
Abstract
Interleukin-6 (IL-6) signaling may play a causal role in the development of coronary heart disease. However, the relationship between IL-6 genotypes and plasma levels of IL-6 appears to be complex. To help clarify the inconsistent findings, we conducted a meta-analysis of the published genetic association studies of the −174 G/C polymorphisms in the IL-6 gene and the circulating IL-6 levels in a normal population. In this meta-analysis, no significant association of IL-6 −174G/C polymorphism and circulating IL-6 levels in a normal population was observed. However, when compared among GG, GC, and CC genotypes, heterogeneity existed among the studies. Sensitivity analysis revealed that, the independent study by Shen et al. influenced the heterogeneity in the homozygous and heterozygous comparison. Although Shen et al.'s study was excluded, no significant association was observed between IL-6 −174G/C polymorphism and circulating IL-6 levels in a normal population [homozygous comparison (GG vs. CC): the pooled standard mean difference (SMD) was −0.01, 95% confidence interval (CI): −0.1–0.08; heterozygous comparison (GC vs. GG or CC): the pooled SMD (GG vs. GC) was −0.05, 95%CI: −0.11–0.01, and the pooled SMD (CC vs. GC) was 0.03, 95%CI: −0.03–0.1]. Under the dominant model, the pooled SMD was −0.05, 95%CI: −0.11–0.01). The meta-analysis provides evidence that the −174G/C polymorphism in the IL-6 gene is not significantly associated with circulating IL-6 levels in a normal population.
Introduction
The circulating concentrations of IL-6 are likely to be influenced by several environmental and genetic factors, including a polymorphic site (−174G/C) in the IL-6 gene promoter (Fishman et al., 1998). Although the presented evidence favors the proinflammatory character of the −174GG genotype, some other data may raise doubts about the universality of this phenomenon. For example, in the studies of Humphries et al. (2001), it was the −174C allele carriers that had the increased risk of higher systolic blood pressure and coronary heart disease. In addition, it was the −174C allele carriers that produced in vivo the highest amount of IL-6 and had less favorable outcomes of coronary revascularization surgery than the GG homozygotes (Bittar et al., 2005).
We hypothesized that variation in the promoter region of the IL-6 gene, if of functional consequence, would influence IL-6 gene transcription and thus affect plasma or tissue levels of IL-6, which would in turn influence plasma levels of key coronary heart disease risk factors and thus coronary heart disease risk itself.
However, the relationship between IL-6 genotypes and plasma levels of IL-6 appears to be complex. Several studies have suggested that a common variant (−174G/C) located in the promoter of the IL-6 gene could play a significant role in regulating the level of circulating and locally produced IL-6 (Liu et al., 2006). In the first report of 102 healthy subjects, the −174CC group had the lowest levels (Fishman et al., 1998); the Fishman's study showed that the −174 G/C polymorphisms in the IL-6 gene could increase the plasma concentrations of IL-6, but in subsequent large studies, the −174C allele has been associated with higher levels of IL-6 (Brull et al., 2001; Jones et al., 2001). Several large studies (Bennet et al., 2003; Qi et al., 2006; van Oijen et al., 2006; Danesh et al., 2008), with the largest (Herbert et al., 2006), including 1,526 Caucasians, did not find evidence for association. However, other studies with the largest Cardiovascular Health Study (CHS) with 4,714 elderly Caucasians (72 years median age) and a high prevalence of T2DM (28%) showed borderline significantly higher IL-6 levels for CC-genotype subjects compared to GG (p=0.04) (Walston et al., 2007).
These studies showed conflicting results regarding the effect of −174G/C IL-6 polymorphism on plasma IL-6 levels and circulating IL-6 levels. To help clarify the inconsistent findings, we conducted a meta-analysis of the published genetic association studies of the −174 G/C polymorphisms in the IL-6 gene and the circulating IL-6 levels in the normal population.
Method
Publication search
This review was performed according to the Quality of Reporting of Meta-Analysis (QUORUM) guidelines (Moher et al., 1999). Three online electronic databases (PubMed, Embase, Web of Science) were searched using the terms: “IL-6” or “IL 6” or “IL6” or “interleukin6” or “interleukin 6” or “interleukin-6” paired with “174C,” “174G,” “rs1800795,” respectively (last search was updated on 30 April, 2013). In addition, the references of the candidate articles were examined in an effort to identify eligible studies that may not have been identified by the initial search. Abstracts or unpublished reports were not included and only the studies published in the English language were considered in the present meta-analysis.
Inclusion and exclusion criteria
For inclusion in the present meta-analysis, the identified articles had to provided all the following criteria: (1) studies on the IL-6 −174G/C polymorphism of a normal population (people without disease, especially disease related to inflammation and obviously influenced the IL-6 levels, such as coronary heart disease, diabetes mellitus, hypertension, nephropathy, obesity, metabolic syndrome, cancer, pregnancy, Alzheimer's disease, and stroke); (2) published case–control studies; (3) cohort studies; (4) studies published as full-length articles in English; (5) studies with data for −174 genotype in the IL-6 gene promoter; and (6) studies with sufficient data for estimating the mean differences of circulating IL-6 or with 95% confidence interval (CI). The exclusion criteria included (1) no control population; (2) no usable data reported; and (3) duplicates.
Data extraction
Two investigators independently extracted data (Huang and Wang) and discrepancies were resolved by group discussion. For each study, the following information was extracted: name of the first author, year of publication, ethnic origin of the studied population, number in −174 genotype groups, male percentage, mean ages in −174 genotype groups, genotyping method, and the IL-6 assay method. The studies' baseline and exclusion criteria were inspected to be sure they were adapted to our definition of a normal population. If multiple published reports from the same study population were available, we included only the one with the largest sample size and complete data.
Statistical analysis
The mean differences were pooled through a random effects model, using the inverse variance approach when heterogeneity existed among these studies. Otherwise, a fixed effects model was adopted. Heterogeneity between studies was assessed by the Q-test and I 2 statistic, where p<0.10 and I 2>30% indicated evidence of heterogeneity. For −174G/C polymorphism, the association between the allele and the circulating IL-6 levels, as well as homozygous comparison (GG vs. CC), heterozygous comparison (GC vs. GG or CC), dominant genetic model (CC+GC vs. GG), and recessive genetic model (CC vs. GG+GC) were explored. Forest plots were used to demonstrate the differences between studies and to provide an estimate of the overall result. Sensitivity analysis (one-study removed test) was performed to assess the impact of each study on the combined effect of the present meta-analysis. The single given study is excluded, when it is omitted in the sensitivity analysis and the estimate value is outside the pooled 95% CI. The Egger's and Begg's graphical methods were used to provide the diagnosis of the potential publication bias (Egger et al., 1997). All statistical analyses were performed using Stata software (version 12.0; Stata Corporation, College Station, TX) and RevMan software (version 5.2; Cochrane Collaboration). All p values were for a two-sided analysis and values of p<0. 05 were considered statistically significant.
