
Abstract
Physiological occlusal force constitutively exists in the oral environment and is important for periodontal homeostasis and remodeling. Cyclic tensile stress (CTS) triggers the biological response of periodontal ligament (PDL). However, a few reports have studied the correlation between CTS during physiological occlusal force and PDL cell activities such as osteogenic differentiation. In the present study, human PDL cells (hPDLCs) were subjected to 10% elongation CTS loading at 0.5 Hz for 24 h, which represents the physiological conditions of occlusal force. Gene expression microarray was used to investigate the mechano-induced differential gene profile and pathway analysis in vitro. The osteogenic relative factors, that is, SPP1, RUNX2, and SP7, were assessed by real-time PCR and Western blot. The involvement of mitogen-activated protein kinase (MAPK) signaling pathways was investigated by Western blot with a specific inhibitor. The expressions of SPP1, RUNX2, SP7, p-ERK1/2, and p-Elk1 were up-regulated after 10% CTS exposure. However, these up-regulated expressions were prevented by ERK1/2 inhibitor U0126 in the physiological occlusal force-applied hPDLCs. These results showed that 10% CTS could enhance osteogenic differentiation of hPDLCs via ERK1/2-Elk1 MAPK pathway, indicating that CTS during physiological occlusal force is a potent agent for PDL remodeling.
Introduction
To clarify the effect of mechanical stress on PDLCs, many studies have used different systems to mimic the stress in an oral environment to observe the different molecular expression. However, the expression patterns of these molecules in response differed according to the modes of the mechanical stress, such as shear, compression, and tension as well as the magnitudes of the stress (Redlich et al., 2004; Ozaki et al., 2005; Garlet et al., 2007; Kook et al., 2009). Previous evidence has shown that cyclic tensile stress (CTS) during orthodontic tooth movement (OTM) increases the expression of osteogenic genes and proteins such as alkaline phosphatase (ALP), secreted phosphoprotein 1 (SPP1; also known as bone sialoprotein I/osteopontin), transcription factor Sp7 (SP7; also known as osterix), and runt-related transcription factor 2 (RUNX2) of hPDLCs (Wescott et al., 2007; Cho et al., 2010; Tang et al., 2012). However, ignoring physiological occlusal force constitutively exists in the oral environment and is important for periodontal homeostasis and remodeling. So, studying the effect of CTS during physiological occlusal force on the osteogenic differentiation of human PDLCs (hPDLCs) could extend our understanding of the effect of mechanical stress on PDL remodeling.
Previous findings suggested that the mitogen-activated protein kinases (MAPKs) controlled a range of cellular activities, such as embryogenesis, differentiation, proliferation, and death (Zhang and Liu, 2002; Chambard et al., 2007; Kook et al., 2011a). The direct involvement of MAPKs on mechano transduction has been largely suggested in muscle cells (Li et al., 2000; Zampetaki et al., 2005) and in bone cells (Franceschi and Xiao, 2003; Liedert et al., 2006). In periodontium, centrifugal force during OTM up-regulated COL I expression in human periodontal fibroblasts through the activation of MAPK-AP-1signaling (Kook et al., 2009; Hong et al., 2010). However, little knowledge is available about the mechanism of physiological occlusal force on hPDLCs.
In this study, we examined the effect of 10% CTS, which represents physiological occlusal force (Matsuda et al., 1998a; Fujihara et al., 2010) on the osteogenic differentiation of hPDLCs using gene expression microarray. In addition, the molecular mechanism of 10% CTS enhancing the osteogenic differentiation of hPDLCs was also investigated.
Materials and Methods
Cell culture
PDL tissues were gently scraped from the middle third of periodontally healthy, noncarious premolar tooth roots, extracted from donors aged between 12 and 20 years of age for orthodontic reasons with informed consent. Teeth were washed, and fragments of the PDL attached to the middle third of the root were removed with a scalpel. Tissue explants were plated onto 25 cm2 cell culture flasks (Corning) in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 10% fetal calf serum (Gibco) and antibiotic reagent (10,000 units penicillin,10,000 mg streptomycin; Gibco). In the third passage, cells were stained with vimentin and cytokeratin antibodies for characterization. Cells at passages 3 to 6 were used for mechanical loading. Collection and culture of hPDLCs was approved by the Ethical Committee of Nanjing Medical University. The different batches of cells were applied in the same assays to avoid the effect of the donor on the outcome.
