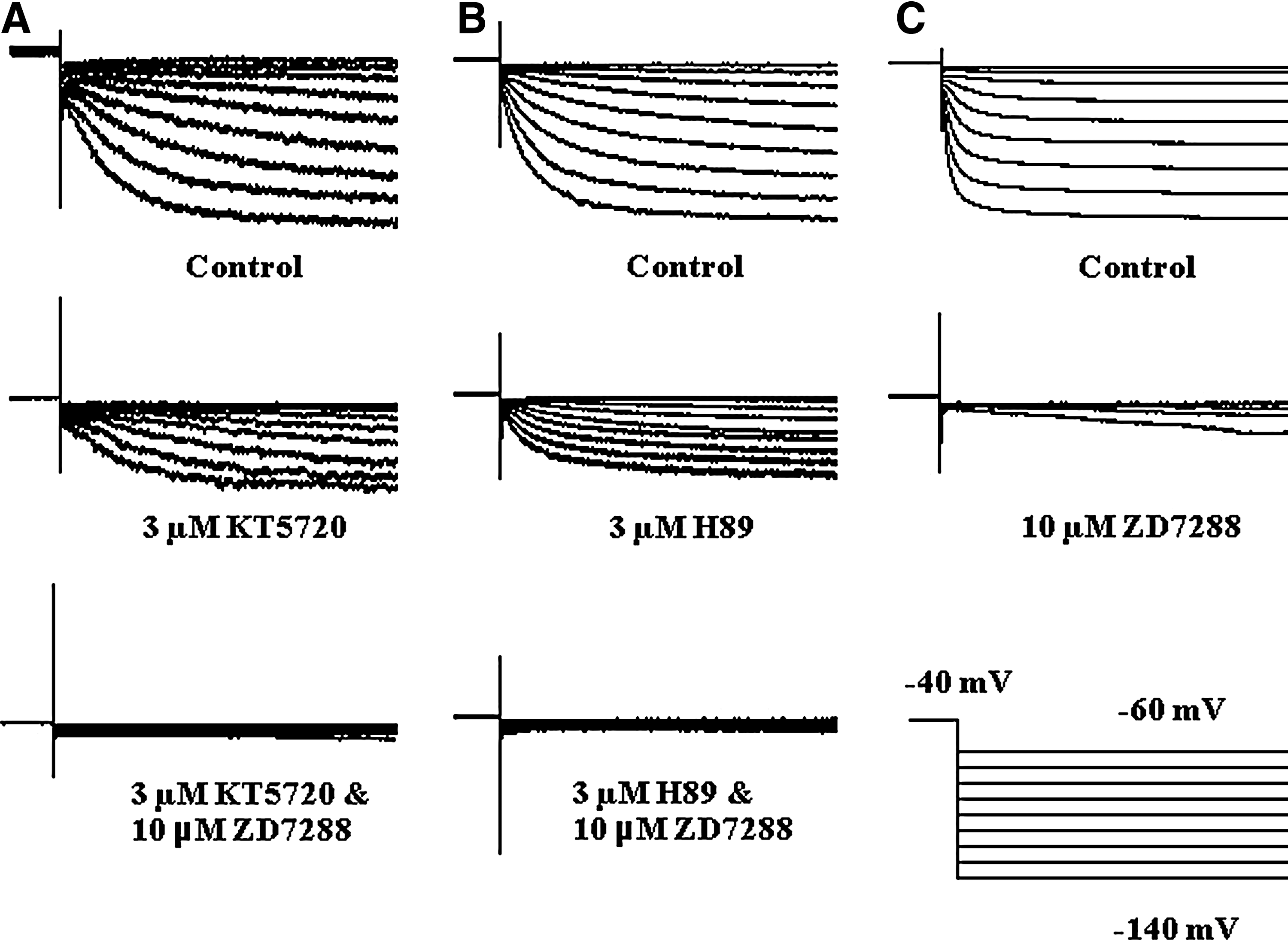
Whole-cell recording of Ih
on freshly isolated DRG neurons.
