
Abstract
Many nonsynonymous single-nucleotide polymorphisms (SNPs) in the human deoxyribonuclease I-like 1 (DNase 1L1) gene, possibly implicated in the blocking of endocytosis-mediated foreign gene transfer, have been identified, but only limited population data are available and no studies have evaluated whether such SNPs are functional. Genotyping of all 21 nonsynonymous human DNase 1L1 SNPs was performed in 16 different populations representing three ethnic groups using the PCR-restriction fragment length polymorphism technique. All of the nonsynonymous SNPs, except for SNP p.Val122Ile in Caucasian populations, exhibited a monoallelic distribution in all of the populations. On the basis of alterations in the activity levels resulting from the corresponding amino acid substitutions, two activity-abolishing and four activity-reducing SNPs were confirmed to be functional. Although all of the nonsynonymous SNPs that affected the catalytic activity showed extremely low genetic heterogeneity, it seems plausible that a minor allele of six SNPs producing a loss-of-function or extremely low-activity variant could serve directly as a genetic risk factor for diseases. Especially, the amino acid residues in activity-abolishing SNPs were conserved in animal DNases 1L1. Furthermore, results of phylogenetic analysis suggest that DNase 1L1 might have appeared latest among the DNase I family during the course of molecular evolution.
Introduction
D
Among the SNPs in DNASE1L1, we have focused on nonsynonymous SNPs potentially affecting the DNase activity through the corresponding amino acid substitution as a functional SNP; three nonsynonymous SNPs, p.Val122Ile (rs34952165), p.Gln170His (rs6643670), and p.Asp227Ala (rs5987256), have been examined in previous studies (Fujihara et al., 2010; Ueki et al., 2010a). Since then, a total of 18 nonsynonymous SNPs in the gene have been registered in the NCBI dbSNP database (
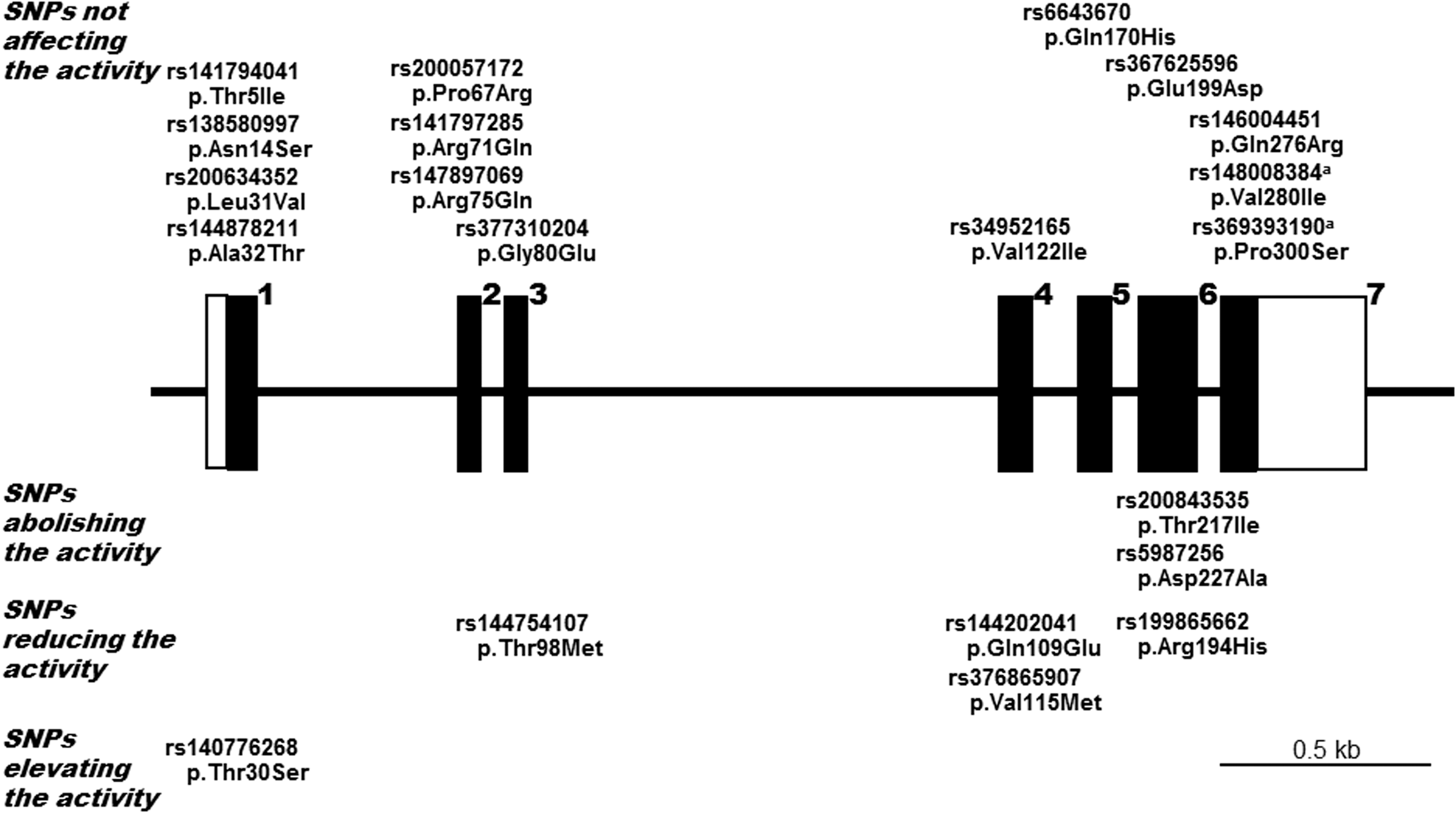
Schematic representation of the genomic structure of the human DNASE1L1 and the positions of the single-nucleotide polymorphisms (SNPs) examined. Each ID number in the NCBI database is shown. Exons are shown by solid boxes. According to alterations in the levels of the DNase 1L1 activity, 19 nonsynonymous SNPs were classified into 14 SNPs not affecting the activity, 2 abolishing the activity, 4 reducing the activity, and 1 elevating the activity; each of the SNPs in these four groups are shown at the sites of the exons where they are located. aThe effect of the amino acid substitution derived from SNPs p.Val280Ile and p.Pro300Ser, on the activity was not examined.