Result
Characteristics of studies
The study selection process is detailed in Figure 1. Based on our preliminary search criteria, a total of 19 publications were eligible. Among these articles, thirteen publications were homozygous (CC vs. GG) or heterozygous (GC vs. GG or CC) comparisons, five publications were dominant genetic model (CC+GC vs. GG), and only one publication was recessive genetic model (CC vs. GG+GC). However, only 18 studies were included as the recessive genetic model study (only one publication was excluded in the final meta-analysis), comprising in total of 9432 homozygous subjects, 8670 heterozygous subjects, and 6528 subjects with dominant genetic model. All studies were case control in design. Table 1 shows the main characteristics of these studies. For homozygous comparison, 13 studies were available (all Caucasian), including 5971 subjects with GG genotype and 3461subjects with CC genotype. Three studies considered that the −174CC group had the lowest IL-6 levels, but nine studies suggested there was no association between the IL-6−174G/C polymorphism and IL-6 levels. Furthermore, one study found an association of the IL-6 −174G/C polymorphism and an increased level of IL-6. For the dominant genetic model (CC+GC vs. GG), five studies were available (four Caucasian, one North Indian), including 6528 subjects with CC/GC genotype and 3719 subjects with GG genotype.
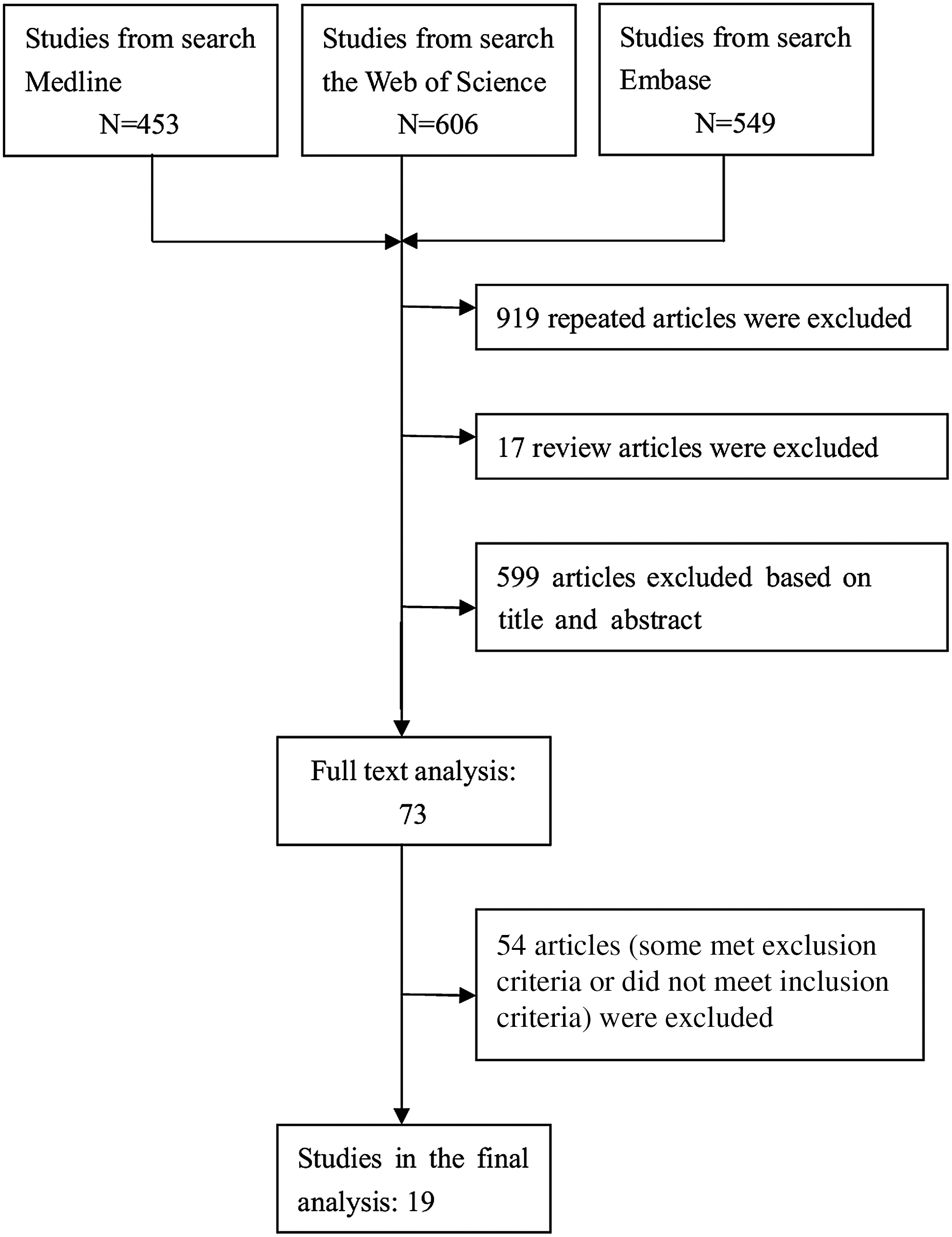
Flow diagram of the article selection process for Interleukin-6 (IL-6) −174G/C polymorphism and circulating IL-6 levels.
NA, not available; RFLP, restriction fragment length polymorphism; PCR, polymerase chain reaction; DASH, dynamic allele-specific hybridization; SSCP, single-strand conformation polymorphism; ELISA, enzyme-linked immunosorbent assay.
Meta-analysis
No significant association of IL-6 −174G/C polymorphism and circulating IL-6 levels in a normal population was observed (Figs. 2 –4 and Table 2). However, heterogeneity was observed among the studies when compared among the GG, GC, and CC genotypes (Table 2). Sensitivity analysis revealed that, for IL-6 −174 G/C polymorphism, the independent study by Shen et al. influenced the heterogeneity in the homozygous and heterozygous comparison (Figs. 5 –7). After Shen et al.'s study was excluded, the remaining studies' heterogeneity became acceptable (Table 2). When all 12 studies were pooled into the meta-analysis, there was no evidence of an association between IL-6 −174G/C polymorphism and circulating IL-6 levels; for homozygous comparison (GG vs. CC): the pooled standard mean difference (SMD) was −0.01, 95%CI: −0.1–0.08, I 2=63.6%, p=0.001(Fig. 8); for heterozygous comparison (GC vs. GG or CC), the pooled SMD (GG vs. GC) was −0.05, 95%CI: −0.11–0.01, I2=47.6%, p=0.024 (Fig. 9) and the pooled SMD (CC vs. GC) was 0.03, 95%CI: −0.03–0.1, I 2=43.6%, p=0.041 (Fig. 10). Under dominant model, five eligible studies had been identified: the pooled SMD was −0.05, 95%CI: −0.11–0.01, I 2=33.2%, p=0.175 (Fig. 11), there was no association between IL-6 −174G/C polymorphism and circulating IL-6 levels. Under recessive genetic model, only one eligible study had been identified, and therefore it was excluded from the analysis.