HCN channel activation curve was leftward shifted and its current density was significantly reduced by PKA inhibition
In the above paragraph, we already demonstrated that PKA inhibition with KT5720 induced a dramatic reduction of Ih . This inhibition not only influences the current amplitudes, but also has an impact on the channel active threshold. The aftermath is the HCN channel activation curve shifted to the negative direction. Those points were obtained from the tail current amplitudes following application of hyperpolarizing voltage steps, and were normalized to the largest one at −140 mV. The activation curves have been fitted with a Boltzmann function. As illustrated in Fig. 2B, the HCN channel V1/2 changed from −81.91 mV (control) to −100.85 mV (KT5720) (n=9, N=6). Furthermore, in the presence of 3 μM KT5720, the Ih density was declined as well. To ensure that the HCN channel has been fully activated, we chose −140 mV as the statistical analysis voltage, the current amplitudes averaged from the last 100 ms of the step-stimulation. Statistic results showed that the Ih density decreased from 16.82±1.77 (control) to 3.57±0.52 (KT5720) (n=6, N=5; ***p<0.005) (Fig. 2C).
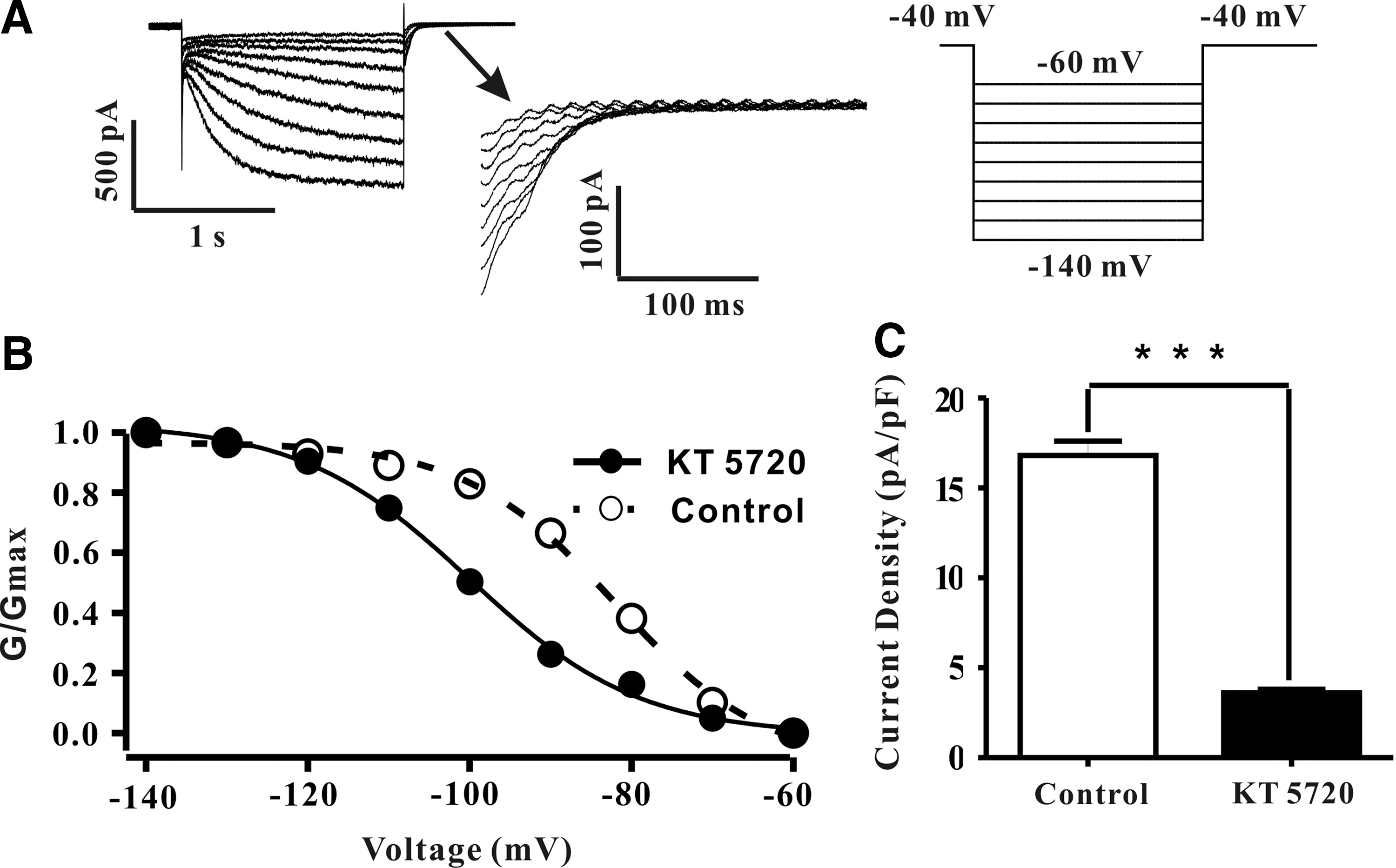
KT5720 significantly shifted HCN channel activation curve to the hyperpolarization direction and decreased its current density.