In the present study, we investigated the genotype distribution of the 18 nonsynonymous SNPs in DNASE1L1 in 16 different populations worldwide derived from three ethnic groups, using a novel genotyping method for each SNP, and the effects of all these SNPs on the enzymatic activity, to evaluate the genetic heterogeneity and functionality of each SNP. Furthermore, multiple alignment analysis of the amino acid sequences of animal DNase 1L1, including phylogenetic analysis, was performed to examine the role of each SNP-related amino acid residue in the protein.
Materials and Methods
Samples from subjects
Genomic DNA was extracted using a QIAamp DNA Mini Kit (Qiagen, Chatsworth, CA) from blood or bloodstain samples randomly collected from healthy subjects (n=1752) derived from 16 different populations. The Asian populations included 110 Japanese (Shimane Prefecture), 352 Koreans (Busan, South Korea), 193 South Chinese (Shenyang and Guangzhou of China), 112 Mongolians (Ulaanbaatar, Mongol), 153 Tibetans (Katmandu of Nepal), 35 Sri Lanka Tamils (Kandy of Sri Lanka), 48 Sri Lanka Sinhalese (Kandy of Sri Lanka), and 40 Tamangs (Kotyang of Nepal); the Caucasian population included 136 Turks (Adana area, Southern Turkey), 68 Germans (Munich), and 199 Mexicans (60 Mestizos, 88 Nahuas, and 51 Huicholes); the African populations included 126 Ovambos (Bantusin, Namibia), 105 Ghanaians (Ghana), and 75 Xhosas (Cape Town, South Africa). Some of the samples were the same as those used for previous studies (Fujihara et al., 2010; Ueki et al., 2010a). Written informed consent was obtained from each participant. The study was approved by the Human Ethical Committees of the institute.
Construction of expression vectors encoding human DNase 1L1 and its variant forms corresponding to each SNP
Expression pcDNA3.1 (+) vectors (Invitrogen, San Diego, CA) inserted with the entire coding sequence of human DNase 1L1 cDNA was prepared from the total RNA of human skeleton muscle (Clontech, Palo Alta, CA) by reverse transcriptase–PCR amplification using an Expanded High Fidelity PCR System (Roche Diagnosis, Mannheim, Germany) with a set of two primers, 5′-GAATTCCAGCCACACAGCCATGCACTACC-3′ (sense) and 5′-CTCGAGCGCTCAGGCAGCAGGGCACAG-3′ (antisense), corresponding to the sequences of cDNA from positions −13–10 and 900–932, respectively; inserted cDNAs was derived from the predominant haplotype of the SNPs in Japanese. Using this construct as a template, the DNase 1L1 ΔC construct, which was devoid of the nucleotide region corresponding to the C-terminal domain, including Ala277–Ala302, used for the activity assay was prepared using the KOD-Plus Mutagenesis kit (Toyobo Co., Ltd., Osaka, Japan) and used as a wild type. Nineteen amino acid-substituted constructs corresponding to each SNP were prepared by site-directed mutagenesis using a KOD-Plus Mutagenesis Kit with the wild-type construct as a template. In these constructs, the amino acid residue was replaced by the counterpart derived from a minor allele in each SNP; for example, the T5I construct of DNase 1L1, in which a Thr residue at position 5 in the protein is replaced by Ile derived from the minor allele, corresponds to SNP p.Thr5Ile. Furthermore, the five other amino acid-substituted constructs related to the SNPs were prepared in the same manner. Nucleotide sequences of all the constructs were confirmed by DNA sequence analysis. Purification of each construct used for transfection was performed using the Plasmid Midi kit (Qiagen).