Meta-analysis of studies of the IL-6 −174G/C polymorphism for homozygous comparison (GG vs. CC).
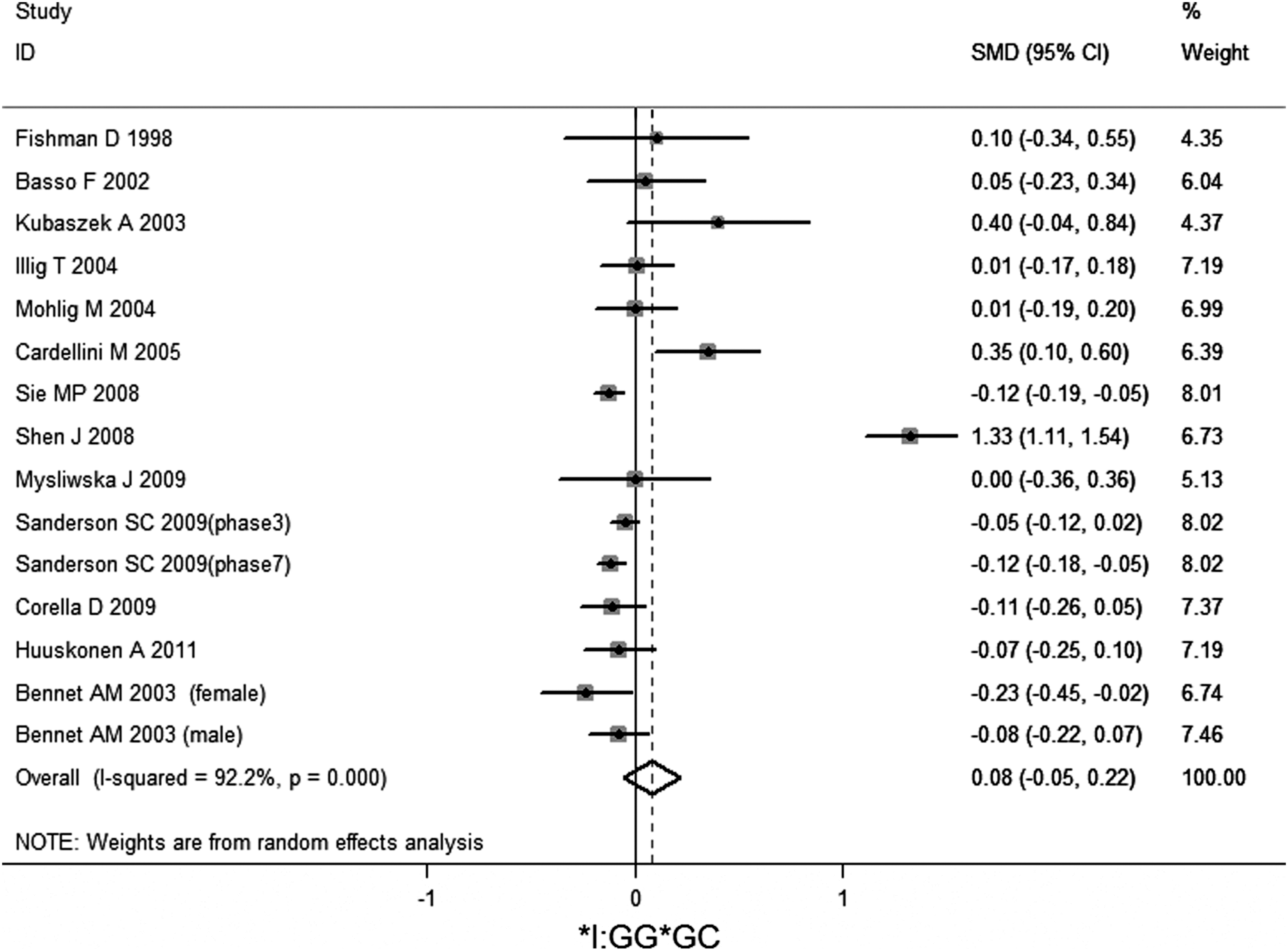
Meta-analysis of studies of the IL-6 −174G/C polymorphism for heterozygous comparison (GG vs. GC).