Application of CTS on hPDLCs
hPDLCs suspension (1.5×105 cells/well) were spread into six-well, 35 mm flexible-bottomed BioFlex® Culture Plates coated with type I collagen. When the cells reached 80% confluence, the plates were subjected to an in-plane cyclic deformation of 10% elongation with 0.5 Hz (30 cycles/min) for 12, 24, and 48 h in a standard Bioflex baseplate linked to a Flexcell® FX-5000™ Tension Unit (Flexcell Corp.). This force represented the physiological conditions of occlusal force (Matsuda et al., 1998a; Fujihara et al., 2010). For the control experiments, hPDLCs were cultured in the same plates without any mechanical stress.
Gene expression microarray analysis
Total RNA in each group was isolated using TRIzol reagent (Invitrogen). The concentration and purity of the RNA samples were determined by the absorbance of RNA at 260 and 280 nm, respectively. O.D. A260/A280 ratios between 1.8 and 2.1 are acceptable. For microarray analysis, cells were subjected to 10% elongation CTS loading at 0.5 Hz for 24 h. The microarray analysis was carried out in accordance with the NimbleGen Gene Expression Analysis protocol (Roche NimbleGen, Inc.). All gene level files were imported into Agilent GeneSpring GX software (version 11.5.1) for further analysis. In this study, a 1.5-fold change was used as the cut-off value, and those genes showing changes that were greater than 1.5 fold were identified as differentially expressed. Then, gene ontology (GO) analysis and pathway analysis based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database were used for further investigation on the differential genes of interest. The microarrays were carried out in triplicate.
Real-time PCR analysis
Real-time PCR was performed using FastStart Universal SYBR Green Master (Roche) in a quantitative PCR System (ABI 7300). Gene expressions were normalized to the housekeeping gene Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), which was used as an internal control. The primer sequences for SPP1, RUNX2, SP7, and GAPDH were presented in Table 1. The cycling conditions were 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 58°C for 1 min. Gene expression levels were calculated using the 2−ΔΔCt method. Data were expressed as mean±standard deviation of three independent experiments.
Western blot analysis
Cells were collected, washed, and lysed in RIPA lysis buffer (Beyotime) containing 1 mM phenylmethylsulfonyl fluoride (PMSF; Beyotime). After centrifugation, the supernatant was collected and assayed quantitatively with a BCA protein assay kit (Beyotime). Equal amounts of protein extracts (30 μg/sample) were separated by 12% SDS-PAGE and blotted onto polyvinyl difluoride membranes (PVDF; Millipore) at 300 mA for 1 h in a blotting apparatus (Bio-Rad). The blots were incubated, respectively, with primary antibodies (SPP1, 1:800, Abcam; RUNX2, 1:800, Abcam; SP7, 1:800, Abcam; GAPDH, 1:800, Bioworld; ERK1/2, 1:1000, Cell signaling technology; p-ERK1/2, 1:1000, Cell signaling technology; ERK5, 1:1000, Cell signaling technology; p-ERK5, 1:1000, Cell signaling technology; JNK, 1:1000, Cell signaling technology; p-JNK, 1:1000, Cell signaling technology; P38, 1:1000, Cell signaling technology; p-P38, 1:1000, Cell signaling technology; p-Elk, 1:800, Santa Cruz; and GAPDH, 1:800, Bioworld) overnight at 4°C. Finally, the membranes were incubated with appropriate horseradish peroxidase-conjugated secondary antibodies (1:10,000; Boster Biotech. Co. Ltd.) at room temperature for an additional 1 h. The membranes were developed with Immobilon Western Chemiluminescent HRP Substrate (Millipore) and were exposed to X-ray film (Eastman-Kodak). Band intensity was calculated using Gel-Pro Analyzer 4. ERK1/2 MAPK inhibitor, and U0126 was purchased from Beyotime. GAPDH serves as the internal control in these experiments. This experiment was repeated in triplicate.