Transient expression of the expression vectors and assay for the DNase 1L1 activity
COS-7 cells were maintained in Dulbecco's modified Eagle's medium containing 1 mM
Genotyping of nonsynonymous SNPs in DNASE1L1 by the PCR–restriction fragment length polymorphism method
Genotyping assays for each of 18 nonsynonymous SNPs in DNASE1L1 were separately performed using PCR followed by restriction fragment length polymorphism (RFLP) analysis (Yasuda et al., 1995), in the same manner as the other SNPs (Ueki et al., 2010a). Primers for the specific amplification of the DNA fragments encompassing a substitution site corresponding to each SNP were designed on the basis of the nucleotide sequences of the human DNASE1L1 (NCBI Reference Sequence: NG 012884.1) (Supplementary Table S1; Supplementary Data are available online at
PCR amplification was performed in a 25 μL reaction mixture using ∼5 ng of DNA. The reaction mixture contained a 1×buffer (15 mM Tris-HCl, pH 8.0, 50 mM KCl), 1.5 mM MgCl2, 0.5 μM of each primer, 200 μM dNTPs, and 1.25 U of Taq polymerase (AmpliTaq Gold; Applied Biosystems, Foster City, CA). PCR was performed with a protocol consisting of initial denaturation at 94°C for 7 min, followed by 30 cycles with denaturation at 94°C for 30 s, annealing at 55°C–65°C (Supplementary Table 1) for 30 s, and extension at 72°C for 1 min, followed by a final extension at 72°C for 7 min. Two microliters of the PCR product obtained using each pair of primers was digested with 5 U each enzyme listed in Supplementary Table 1 (New England Biolabs, Ipswich, MA) at 37°C for 3 h in a final reaction mixture volume of 15 μL to determine the genotype of each SNP. The digested products (5 μL) were separated in an 8% polyacrylamide gel, and the patterns on the gels were visualized by silver or ethidium bromide staining, as described previously (Yasuda et al., 2010).
Direct sequencing of the genomic DNA region including substitution site of each SNP in the DNase 1L1 gene for the representative subjects was performed by the dideoxy chain-terminating method with the BigDye®Terminator Cycle Sequencing kit (Applied Biosystems). The sequencing run was performed on a Genetic Analyzer 310 (Applied Biosystems).
Analytical methods
Each Ortholog of human DNases 1L1 and 1L2 in other available animal species were surveyed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (
DNase 1L1 activity was compared between wild-type and variant enzymes by means of the unpaired, Student's t-test. Differences at p<0.05 were considered to be statistically significant.
Results
Effect of amino acid substitution resulting from each nonsynonymous SNP in DNASE1L1 on the expression of DNase activity
We have previously developed the SRED method for quantification of endonuclease activity, including DNase I, utilizing the radial diffusion of the enzyme in a thin agarose gel plate containing DNA as a substrate (Nadano et al., 1993; Yasuda et al., 1998). The SRED method allowed us to detect and quantify extremely low levels of the DNase activity. However, the SRED method can only be applied for assay of the DNase activity in a soluble state. Since DNase 1L1 is a GPI-anchored membrane DNase and its C-terminal hydrophobic domain serves as a signal sequence for GPI attachment (Shiokawa et al., 2007), it is not possible to measure the activity of intact DNase 1L1 in cells with sufficient sensitivity to clarify the effects of amino acid substitutions corresponding to individual nonsynonymous SNPs in DNASE1L1 on the activity. To overcome these problems, we constructed a DNase 1L1 ΔC form lacking the C-terminal hydrophobic domain, thus allowing the conversion of the GPI-anchored form of DNase 1L1 to a soluble form. This allowed us to quantify the DNase 1L1 with sufficient sensitivity using the conventional SRED method. An expression construct of DNase 1L1 ΔC containing the entire coding sequence of human DNase 1L1, but lacking the C-terminal hydrophobic domain, derived from the predominant haplotype of these SNPs was used as the wild type; 19 variant constructs of the DNase 1L1 protein derived from the minor allele in each SNP were prepared separately, transiently expressed in COS-7 cells, and the resulting DNase 1L1 activities in lysates derived from transfected cells (Table 1) and their conditioned medium (Fig. 2) were determined separately by the SRED method. Generally, alterations in the activity induced by amino acid substitution corresponding to each SNP in the lysates derived from the transfected cells were similar to those in the conditioned medium.