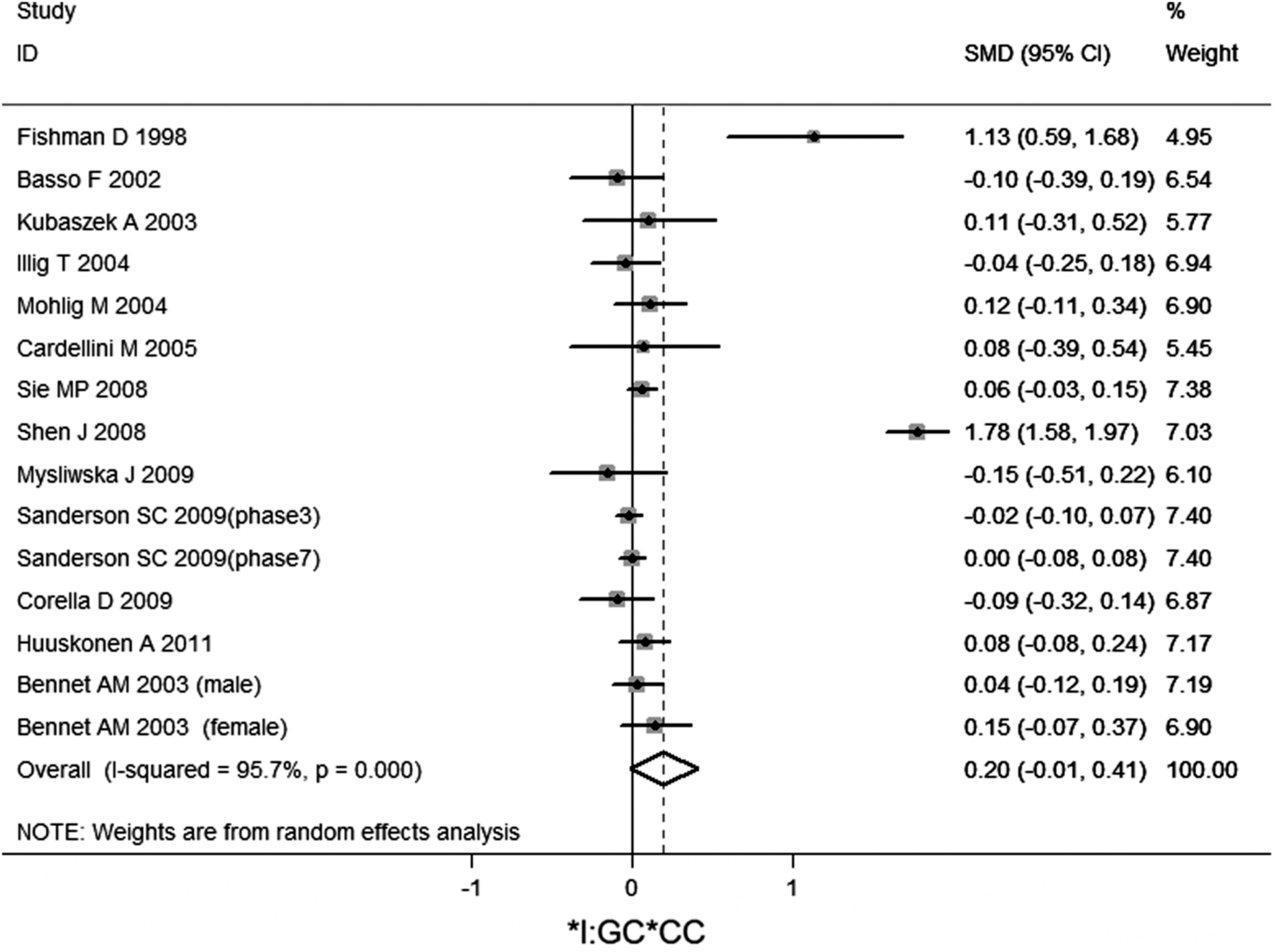
Meta-analysis of studies of the IL-6 −174G/C polymorphism for heterozygous comparison (GC vs. CC).
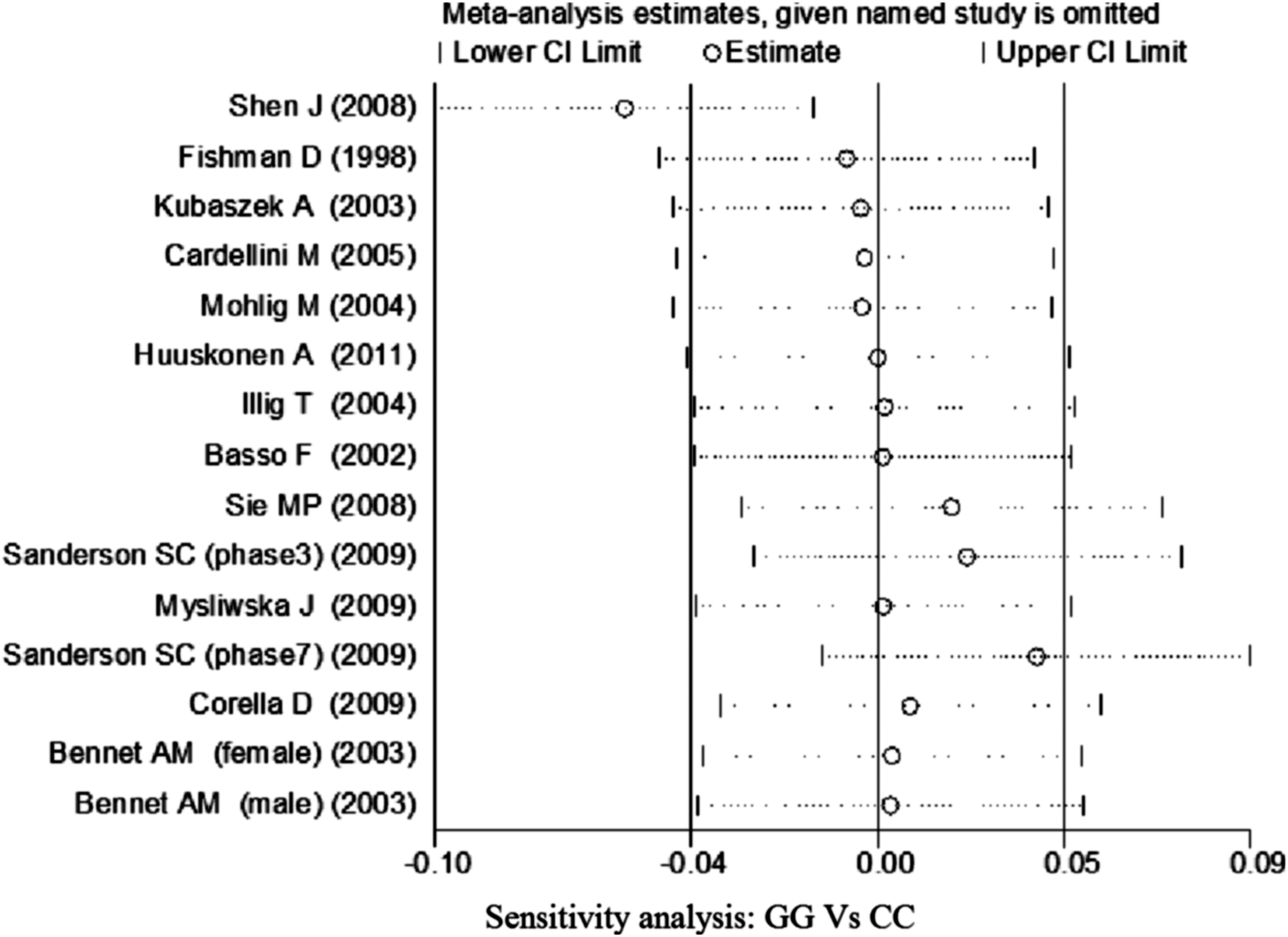
Sensitivity analysis of studies of the IL-6 −174G/C polymorphism for homozygous comparison (GG vs. CC).