Statistical analysis
Unless specified otherwise, all data are expressed as the mean±standard deviation (SD). A one-way analysis of variance (ANOVA) (SPSS version 17.0 software) was used for multiple comparisons. A value of p<0.05 was considered statistically significant.
Results
Effect of 10% CTS on hPDLCs
Difference in cellular orientation was observed as early as 12 h after the application of CTS. As shown in Figure 1, control cells remained randomly orientated (Fig. 1A). However, mechanically stretched cells re-orientated along with the direction of the applied tensile force (Fig. 1B). There was no significant difference among cell morphology of 12, 24, and 48 h stress loading (data not shown). After the third passage, hPDLCs stained positively for vimentin, but negatively for cytokeratin, confirming their mesodermal origin (Fig. 1C, D).
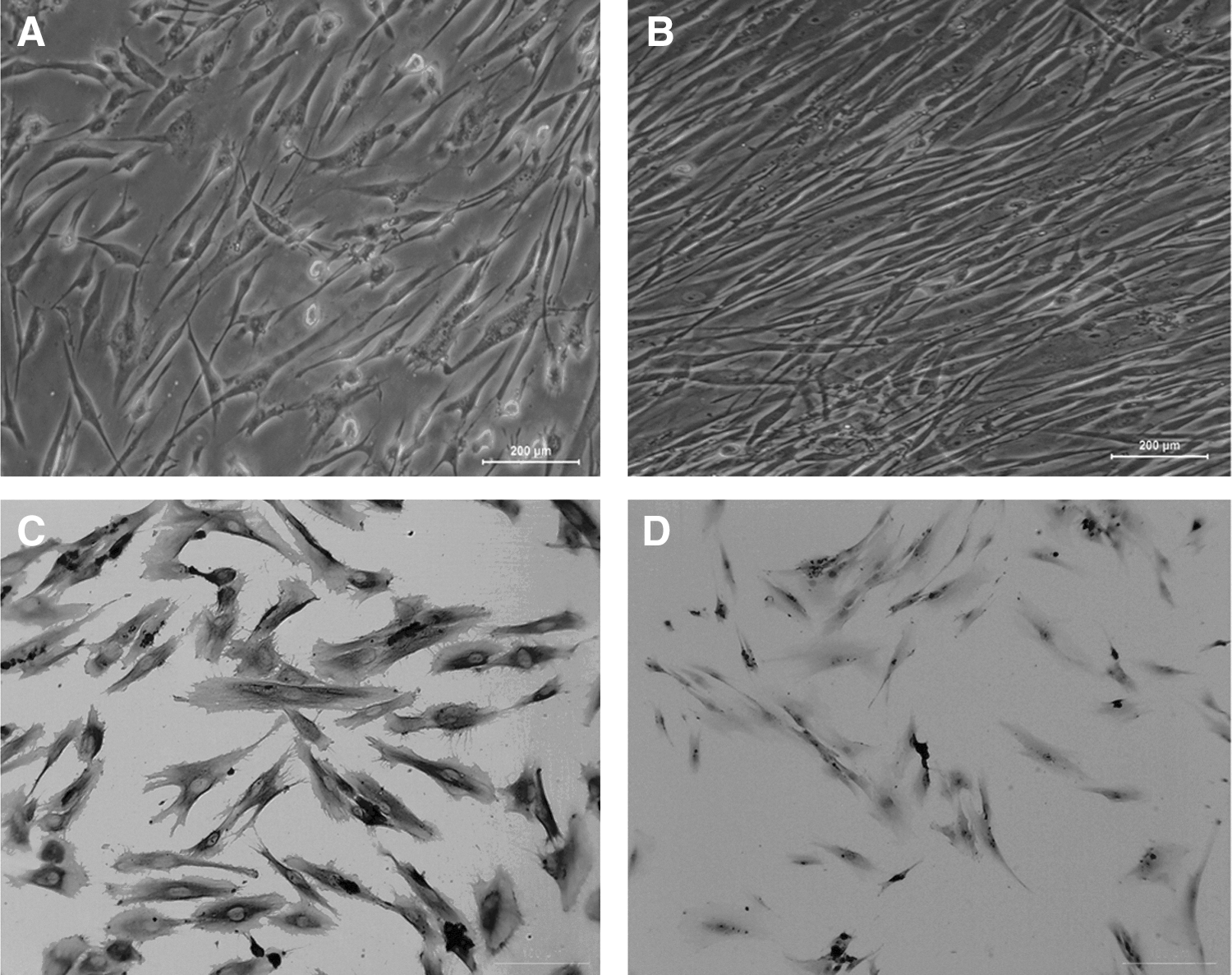
Effect of cyclic tensile stress on hPDLCs.