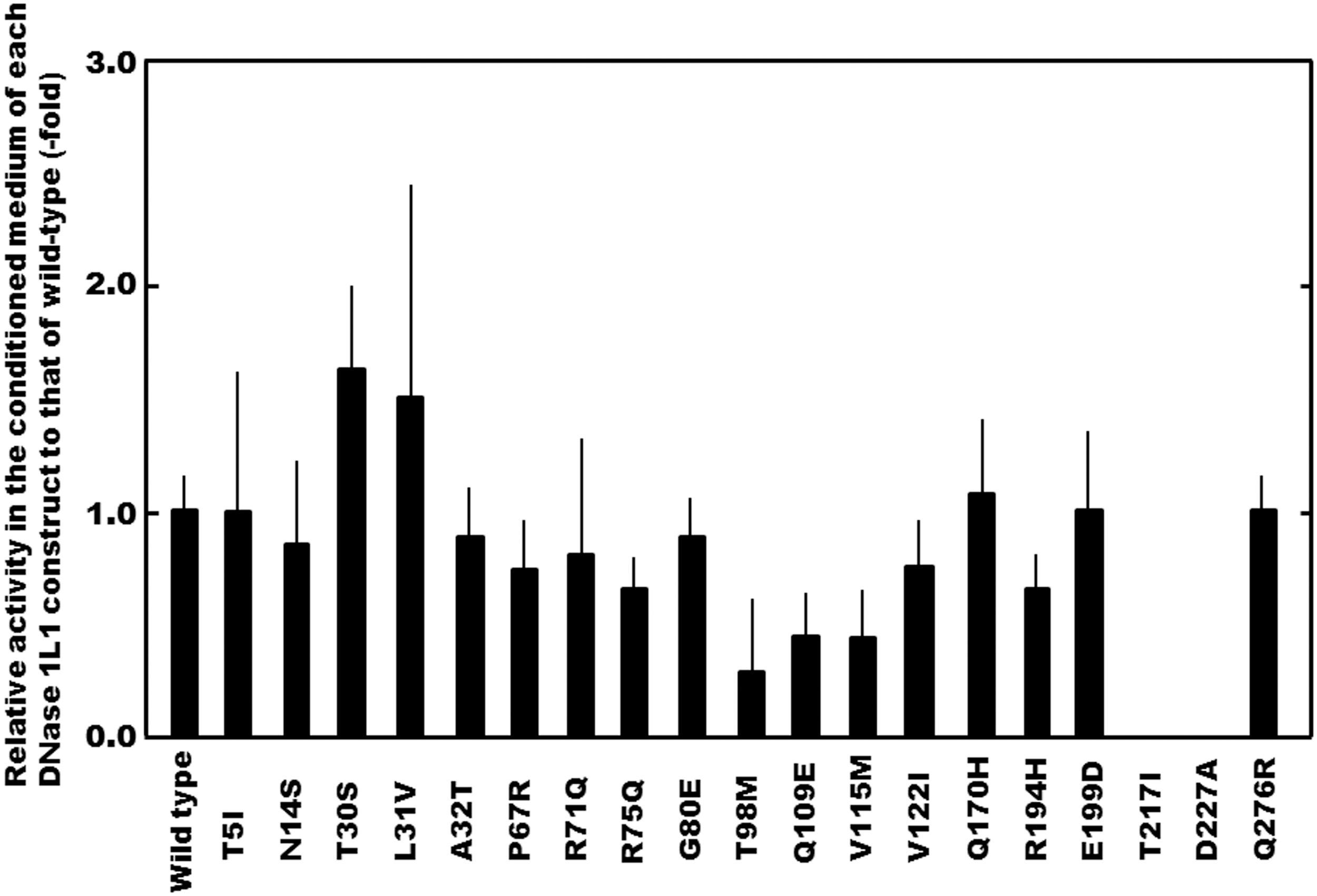
Relative activity of variant constructs corresponding to nonsynonymous SNPs to that of the wild-type DNase 1L1. The DNase 1L1 activity in the conditioned medium derived from the cells transfected with each construct was assayed by the single radial enzyme diffusion (SRED) method (Ueki et al., 2010a). The constructs, T217I and D227A, exhibited no DNase 1L1 activity detectable under our assay condition. The bars present SD (n=4).
Taken from the dbSNP.
Activity of each amino acid substituted DNase 1L1 is expressed as relative one to that of wild-type enzyme; the activity (n=4) of each construct in the lyzates derived from the transfected cells was compared with that of the wild-type enzyme (1.0±0.11).
Multiple alignment analysis of amino acid sequence was performed using 23 animal DNases 1L1 available in database. Number in parenthesis is the number of species in which the corresponding amino acid is substituted.
The expression construct DNase 1L1 ΔC lacking the C-terminal hydrophobic domain was used for the activity assay; since the amino acid residues corresponding to SNPs, pVal280Ile and p.Pro300Ser, were located in the C-terminal hydrophobic domain, the corresponding amino acid-substituted constructs could not be prepared, and therefore, the effect of these substitutions on the activity was not examined.
N.D., not determined; n.d., not detected; SNP, single-nucleotide polymorphism; n.e., not examined.
Among them, the levels of the DNase 1L1 activity derived from the T98M, Q109E, V115M, and R194H constructs were significantly lower than that of the wild-type enzyme. Substitution of Gln, Val, and Arg residues at positions 109, 115, and 194, respectively, in the DNase 1L1 protein obviously reduced the activity to ∼40%–50% of that of the wild-type enzyme, and furthermore, substitution of a Thr residue at position 98 reduced the activity greatly to ∼20%. It was noteworthy that the two constructs, T217I and D227A, exhibited no DNase 1L1 activity under our assay conditions. Therefore, among the SNPs affecting the DNase 1L1 activity, the activity was found to be abolished by amino acid substitutions resulting from p.Thr217Ile and p.Asp227Ala. On the other hand, when compared to the activity level of the wild type, the activity level of T30S was significantly high; substitution of a Thr residue at position 30 in the protein elevated the DNase 1L1 activity to about twofold that of the wild-type enzyme. However, the levels of the DNase 1L1 activity derived from the other amino acid-substituted constructs were similar to that of the wild type, indicating that substitution of Ile5, Asn14, Leu31, Ala32, Pro67, Arg71, Arg75, Gly80, Val122, Gln170, Glu199, and Gln276 residues in the DNase 1L1 protein had little effect on the activity. These findings permitted us to demonstrate that, among the amino acid residues related to nonsynonymous SNPs in the DNase 1L1 protein, each of the amino acid residue at positions 98, 109, 115, and 194 might be involved in expression of the enzyme activity, and that furthermore, those at positions 217 and 227 might be indispensable.
From these analyses of the variant forms corresponding to each of the nonsynonymous SNPs, we were able to divide these SNPs into four classes according to the observed alterations in the enzyme activity levels resulting from the corresponding amino acid substitutions: those not affecting the activity, those abolishing it, those reducing it, and those elevating it; among the 19 nonsynonymous SNPs examined in DNASE1L1, 12 did not affect the activity, 2 abolished it, 4 reduced it, and 1 elevated it (Fig. 1).