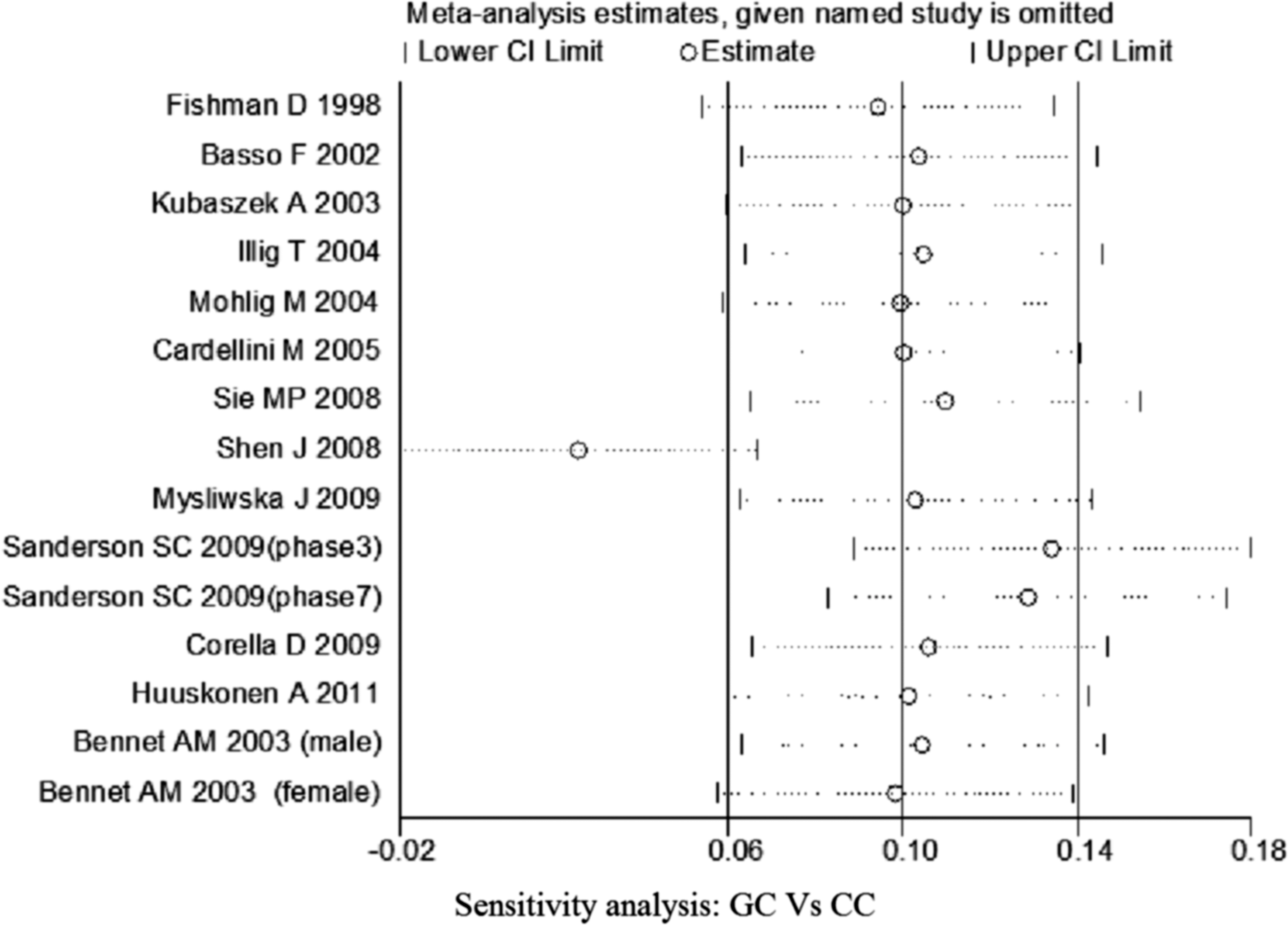
Sensitivity analysis of studies of IL-6 −174G/C polymorphism for heterozygous comparison (GC vs. CC).

Sensitivity analysis of studies of IL-6 −174G/C polymorphism for heterozygous comparison (GG vs. GC).
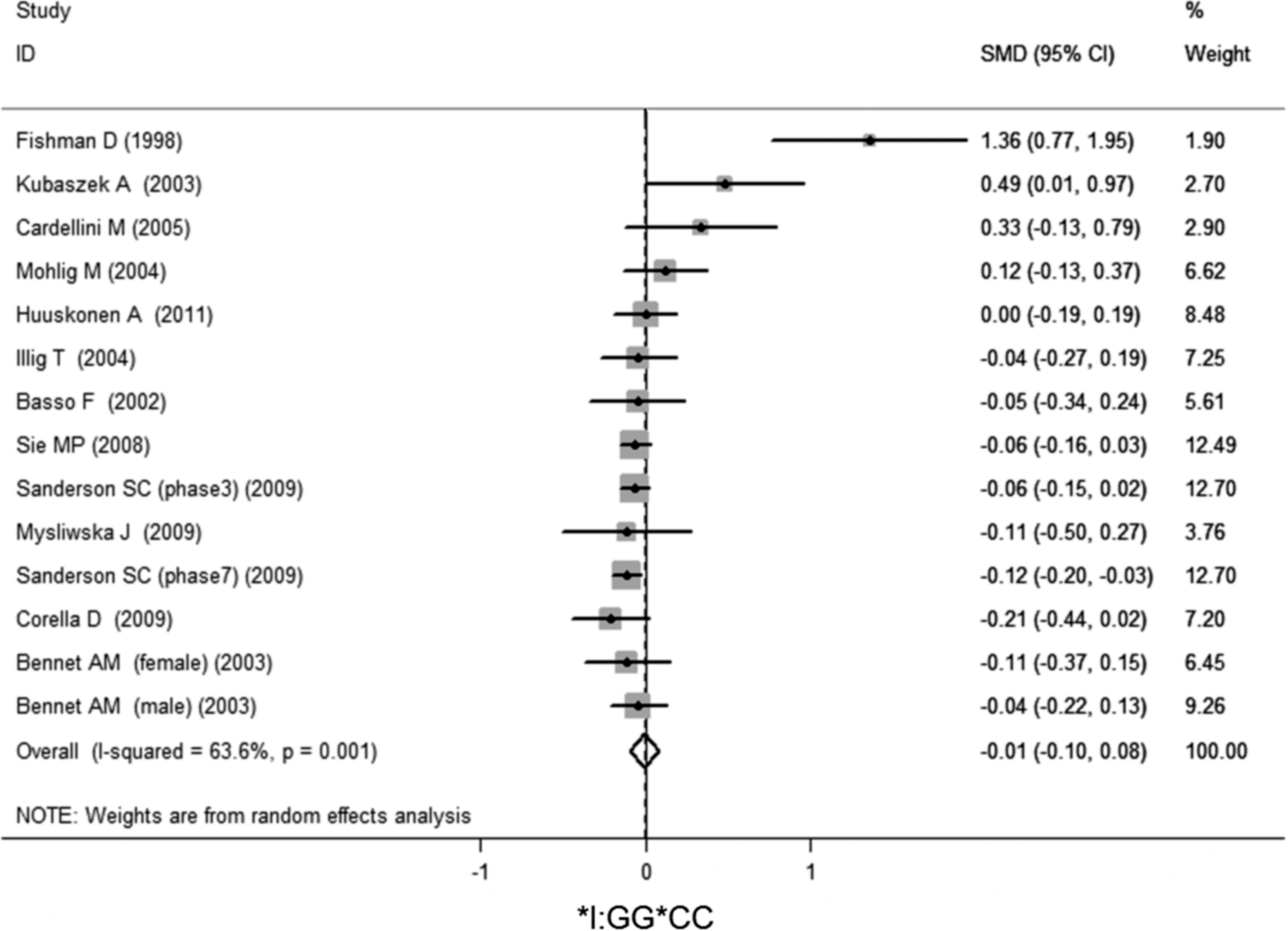
Meta-analysis of studies of the IL-6 −174G/C polymorphism for homozygous comparison (GG vs. CC) after Shen et al.'s study was excluded.