KT5720 (3 μM) slows down HCN channel activation kinetics
Since KT5720 (3 μM) attenuated Ih
amplitude and increased its activation threshold, thus shifting the V1/2
to more negative potentials. We further postulated that PKA inhibition with KT5720 can slow down Ih
activation kinetics. Ih
was elicited with a sustained −130 mV stimulation for 2 s. To explore the effect of KT5720 on Ih
activation, we first compared the original recording between control and KT5720 (3 μM). The activation time constant was fitted with a monoexponential equation (I=Io+Ae−(x−x
o
)/τ
), within which,
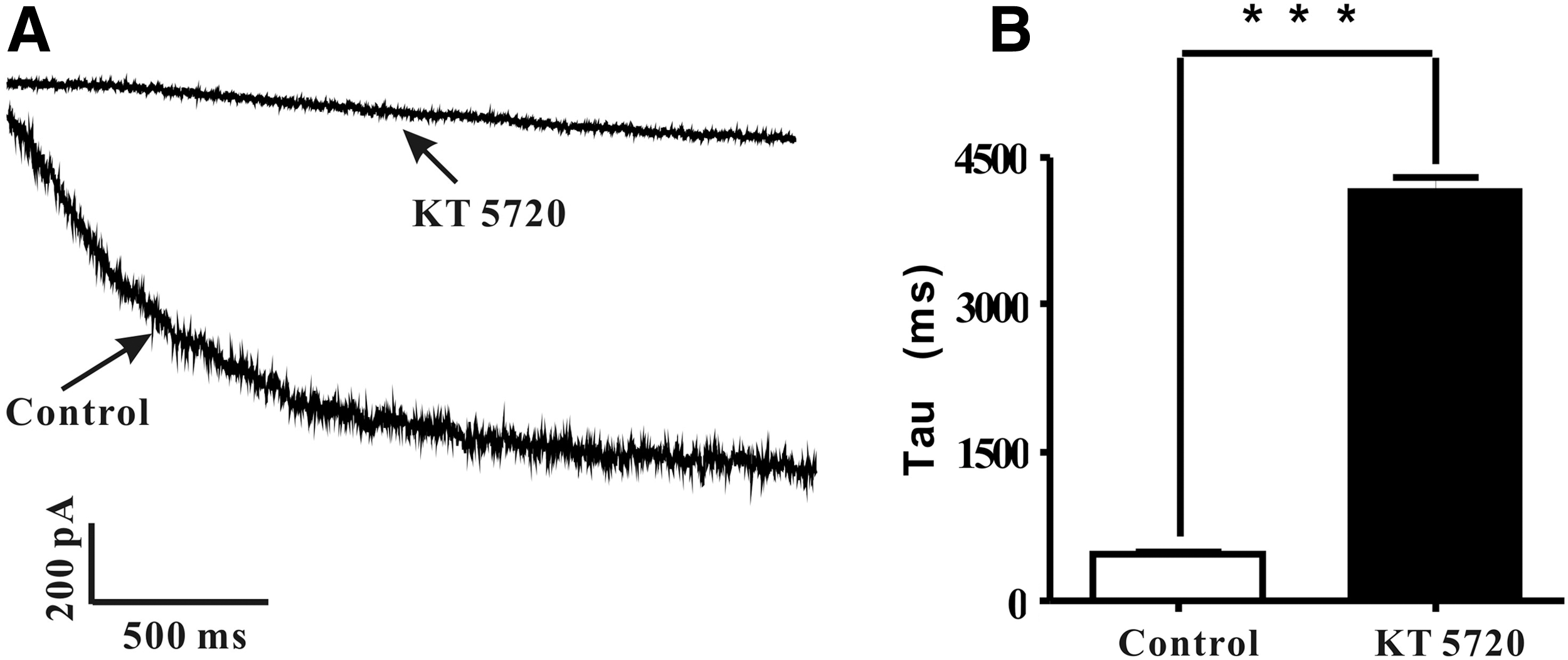
The activation time constant of HCN channel was dramatically increased by 3 μM KT5720.