Multiple alignment analysis of the amino acid sequences of animal DNase1L1 and possible involvement of the amino acid residues corresponding to individual SNPs in expression of the enzyme activity
Multiple alignment analysis of the amino acid sequences of animal DNase 1L1 is useful for assessing the roles of individual SNP-related amino acid residues in the DNase 1L1 protein. Therefore, to survey orthologs of DNase 1L1, together with DNase 1L2, in Animalia organisms available in the KEGG database, a BLAST search of each genome database was performed using the amino acid sequence of human DNases 1L1 or DNase 1L2 as a query sequence. Orthologs of human DNase 1L1 with a low E-value were found in 23 organisms, demonstrating that DNase 1L1 is well distributed in vertebrates including not only mammals as reported previously (Shiokawa et al., 2005) but also birds, reptiles, amphibians, and fish (Fig. 3). Furthermore, lancelets (Florida lancelet), ascidians (sea squirt), and echinoderms (purple sea urchin) were demonstrated to possess each ortholog of human DNases 1L1 and 1L2. In addition, an ortholog of human DNase 1L2 was also found in a cnidarian (sea anemone). Recently, we demonstrated that an ortholog of human DNase 1L3 was present in a poriferan sponge (Amphimedon queenslandia), representing the lowest class in Metazoa, whereas the placozoan Trichoplax adhaerens, probably situated in a molecular–phylogenetic position intermediate between Porifera and Animalia, possessed both DNases I and 1L3 (Ueki et al., 2013b). On the other hand, an ortholog of human DNase 1L1 was found only in Deuterostomia but not in Porifera, placozoans and cnidarians; furthermore, similar to other DNases, DNase 1L1 was not distributed in any of the genome databases available for Protostomia, including insects and nematodes, among the Animalia, in contrast to another distinct type of DNase, DNase II, which has been identified even in non-metazoan organisms (MacLea et al., 2003). This phylogenetic distribution of the DNase I family in Animalia allows us to assume that DNase 1L1 might have appeared later in the course of molecular evolution of the DNase I family than did DNases I, 1L2, and 1L3; DNase 1L1 might have appeared at the phylogenetic level of the Deuterostomia.
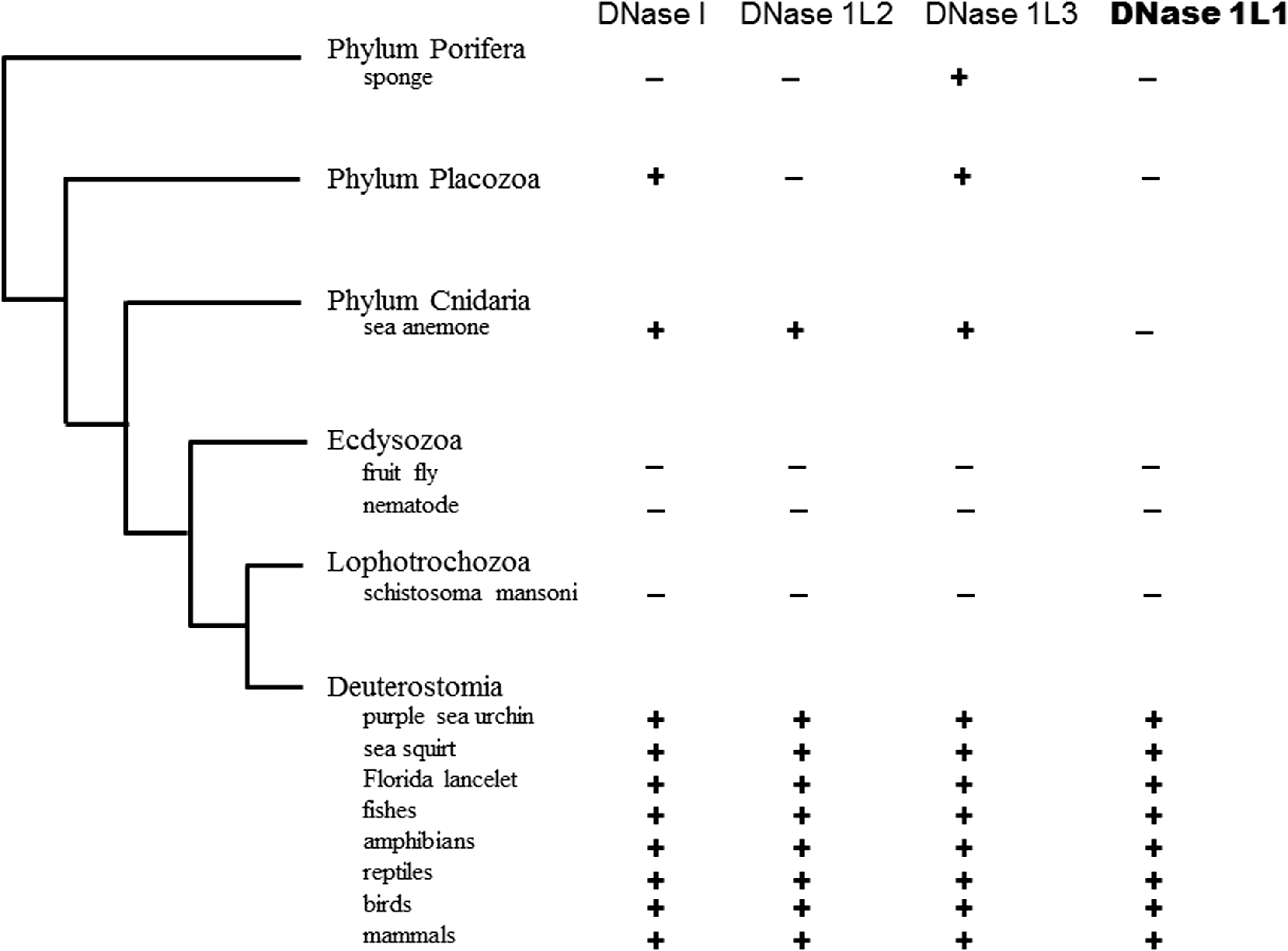
Molecular–phylogenetic distribution of DNases 1L1 and 1L2 in animals. A Basic Local Alignment Search Tool (BLAST) search was performed in each genome database available in the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (
We then investigated whether the amino acid residues corresponding to the various SNPs were well conserved in animal DNase 1L1 by multiple alignment analysis of the amino acid sequences of DNase 1L1 from Animalia organisms available on the genome database (Supplementary Fig. S1). The C-terminal hydrophobic domain serving as a signal sequence for the GPI attachment characteristic of mammal DNase 1L1 (Shiokawa et al., 2007) was deleted in the lower classes of Animalia, including Monotremata (platypus) and Marsupialia (opossum), suggesting that the localization of DNase 1L1 on the cell surface might have been acquired at the level of Marsupialia/Eutheria