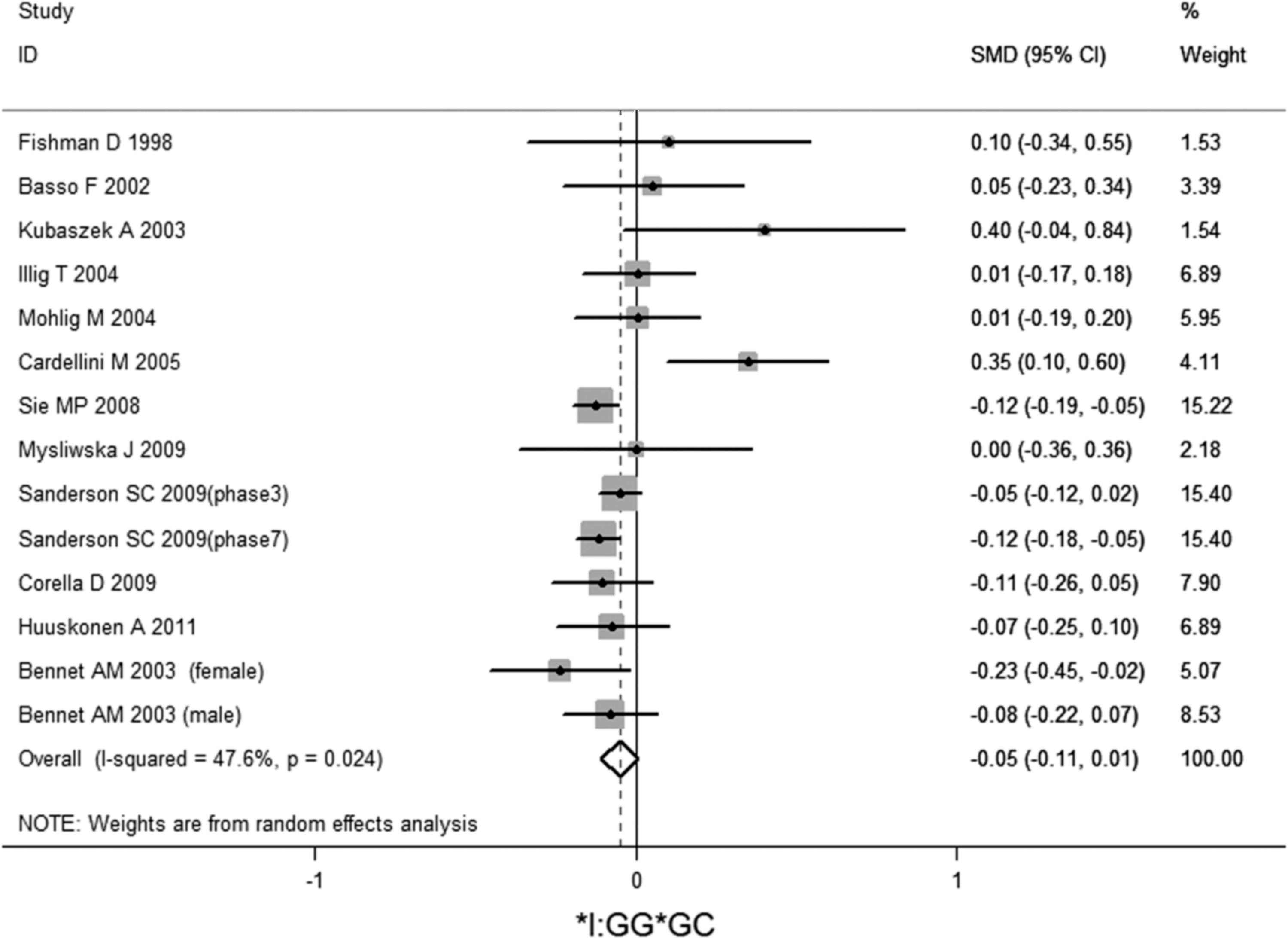
Meta-analysis of studies of the IL-6 −174G/C polymorphism for heterozygous comparison (GG vs. GC) after Shen et al.'s study was excluded.