Identification of genes in response to 10% CTS
The microarray data showed that 453 out of 45,033 genes were differentially expressed. Among the 453 genes, 263 were up-regulated (Fold Change >1.5 and p<0.05), whereas 190 were down-regulated (p<0.05). The differentially expressed genes were partially summarized and listed in Table 2. GO analysis demonstrated that these differentially expressed genes participated in a variety of biological processes and molecular functions, such as regulation of cellular process, response to stimulus, cell proliferation, signal transduction, receptor binding, and multicellular organismal process. SPP1 in the up-regulated genes group with higher expression are involved in most of the biological processes, including response to stimulus, multicellular organismal process, negative regulation of cell proliferation, axon and neuron projection regeneration, regulation of cell growth, cell size, cell adhesion, and anatomical structure morphogenesis. For this reason coupled with SPP1 being one of the main bone-related components, the gene was selected as a gene of interest for further analysis in our study.
hPDLC, human periodontal ligament cells; Neg., negative.
Signaling pathway analysis of genes in response to 10% CTS
Based on the KEGG database, 43 signaling pathways associated with genes differentially expressed in response to CTS (p<0.05) were identified. The differentially expressed pathways were partially summarized and listed in Table 3.
10% CTS enhanced the osteogenic differentiation of hPDLCs