during the molecular evolution of animal DNase 1L1. There was no nonsynonymous SNP at the codon for the four amino acid residues crucially responsible for expression of the DNase activity, corresponding to His148, Thr222, Tyr226, and His265 in the DNase 1L1 protein (Shiokawa et al., 2005). However, in SNP p.Asp227Ala, an Asp residue at position 227 served as a general base in its catalysis is replaced by Ala; in fact, the minor allele giving rise to occurrence of an Ala residue in the DNase 1L1 protein produced a loss-of-function form of the enzyme. It is worth noting that the amino acid residues related to loss of the enzyme activity through amino acid substitution derived from p.Thr217Ile and p.Asp227Ala were completely conserved in animal DNase 1L1, whereas those corresponding to SNPs not affecting the activity were not conserved (Table 1), being compatible with our finding that all of the amino acid residues in activity-abolishing SNPs were completely or well conserved in animal DNase family proteins (Ueki et al., 2013a, 2013b).
To clarify the potential involvement of each amino acid residue corresponding to these SNPs in the expression of the enzyme activity, a series of amino acid-substituted constructs were examined (Fig. 4). Similar to substitution of the Thr217 residue by Ile (T217I) resulting from SNP. p.Thr217Ile, that by Ser (T217S) ablated the activity completely, even though it is a hydroxyl amino acid. Therefore, Thr itself located at position 217 in the protein, completely conserved in animal DNase 1L1, was found to be indispensable for the expression of the activity. Furthermore, replacement of Thr30 by a Val residue (T30V) abolished the DNase 1L1 activity, unlike the situation for the T30S construct, indicating that the hydroxyl group of the amino acid residue at position 30 in the protein, and not a hydrophobic residue, is required for expression of the enzyme activity. Since substitution of Thr30 by Ser elevated the DNase 1L1 activity, it seems plausible that a Ser residue is a more suitable hydroxyl amino acid than a Thr residue for expression of the DNase activity. With regard to SNPs reducing the activity, substitution of Thr98 by Ala (T98A) reduced the activity in the same manner as T98M, whereas that by Ser (T98S) did not greatly affect the activity. In other species, Lys or Gln residues are situated at position 98, in place of the Thr in human DNase 1L1. Also, substitution of Arg194 by Ala (R194A) reduced the activity more markedly than substitution by His (R194H). In this context, although these residues at position 194 are not conserved in animal DNases 1L1, substitution of several hydrophilic amino acids at the corresponding positions can all occur. Therefore, it seems plausible that at least a hydrophilic amino acid residue at positions 98 and 194 might be required for full expression of the enzyme activity. However, each amino acid residue corresponding to these SNPs reducing the activity was not necessarily well conserved in animal DNase 1L1, suggesting that the effect of amino acid substitutions resulting from these SNPs on the enzyme activity may be attributable to the properties of individual amino acids.