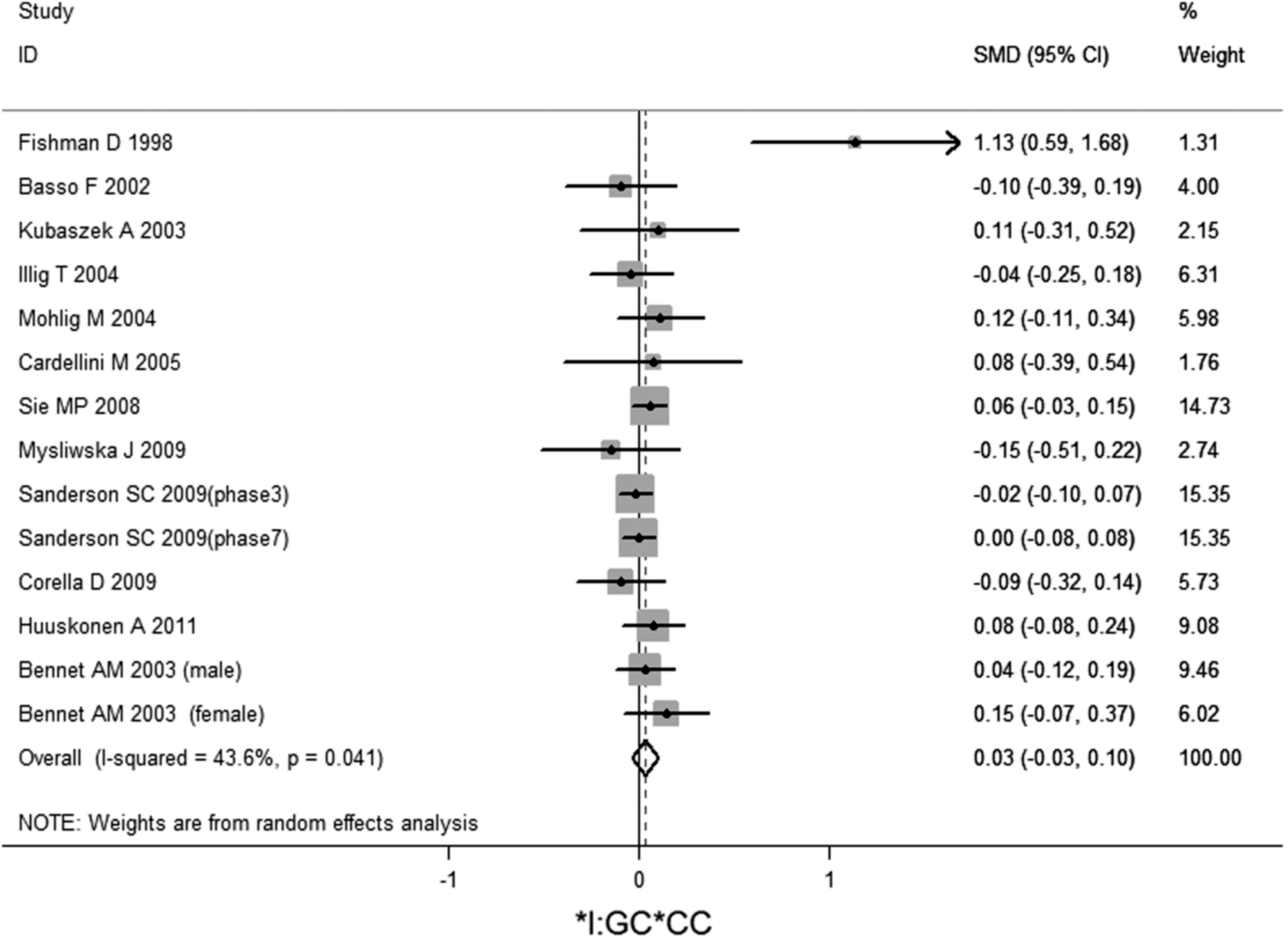
Meta-analysis of studies of the IL-6 −174G/C polymorphism for heterozygous comparison (GC vs. CC) after Shen et al.'s study was excluded.
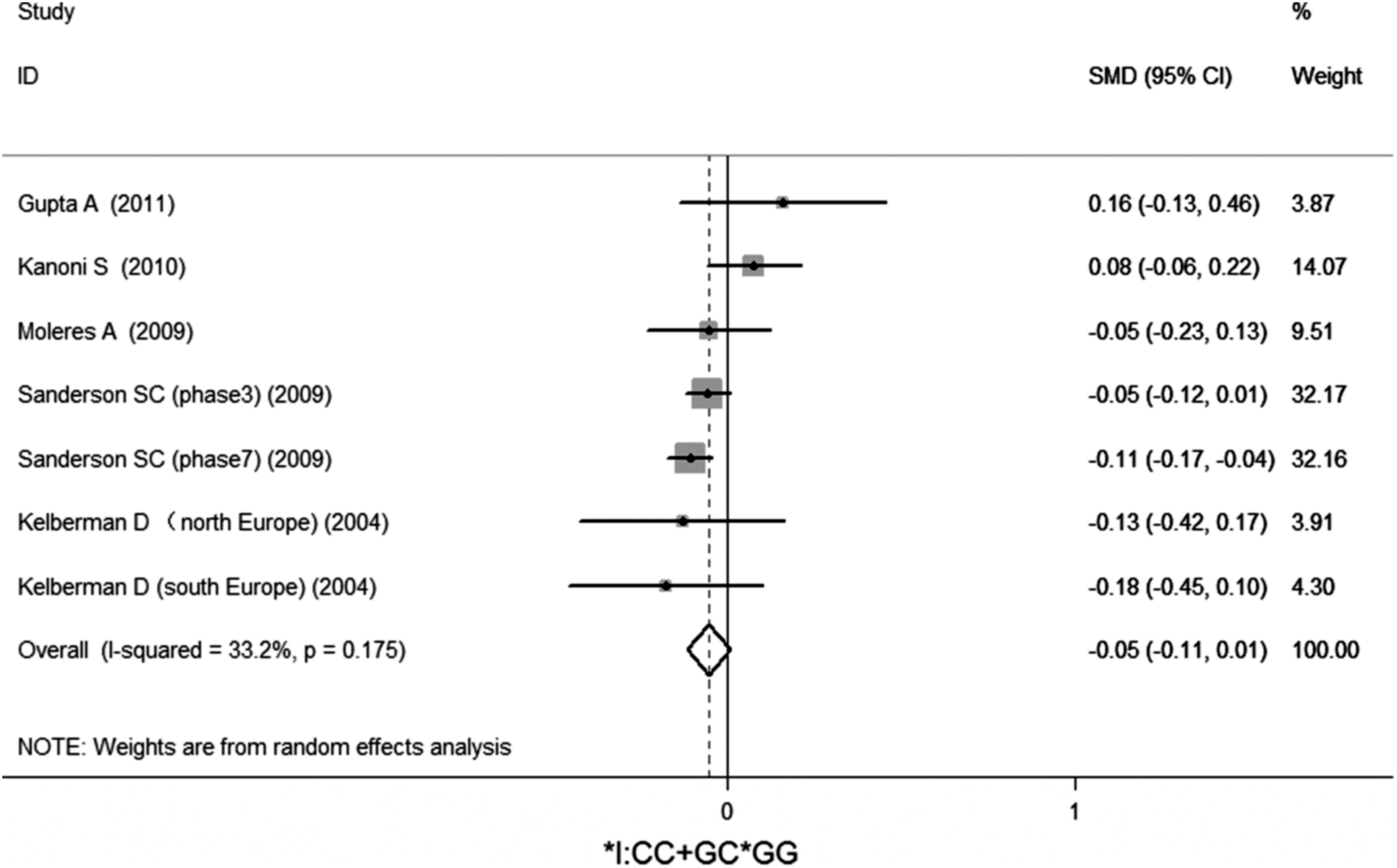
Meta-analysis of studies of the IL-6 −174G/C polymorphism in the dominant genetic model (GC+CC vs. GG).