As shown in Figure 2A, RUNX2 mRNA was significantly up-regulated for 12 h CTS exposure, while the increase was significant decreased for 48 h loading as compared with CTS-untreated groups. SPP1 mRNA was significantly increased for 24 and 48 h CTS exposure. However, SP7 mRNA was down-regulated after CTS loading. Western blot results demonstrated that the protein levels of RUNX2 were significantly increased for 12 h and then decreased for 24 and 48 h CTS exposure as compared with CTS-untreated groups. SPP1 levels were elevated for 12 h CTS exposure with a peak increase for 24 h and then declined. Inconsistent with mRNA levels, SP7 level was up-regulated for 24 h CTS loading (Fig. 2B).
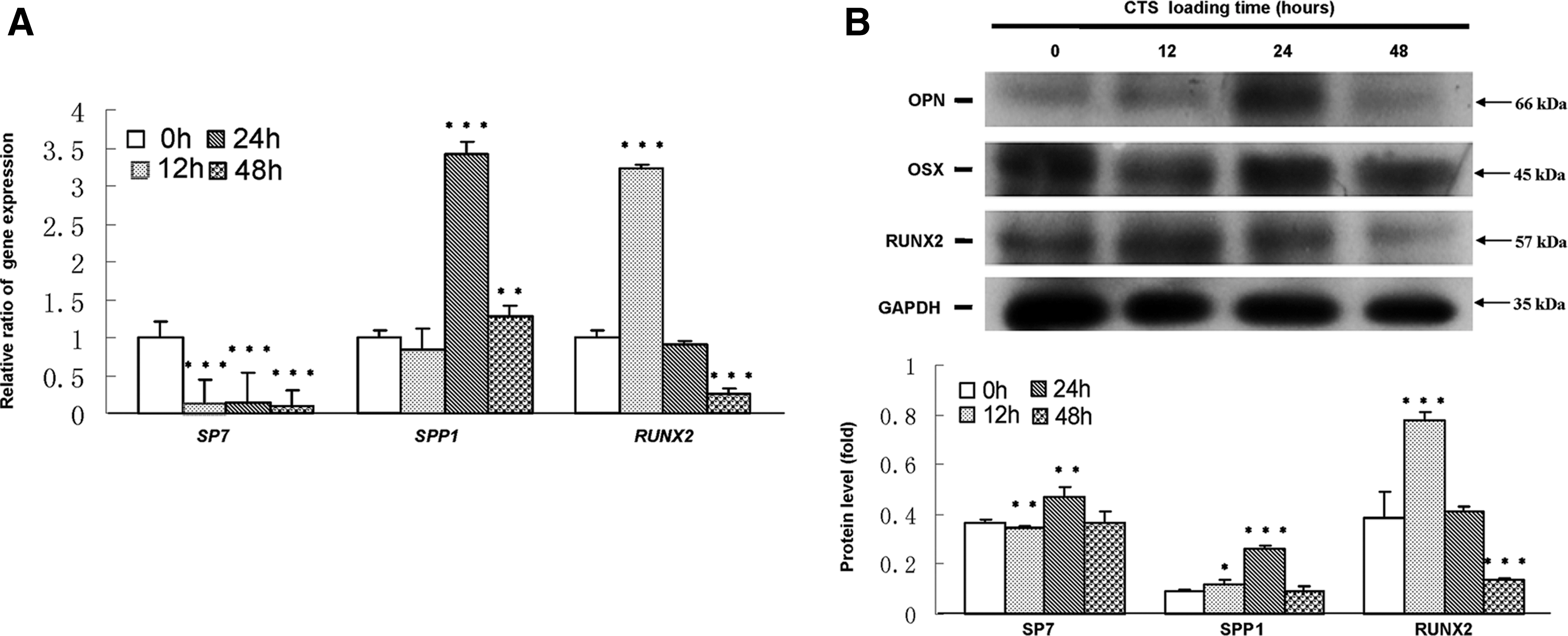
CTS up-regulated the expression of RUNX2, SP7, and SPP1 in hPDLCs.
10% CTS could activate ERK1/2-Elk1 MAPK pathway in hPDLCs
Based on pathway analysis, MAPK pathway was found to be related to osteogenic differentiation of hPDLCs induced by CTS. As shown in Figure 3, the levels of p-ERK1/2 (Fig. 3A) in CTS groups were up-regulated. Quantitatively, the p-ERK1/2 increased approximately 18-fold for 24 h, 14-fold for 12 h, and seven-fold for 48 h in CTS groups as compared with control groups. However, the levels of p-JNK, p-P38, and p-ERK5 were not affected (Fig. 3B–D). 10 μM ERK1/2 MAPK inhibitor U0126 was treated with hPDLCs for 1 h before the cells' exposure to CTS. Pretreatment with U0126 significantly prevented the CTS-induced increases of SPP1, RUNX2, SP7, p-ERK1/2, and p-Elk1 expression (Fig. 4).
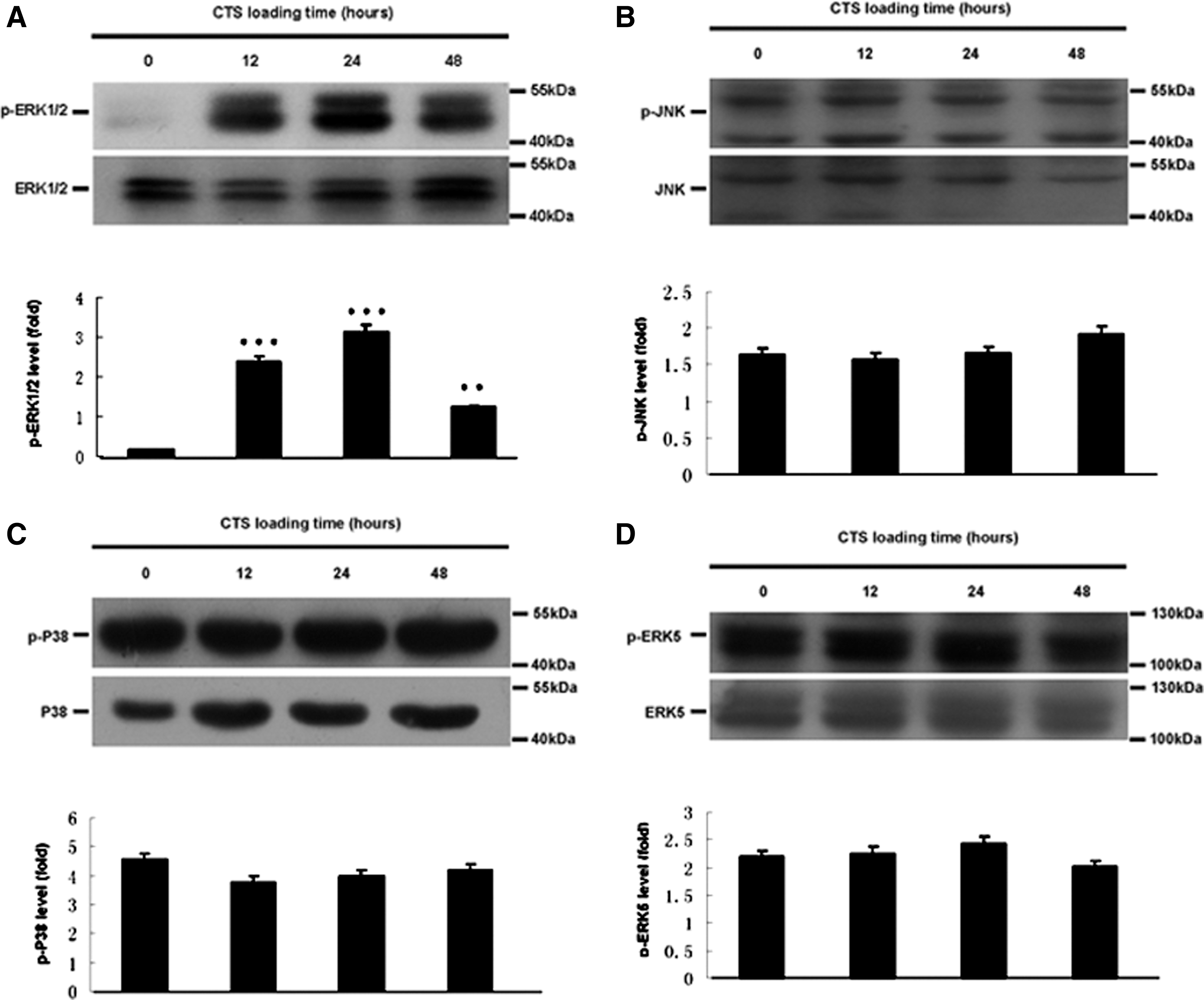
Effect of CTS on the MAPK pathway of hPDLCs.