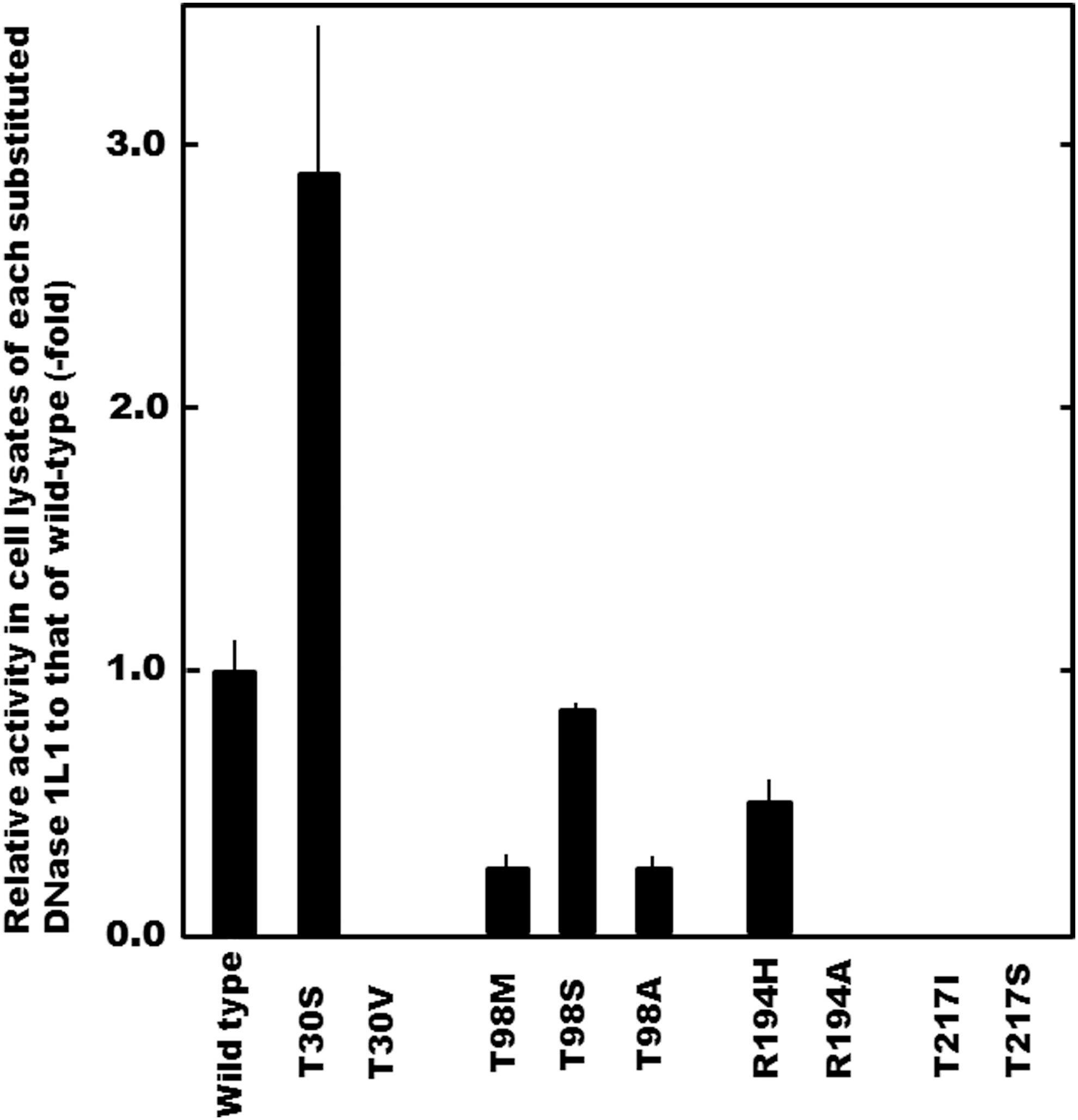
Relative activity of amino acid-substituted variant constructs related to non-synonymous SNPs to that of the wild-type DNase 1L1. The DNase 1L1 activity in the lysates derived from the cells transfected with each construct was assayed by the SRED method (Ueki et al., 2010a). The constructs, T30V and T217S, exhibited no DNase 1L1 activity detectable under our assay condition. The bars present SD (n=4).
A simple procedure for genotyping each SNP in DNASE1L1 using a PCR-RFLP method
A simple and novel genotyping procedure was developed using PCR-RFLP for all of the 18 SNPs in DNASE1L1, those for the other 3 SNPs having been developed previously (Ueki et al., 2010a). As the substitution sites corresponding to the 12 SNPs in DNASE1L1 neither suppressed nor created any known restriction enzyme recognition sites, we employed a mismatched PCR-amplification method for genotyping (Yasuda et al., 1995); incorporation of a deliberate mismatch close to the 3′-terminus of a PCR primer allows the creation of a recognition site for each enzyme. After digestion of the amplified DNA fragment for each SNP, the appearance of the expected product, as shown in Supplementary Table 1, derived from the respective alleles in each SNP, allowed us to determine the genotypes easily. To confirm the validity of the genotyping results obtained by these methods, direct sequencing analysis of the genomic DNA region including the substitution site of each SNP in DNASE1L1 derived from several representative subjects was performed. The genotyping results for each SNP using a PCR-RFLP method were completely congruent with those obtained by direct sequencing, indicating the validity of the novel genotyping methods developed in this study. Furthermore, the technical simplicity and lack of need for any specialized instruments makes the genotyping methods developed in this study advantageous over other methods.
Distribution of the 21 nonsynonymous SNPs in DNASE1L1 for 16 different populations worldwide
The distribution of the genotype and allele frequencies for 18 nonsynonymous SNPs in DNASE1L1 were determined in 16 different populations, including three ethnic groups. Consequently, in addition to three nonsynonymous SNPs in DNASE1L1 examined previously in the same populations (Fujihara et al., 2010; Ueki et al., 2010a), these results allowed us to clarify the genetic distribution of all of the nonsynonymous SNPs (total 21 SNPs) in DNASE1L1 that have already been registered; the allele distributions of the SNPs in our study populations are shown in Supplementary Table S2.