For the studied IL-6 promoter polymorphisms, the visual examinations of the shape of funnel plots did not reveal any evidence of obvious asymmetry, suggesting the absence of publication bias. Subsequently, the Egger's test was used to provide statistical evidence of funnel plot symmetry and, as expected, the results showed no evidence of publication bias (Table 2).
Discussion
In this meta-analysis of a large normal population, we have demonstrated that the presence of the minor G allele at the IL-6 −174G/C single-nucleotide polymorphism (SNP), reportedly responsible for higher IL-6 expression, was not significantly associated with circulating IL-6 levels. However, because of the high heterogeneity, we were not able to determine a definitive conclusion. Therefore, we conducted a sensitivity analysis to identify the source of the heterogeneity, and the independent study by Shen et al. influenced the heterogeneity in the homozygous and heterozygous comparison. We found their participants were all with higher fasting plasma total TG, VLDL, and large VLDL. It has been suggested that increased adiposity may lead to higher IL-6 levels and worse insulin resistance (Fernandez-Real et al., 2000). It is likely that the known relationship between inflammation, obesity, and insulin resistance in humans is involved (Donath and Shoelson, 2011). Obesity is associated with a proinflammatory state (Madec et al., 2010), and maybe such a proinflammatory state influenced the heterogeneity in our meta-analysis.
Many studies have demonstrated associations of IL-6 −174G/C polymorphism with obesity, insulin resistance, dyslipidemia, and cardiovascular disease (CVD) (Fernandez-Real et al., 2000; Humphries et al., 2001; Stephens et al., 2004), and both in vitro and in vivo studies have indicated that the −174G/C polymorphism alters the IL-6 gene transcription rate and protein expression (Terry et al., 2000).
However, some subsequent articles did not confirm the association with IL-6 levels in healthy subjects, but many did see this in patients undergoing an inflammatory stimulus such as after coronary artery bypass surgery (CABG) (Brull et al., 2001). Five to seven days after CABG, the mean IL-6 levels rose sixfold in the whole cohort (p<0.0001) compared with baseline; the magnitude of the rise was also IL-6 genotype dependent: patients carrying −174C allele had IL-6 levels significantly higher than those of −174GG genotype (Wypasek et al., 2010). Kuo et al. (2008) showed that in patients with stable coronary artery disease (CAD), IL-6 and CRP were independently related to the presence of complex coronary plaque; however, the IL-6 genotypes were not related to serum IL-6 levels. A higher frequency of −174G/C IL-6 polymorphism together with increased IL-6 levels was observed in patients with extensive CAD (Mysliwska et al., 2006). In the Wypasek et al. (2010) study, neither procedure-related data during CABG nor major adverse cardiovascular events during the perioperative period were associated with −174G/C IL-6 polymorphism, but the intubation time or duration of intensive care was positively correlated with the magnitude of a postoperative increase in IL-6 levels.
This could be explained by the fact that it is the inflammatory processes, which influenced circulating IL-6 levels; the effects of −174G/C IL-6 polymorphism were modified by inflammation, but were not significantly associated with normal populations without inflammation.
The key question is whether there is a genetic predisposition in the disease under investigation to produce higher IL-6 levels, leading to a chronic proinflammatory state. Because IL-6 is involved in a multitude of inflammatory processes, it is possible that this SNP is directly related to other genes and/or other inflammatory pathways. In many cases, the increase in IL-6 levels is believed to be secondary to other inflammatory mediators such as IL-1 and TNF (Woo and Humphries, 2013). Our meta-analysis gave us a clue that the influence of IL-6 −174G/C polymorphism to IL-6 levels is minimal or it is not the IL-6 −174G/C polymorphism that directly influenced IL-6 levels; otherwise, they can be detected by our study.
In addition, circulating IL-6 may be less important than local IL-6 expression as studies have shown that locally produced cytokines possess important auto-/paracrine properties that influence the diverse functions of other tissues (Fried et al., 1998).
In addition, a polymorphism in the promoter sequence of the IL-6 gene (rs1800795; IL6 −174G/C) has been found to modulate individual sensitivity to the effects of threatening or adverse environments in activating IL-6 transcription (Cole et al., 2010). In people bearing the ancestral G allele of the polymorphism, adversity-induced activation of the GATA1 transcription factor leads to increased IL-6 gene transcription, and those individuals show increased risk of inflammation-related disease and mortality when confronted by extended periods of significant life adversity (Cole et al., 2010). In contrast, individuals bearing the C allele of the polymorphism are protected from such effects, because the GATA1 transcription factor cannot efficiently bind to the IL-6–174 locus after SNS-induced activation (Cole et al., 2010).
Furthermore, a single point measurement of plasma IL-6 concentrations may not adequately reflect an individual's long-term exposure of chronic inflammation. It is unknown whether average, peak, or inflammatory levels of IL-6 are relating to −174G/C polymorphism; and fasting IL-6 measurements may not be appropriate for examining the identified genotype–phenotype relationship (Herbert et al., 2006). The accurate measurement of IL-6 in serum is complicated by the fact that IL-6 levels vary in a diurnal manner in the healthy individual, so the time of sampling would affect the final results (Woo and Humphries, 2013). Unfortunately, none of the articles describing serum IL-6 levels provide data on the time of collection or time to storage and length of storage.
Several potential limitations of this meta-analysis should be considered. First, this meta-analysis only focused on articles published in English. Our data were derived mostly from subjects of Caucasian descent; therefore, it may be that results do not apply to cases from other ethnicities. Second, not all the control subjects were age and sex matched to cases, which was likely one of the causes for heterogeneity. Third, because the included studies are observational and confounding, biases might have affected the pooled estimates. Furthermore, large and well-designed studies are necessary to determine the relationship of this genetic polymorphism with IL-6 expression in a normal population. However, this is the first meta-analysis, based on a normal population, about the relationship between IL-6 −174G/C polymorphism and circulating IL-6 levels. The potential sources of heterogeneity in the meta-analysis were assessed.
In conclusion, the meta-analysis provides evidence that the −174G/C polymorphism in the IL-6 gene is not significantly associated with circulating IL-6 levels in a normal population.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation (No.81270127), China.
Disclosure Statement
No competing financial interests exist.