Effect of ERK1/2 inhibitor on the phosphorylation of ERK1/2, Elk and the expression of SPP1, RUNX2, and SP7 in CTS-exposed hPDLCs.
Discussion
Cultured hPDLCs have been widely used to study the effects of mechanical stress on the expression of individual genes of interest (Saito et al., 1991; Shimizu et al., 1994; Howard et al., 1998; Chiba and Mitani, 2004; Yang et al., 2006; Nishijima et al., 2006; Wescott et al., 2007; Pavlidis et al., 2009; Tang et al., 2012). The results are equivocal depending on the culture conditions, stress regime, and the design of the mechanical deformation apparatus (Ozaki et al., 2005; Wescott et al., 2007; Saminathan et al., 2012). For example, cyclic tension has been reported to both inhibit (Chiba and Mitani, 2004) and stimulate (Yang et al., 2006) the synthesis of ALP. In the Flexcell system, a silicone membrane is stretched across a loading post by the application of vacuum pressure and depending on the shape of the loading post, either a biaxial or a uniaxial strain (as in the present study) may be applied to the cells. The silicone membrane is coated with type I collagen, which is more suitable for cell adhesion and simulates the in vivo environment. This system can have a better control of waveforms at low and high amplitudes through a computer-regulated bioreactor. Although the Flexcell system does have some limitations (Vande Geest et al., 2004; Wall et al., 2007; Milne et al., 2009; Saminathan et al., 2012), this system still remains the most effective way to screen cells for the expression of mechano-responsive gene in vitro methodology.