PKA inhibition with KT5720 (3 μM) reduced DRG neurons excitability
At the end of AP, it was believed that Ih can depolarize the membrane potential to the threshold thus inducing a new AP (Accili et al., 2002). It also reported that the HCN channel selective inhibitor ZD7288 can attenuate DRG neuron excitability through inhibiting AP (Chaplan et al., 2003). We further hypothesize that KT5720 inhibiting Ih may also have an impact on DRG neuron excitability, therefore, current-clamp recordings were implemented on six rats as shown in Figure 4A. APs were evoked by a 2 s depolarize current, which confirmed our hypothesis. As demonstrated in Figure 4A, after application with 3 μM KT5720, the APs were profoundly inhibited. Furthermore, AP recording was also carried out in the presence of 10 μM ZD7288. As illustrated in Figure 4B, KT5720 did not have a significant effect under this condition, which confirmed that KT5720 attenuates APs via HCN channel inhibition. AP threshold was changed from −25.25±1.21 mV under control condition to −22.42±1.54 mV in the presence of 3 μM KT5720 (n=6, N=6) (Fig. 4C). Moreover, AP overshoot amplitude significantly decreased from 23.77±2.12 mV (control) to 6.93±1.39 mV (KT5720) (n=6, N=6; **p<0.01) (Fig. 4D).
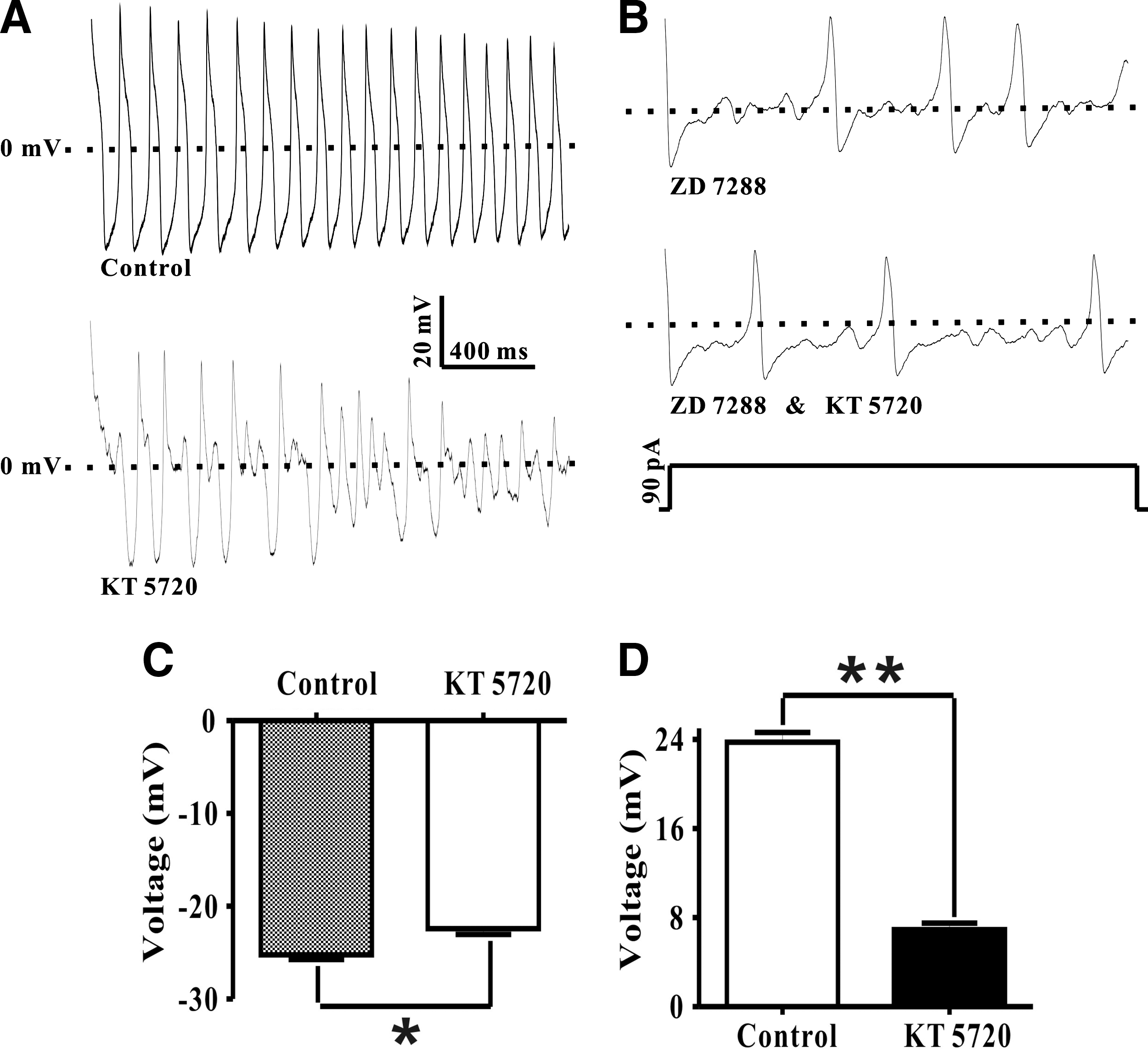
The effect of KT5720 on DRG neuron action potential (AP).
The effect of KT5720 on HCN single-channel properties
To the best of our knowledge, no published literatures reported single-channel recording on freshly isolated DRG neurons so far. Therefore, to further reveal the mechanism of the PKA pathway on regulating HCN channel activation, cell-attached single-channel recording experiments were implemented as illustrated in Figure 5A. To eliminate irrelevant impacts, only those original gigaseal ≥10 GΩ were considered for further recording. After gigaseal formation, 15 more minutes were waited to ensure the seal was stable. Moreover, no rundown was observed under control condition, with 10 μM ZD7288 added to ensure the recorded currents were Ih (data did not show). The open probability has a voltage-dependent characteristic, which showed it increased from 0.21 at −50 mV to 0.91 at −140 mV, and the V1/2 was −82.18 mV (Fig. 5), which was consistent with our whole-cell recording result and the published report as well (Simeone et al., 2005). In the presence of 3 μM KT5720, the voltage-dependent characteristic of Po was totally dampened. At −90 mV, KT5720 inhibited Po for 71%, it also slightly attenuated the current amplitude from 1.06±0.18 pA (control) to 1.02±0.13 pA (KT5720). Moreover, before and after application of 3 μM KT5720, HCN channel conductance decreased from 12.41±0.69 pS to 11.54±0.76 pS, but except for Po neither the changes of current amplitude nor channel conductance were statistical significant (n=13, N=9) (Table 1). So, our data showed that PKA inhibition with KT5720 reduced HCN channel Po.
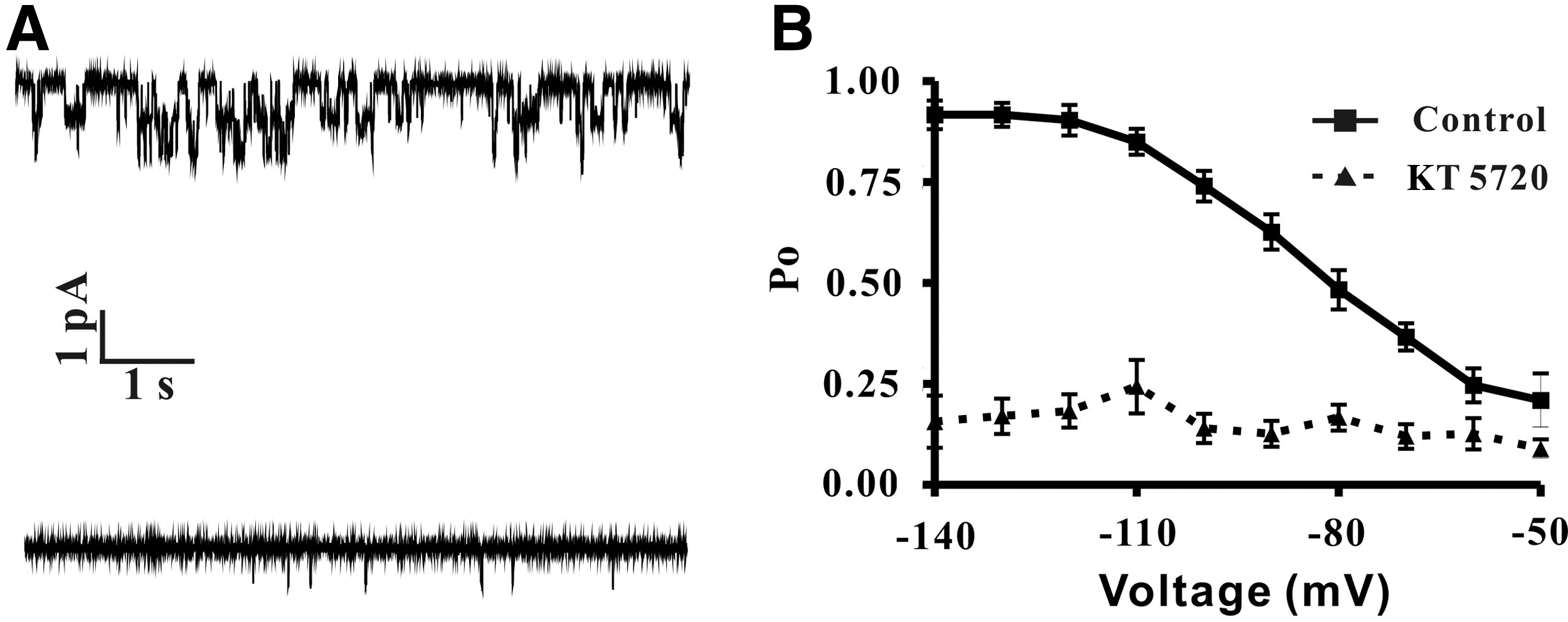
PKA inhibition induced reduction of HCN single-channel open probability.