Laboratory evidence has already affirmed the significance of tensile strain during OTM with regard to the osteogenic differentiation of PDLCs. 3,000 μstrain cyclic uniaxial tensile stress significantly increased mRNA and protein expressions of STAB2, RUNX2, and OSX using a self-made system (Tang et al., 2012). 12% uniaxial tensile during OTM could express multiple genes involved in osteogenesis in hPDLCs (Wescott et al., 2007). However, a few researches are involved in physiological occlusal force, which constitutively exists in a normal oral environment and is important for PDL homeostasis and remodeling. Physiological occlusal force is an important symbol of the stomatognathic system. The homeostasis of periodontal tissue depends on the stimulation of occlusal force. On the contrary, loss of occlusal function caused atrophic changes in the PDL, such as narrowing of the space, disorientation of collagen fibers, and decreases in proteoglycans (Kaneko et al., 2001). Thus, it is important to clarify the physiological functions of mechanical stress with regard to the PDL, and our study aims at investigating the effect of CTS during physiological occlusal force on the behavior of hPDLCs.
Human gene expression microarray was employed in this study, which enables us to assess the whole genome transcriptome changes even though it would take us more time to clarify the whole content in the results. Previous studies have shown that cyclic tension loaded to hPDLCs would either increase (Yousefian et al., 1995; Matsuda et al., 1998b) or inhibit proliferation (Hao et al., 2009). In our microarray analysis, DPT, ASCL2, APOE, BTG1, NPPA, and SPP1, which were involved in negative regulation of cell proliferation, were up-regulated. As we know, slowing down proliferation will benefit cells to respond to external signals and then to affect differentiation (Lee et al., 2007; Wang et al., 2011). Therefore, from another aspect, the microarray results confirm that cells tend to differentiate rather than proliferate on being exposed to 10% CTS.
In the present microarray, SPP1 in the up-regulated genes group with higher expression was involved in most of the biological processes. SPP1 is an early marker of osteoblast differentiation and plays an important role in bone formation, resorption, and remodeling (McKee et al., 1993; Yang et al., 2010). SPP1 is thought to promote or regulate the adhesion, attachment, and spreading of osteoclasts to the bone surface during bone resorption (Standal et al., 2004). Previous studies have demonstrated that mechanical stress-induced expression of SPP1 is mostly composed of, or is present in, almost all osteocytes and in some osteoblasts and bone-lining cells at the resorption site in the early stage of experimental tooth movement (Terai et al., 1999; Kim et al., 2012). Therefore, we chose SPP1 as the major indicators to observe the osteogenic differentiation of hPDLCs under CTS. We demonstrated that the mRNA and protein expressions of SPP1 were up-regulated under 10% CTS in vitro. So, we hypothesized that SPP1 might act as a trigger for bone and cementum remodeling and is involved in osteogenic differentiation caused by 10% CTS.
In the current study, 10% CTS can exert some influence on the expression of osteoblast markers (RUNX2 and SP7) in hPDLCs, indicating that 10% CTS could enhance the osteogenic differentiation of these cells. RUNX2 and SP7 are necessary for the osteogenic differentiation (Komori, 2006; Baek et al., 2009). Genetic and molecular studies have shown that RUNX2 plays a key role in the osteogenesis and serves as an early transcriptional regulator of osteogenic differentiation (Takeda et al., 2001; Karsenty and Wagner, 2002). 10% CTS-treated hPDLCs exhibited higher gene and protein expression levels of RUNX2 for 12 h as well as RUNX2-positive cells were detected in the PDL during the first 3 days, suggesting that CTS can trigger the early-stage osteogenic differentiation of hPDLCs in vitro and in vivo. SP7 is vital in osteogenic differentiation as a result of its ability to regulate the expression of a number of important osteoblast marker genes such as SPP1, osteonectin, osteocalcin (OCN), bone sialoprotein (BSP), and collagen type I (COL I) (Nakashima et al., 2002). We detected that 10% CTS-treated hPDLCs exhibited higher protein expression levels of SP7 for 12, 24, and 48 h along with a large number of SP7-positive cells compared with RUNX2 and KI67 from day 3 to day 10, suggesting that CTS plays an important role not only in the early stage of osteogenesis but also in the later stage of bone formation. Interestingly, we observed that SP7 mRNA was decreased after 10% CTS loading from 12 to 48 h, which was inconsistent with other studies (Zhao et al., 2008; Tang et al., 2012). We hypothesized that 10% CTS might regulate SP7 mRNA, thus affecting its stability and leading to inconsistent transcript and protein levels. The relationship between mRNA and protein is not strictly linear, but has a more intrinsic and complex dependence, deviating from the classical view referred to as the molecular dogma. Different regulation mechanisms such as post-transcriptional processing, the degradation of the transcripts, translation, post-translational processing and modification, acting on both the synthesized mRNA and the synthesized protein, affect the amount of the two molecules differentially. So, it is not exactly the same as the transcription and translation levels. In addition, we noted that the significant expressions of SPP1, RUNX2, and SP7 were in a different CTS loading time in vitro and in vivo. It is speculated that these three factors were affected by CTS in different ways, and the specific mechanisms involved need further investigations.