, p<0.01.
HCN, hyperpolarization-activated cyclic nucleotide gated.
PKA inhibition with KT5720 (3 μM) reduced intracellular Ca2+ concentration
Previous study showed that the HCN channel activity was relevant to intracellular Ca2+ (Hagiwara and Irisawa, 1989), which hints PKA-induced Ih inhibition may be attributed to the change of the intracellular Ca2+ level. To explore the effect of KT5720 on DRG neuron intracellular Ca2+ level, real-time calcium imaging experiments were implemented on freshly isolated DRG neurons. Fura 2-AM (a ratiometric and UV light-excitable dye) was used as the intracellular Ca2+ indicator, the dual-wave excitation UV lights were generated with a TILL's Polychrome V light source. Recording coverslips were coated with poly-l-lysine to fix the cells. At first, the experiment presented in Figure 6A was performed. In the control part, great care was taken to ensure that no intracellular degradation occurred. As showed in Figure 6A, application of KT5720 (3 μM) profoundly reduced the intracellular Ca2+ level, similar results as illustrated in Figure 6A also were obtained from 12 other cells. The fluorescent ratio (340/380 nm) under control conditions was 0.80±0.05 (n=13, N=4). Thereafter, the fluorescent ratio decreased to 0.64±0.06 in the presence of 3 μM KT5720 (n=13, N=4; Fig. 6B). Thus, application of KT5720 reduces the DRG neuron intracellular Ca2+ level, so, PKA inhibition induced Ih attenuation, at least partly, due to reduction of intracellular Ca2+.
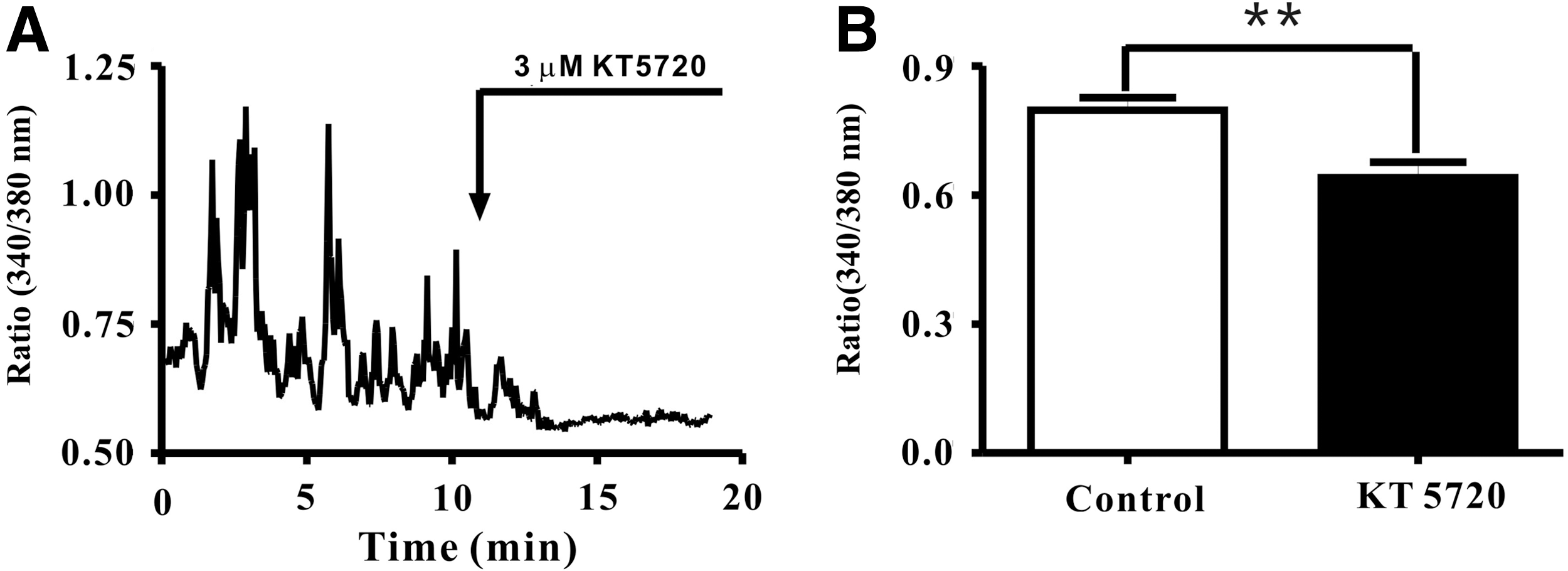
KT5720 (3 μM) induced DRG neuron intracellular Ca2+ concentration reduction.
Discussion
In this study, we provided electrophysiological and calcium imaging data, which strongly suggest that PKA is a pivotal regulator of the HCN channel, thus controling DRG neuron excitability. To the best of our knowledge, our study is the first to illuminate the physiological role and effect of PKA inhibition on the HCN channel activity on freshly isolated DRG neurons. Based on our data, we can draw these conclusions: (1) KT5720 inhibits HCN channel whole-cell currents, Ih thereafter has a more hyperpolarized V1/2 , a decreased current density, and a slower activation time constant as well; (2) The PKA inhibitor KT5720 inhibits HCN single-channel currents, significantly reduced the Po value, which is consistent with our whole-cell experiment result; (3) KT5720 attenuates Ih associated with the reduction of intracellular Ca2+ concentration, which further induced DRG neuron excitability reduction.
We observed robust attenuation of whole-cell HCN channel currents by the PKA inhibitor KT5720 (Fig. 1). When compared the parameters between control and in the presence of KT5720 conditions, the V1/2 decreased 18 mV, which is in line with the published data, which addressed the effect of PKA on the HCN channel activity in mice sinoatrial nodes (Liao et al., 2010). It is well known that cAMP is a critical and ubiquitous messenger that is involved in many intracellular regulation mechanisms (Kronstad et al., 2011; Lania et al., 2012; Billington et al., 2013). An increased intracellular cAMP level was reported potentiated both transferred and native Ih (Gao et al., 2012; Zong et al., 2012). On the contrary, reduction of the cAMP concentration would induce Ih decline (Chen et al., 2005). However, whether the cAMP/PKA pathway is involved in regulation of the HCN channel activity is still under controversy. For instance, Constantin's group reported that the Gonadotropin-Releasing Hormone-1 (GnRH-1) Neuronal activity was regulated by PKA, while independent of HCN channels. Specifically, the HCN channel protein was identified in GnRH-1 neurons, blockade of HCN channel with ZD7288 significantly attenuated the efficiency of forskolin to stimulate GnRH-1 neurons, whereas PKA inhibition did not have this effect. In other words, PKA inhibition has no effect on HCN channel activity (Constantin and Wray, 2008). On the contrary, another study showed that PKA inhibition with H89 profoundly blocked Ih in cultured rat olfactory receptor neurons (Vargas and Lucero, 2002).To clarify this issue, we carried out electrophysiological recording with the compound of KT5720 on freshly isolated rat DRG neurons.
Whole-cell patch-clamp data analyses demonstrated for the first time that KT5720 inhibiting Ih resulted in both the current density (Fig. 2) and the channel active kinetics reduction (Fig. 3). By analyzing the current density, we identified that it decreased 78.77% in the presence of KT5720. In addition, the HCN channel activation time constant increased 796.5% (Fig. 3). PKA-dependent inhibition of transferred TRPC5 channel was reported in HEK cells (Sung et al., 2011), and previous study showed KT5720 inhibits rapid component I(Kr) of cardiac delayed rectifier potassium currents in guinea pigs as well (Wang et al., 2009). However, the effect of PKA inhibition on HCN channel activity remains to be fully explored. Here we revealed the key role of PKA regulating the HCN channel activity by conducting whole-cell patch-clamp experiments on freshly isolated DRG neurons.