The most important kinases involved in mechano transduction are the MAPKs, because most mechanical signals are transmitted to the nucleus through the activation of these kinases (Whitmarsh and Davis, 1996; Liedert et al., 2006). In our microarray analysis, an MAPK pathway was identified. Kook and Lee (2012) demonstrated that 1.5% elongation tension-stimulated expression of COL I and MMP-1 was controlled by the ERK/JNK-AP-1 and ERK-NF-κB signaling pathways and inhibited the proliferation of human PDL fibroblasts through Ras-p38 MAPK up-regulation. Human PDL fibroblasts stimulate osteoclastogenesis in response to compression force through TNF-alpha-mediated activation of CD4+ T cells (Kook et al., 2011b). Osteopontin expression in mechanical force-subjected PDL fibroblasts was activated by the ERK pathway (Hong et al., 2010). The inconsistent findings show that the mechanisms involved in force-applied hPDLCs also depend on the culture conditions and stress regime. Unlike the previous studies, our current findings showed that ERK1/2 not the JNK, P38, and ERK5 was a central mediator of mechanical signals in hPDLCs under 10% CTS during physiological occlusal force. Elk1, one of the Ets family of transcription factors, was a DNA-binding protein that activates transcription through its phosphorylation (Cruzalegui et al., 1999). ERK inhibitor blocked Elk1 phosphorylation as well as SPP1 gene expression stimulated by oscillatory fluid flow in human osteoblast-like cells (Wu et al., 2006). Therefore, we supposed whether the downstream mechano sensor of ERK1/2 was Elk1 or not. Our present results revealed that when CTS was applied to hPDLCs, the phosphorylation of Elk1 was increased, and this increase was significantly prevented by ERK1/2 inhibitor, suggesting that the cascade phosphorylation of ERK1/2 and Elk1 appeared to be, at least in part, associated with SPP1 expression in 10% CTS-exposed hPDLCs. Further studies are needed to clarify the relationship between ERK1/2/Elk1 and their target transcription factors in CTS-induced up-regulation of SPP1. In general, the microarray data is complex, and will take more time to be clarified. Next, more extensive studies are required to investigate whether other pathway mechanisms that are associated with physiological occlusal force enhance osteogenic differentiation of hPDLCs.
In conclusion, we may first demonstrate that physiological occlusal force induces ERK1/2-Elk1 MAPK signaling in the PDL, resulting in enhancement of the osteogenic differentiation of hPDLCs. These findings suggest that physiological occlusal force-induced ERK1/2-Elk1 MAPK signaling is involved in the homeostasis, remodeling, and regeneration of periodontal tissue.
Footnotes
Acknowledgments
This study was supported in part by grants from the Graduate Student Innovation Project of Jiangsu province (Grant No. CXZZ11_0732), National Natural Science Foundations of China (Grant No. 81170962, 81170981, and 81230022), Project of Science and Technology Department of Jiangsu province (Grant No. BK2011763), Ph.D. Programs Foundation of Ministry of Education of China (20113234110003), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Disclosure Statement
No competing financial interests exist.