DRG neurons excitability is essential to many important physiological procedures. For instance, it is well established that the pathophysiology of neuropathic pain is associated with altered DRG neuron excitability (Dib-Hajj et al., 2008; Miller et al., 2009); a rodent study indicated that DRG neuronal excitability also has impacts on visceral hypersensitivity (Xu et al., 2012); as well as regulating DRG neuronal excitability can change the sensory innervation of the urinary bladder thus controlling the bladder's storage and void urine (Sun et al., 2010). Furthermore, a number of ion channels have been reported to be involved in regulating DRG neuron excitability. Impaired Na(v)1.7 induced human DRG neuron hyperexcitability (Han et al., 2012). A recent study in rat DRG neurons indicated that brain-derived neurotrophic factor levels increased under nerve injured conditions would suppress the BK channel activity, which further induced DRG neuron abnormal hyperexcitability (Cao et al., 2012). Selective Kv subunit inhibition demonstrated to be able to enhance DRG neuron excitability too (Lee et al., 2012). It was unanimously agreed that the HCN channel was extensively expressed in DRG neurons (Kouranova et al., 2008), combined with its unique characteristics, which strongly hints that the HCN channel probably plays a key role in regulating DRG neuron function and more attention should be deserved to be paid on this subject. However, the function of the HCN channel and associated cAMP/PKA pathway in DRG neuron excitability remains to be elucidated. To date, only a handful of groups have carried out very preliminary studies on this topic (Wan, 2008; Wang et al., 2012; Weng et al., 2012). To address this critical issue, we conducted whole-cell current-clamp recording on freshly isolated DRG neurons. Our data confirmed our hypothesis that KT5720 not only robustly inhibited AP overshot amplitude, but also increased its threshold (Fig. 4), which therefore indicated HCN channel and associated cAMP/PKA pathway were critical in regulating DRG neuron excitability.
To further investigate the effect of PKA inhibition with KT5720 in attenuating the HCN channel activity, cell-attached patch-clamp single-channel recording was implemented. Our result showed KT5720 significantly decreased the Po value of the HCN channel (Fig. 5), which is compatible with our whole-cell recording results, and further confirmed our hypothesis that KT5720 selectively inhibits PKA would attenuate the HCN channel activity. Therefore, PKA is an important electrophysiological target to regulate the HCN channel activity. To date, HCN single-channel biophysical properties in native DRG neurons are still largely unknown. Those investigations conducted on recombination, cardimyocyte and atrialmyocyte (Michels et al., 2005, 2008a, 2008b) cannot be transferred to freshly isolated DRG neurons directly. Here we reported a HCN single-channel conductance of ∼12 pS, which is in line with the study conducted also with a cell-attached mode (Michels et al., 2005). We chose the cell-attached mode, which can prevent dialysis and thereby maintain the intracellular circumstances; thus, our results have more electrophysiological significances.
The Ca2+ signal is the most important second messenger, which is relevant to both normal and pathophysiological processes (Leybaert and Sanderson, 2012; Skupin and Thurley, 2012). Recent study revealed PKA inhibition blocked T-type Ca2+ currents and reduced membrane excitability in mice DRG neurons (Wang et al., 2011). However, the effect of PKA inhibition on freshly isolated rat DRG neuron cellular Ca2+ remains to be determined. Our result showed KT5720 (3 μM) reduced intracellular Ca2+ levels on freshly isolated DRG neurons, which is in agreement with studies on neonatal rats (Hou and Wang, 2001). In addition, recent study revealed that Ca2+-activated adenylyl cyclases is a new player on regulating HCN channel function (Kryukova et al., 2012), and its combination with HCN2 could generate more efficient biological pacing (Boink et al., 2012). Moreover, evidence indicated that HCN channel can directly conduct Ca2+, although tiny, it contributes significantly the relevant biophysical functions (Michels et al., 2008a). Taken together, even though our study cannot exclude the possibility that other ion channels and receptors may also be involved in this regulation, our effort provided the initial evidence that intracellular Ca2+ involved in the PKA inhibition induced Ih attenuation process.
In summary, this is the first study that addressed the effect of PKA inhibition on the HCN channel activity and further elucidated the effect on DRG neuron excitability. Our whole-cell and single-channel electrophysiological data indicated that PKA inhibition robustly attenuated the HCN channel activity. We provided further evidence that KT5720 selectively inhibits PKA reduced the intracellular Ca2+ level and further reduced DRG neuron excitability. Thus, our data strongly suggests PKA is a pivotal mediator of the HCN channel activity and DRG excitability. Therefore, it may represent a potential pharmacological target for the treatment of HCN channel and DRG neuron-relevant diseases.
Footnotes
Acknowledgments
The authors thank Ms. Yuli Chen and Mr. Xu Li for their kindly help.
Disclosure Statement
There is no conflict of interest between the authors.