All of the nonsynonymous SNPs in DNASE1L1, except for p.Val122Ile, were found to be distributed in a monoallelic manner; only the predominant allele for each SNP was found in all of the subjects in our study populations. In the NCBI dbSNP, no frequency data are available for p.Leu31Val, p.Gly80Glu, p.Gln170His, p.Arg194His, p.Glu199Asp, p.Asp227Ala, p.Val280Ile, and p.Pro300Ser. Since only the predominant allele was observed in each of these 8 SNPs in the 16 different populations, we were able to demonstrate that these SNPs exhibited extremely low genetic diversity with a heterozygosity of 0.000. Therefore, our population data provide valuable information about the genetic heterogeneity of DNASE1L1. A minor allele, A-allele, for SNP p.Val122Ile was distributed in German and Turk populations, albeit at a low frequency of 0.047, whereas only the predominant allele, G-allele, was observed in the other study populations, showing that p.Val122Ile is Caucasian-specific.
All of the 21 nonsynonymous SNPs in the gene, except for SNP p.Val122Ile in Caucasian populations, exhibited a monoallelic distribution in all of the 16 different populations examined, including three ethnic groups, allowing us to conclude that with regard to the nonsynonymous SNPs DNASE1L1 exhibits extremely low genetic heterogeneity. Overall, it could be assumed that for all of the nonsynonymous SNPs in DNASE1L1, only two haplotypes classified on SNP p.Val122Ile are distributed in Caucasian populations, whereas only one haplotype composed of the predominant allele in each SNP is distributed in other populations worldwide.
It is especially noteworthy that in six SNPs affecting the DNase activity, p.Thr98Met, p.Gln109Glu, pVala115Met, p.Arg194His, p.Thr217Ile, and p.Asp227Ala, each of the alleles resulting in reduction/abolishment of the activity was not distributed worldwide, indicating that all of the nonsynonymous SNPs in DNASE1L1 affecting the catalytic activity showed no genetic heterogeneity in any of the populations examined. These findings indicate that these SNPs in DNASE1L1 may, in general, exert no influence on the levels of the DNase activity in human populations. Thus, it can be assumed that DNASE1L1 is generally well conserved, and thereby full levels of the enzymatic activity in the context of these nonsynonymous SNPs are retained in human populations.
Discussion
In the present study, we have continued our ongoing investigation of the genetic and functional characteristics of nonsynonymous SNPs in DNASE1L1 that may potentially serve as a cell surface barrier to endocytosis-mediated transfer of foreign genes. Although all of the 21 nonsynonymous SNPs in the gene, except for SNP p.Val122Ile in Caucasian populations, exhibited remarkably low genetic diversity in all of the 16 different populations examined, including three ethnic groups, it is worth noting that two activity-abolishing and four activity-reducing SNPs were confirmed to be functional (Fig. 1). To our knowledge, this is the first study to have comprehensively demonstrated the distribution of all of the nonsynonymous SNPs in DNASE1L1 in different worldwide populations and also the effect of each SNP on the enzyme activity.
It has been demonstrated that, in the human DNase I family, subjects with a minor allele reducing the activity in nonsynonymous SNPs exhibit a lower level of in vivo DNase activity than those in subjects without the corresponding allele (Yasuda et al., 1999; Yasutomo et al., 2001; Dittmar et al., 2009). Therefore, it seems plausible that subjects who are hetero/homozygous (female) and hemizygous (male) for each minor allele of p.Thr98Met, p.Gln109Glu, p.Val115Met, p.Arg194His, p.Thr217Ile, and p.Asp227Ala, producing DNase 1L1 variants with substantially low DNase activity or no activity as shown in Table 1, may exhibit lower levels of in vivo DNase 1L1 activity than subjects with other genotypes; in particular, a DNase 1L1-deficient phenotype is associated with the minor allele in the two latter SNPs producing a loss-of-function variant. The endogenous DNase I family has been noted as candidate group of endonucleases involved in the breakdown of chromatin during apoptosis and/or necrosis (Counis et al., 2000; Mizuta et al., 2006; Napirei et al., 2006; Fisher et al., 2007). In this context, triggered by nuclear antigens, the clearance of cell debris resulting from cell death through apoptosis and/or necrosis might be primarily involved in the prevention of autoimmune conditions; the DNase I family has been especially highlighted for its possible involvement in the pathogenesis of autoimmune diseases (Tsukumo et al., 2004; Valle et al., 2008; Hedberg et al., 2011). Therefore, we could postulate that a minor allele of activity-abolishing SNPs producing loss-of-function variants of the DNase I family would represent a direct genetic risk factor for autoimmune diseases (Ueki et al., 2009; Yasuda et al., 2010; Kimura-Kataoka et al., 2013). In contrast, among the members of the DNase I family, no association of DNase 1L1 with diseases potentially attributable to a failure in the breakdown of chromatin during apoptosis and/or necrosis through reduction/abolishment of in vivo DNase activity has been reported. Although a 196-bp deletion in the 3′-untranslated region in exon 9 of the DNase 1L1 gene has been shown to be present at a high frequency in patients with Pomp disease (Malferrari et al., 2001), and a partial deletion of the gene has been found in patients with Barth syndrome (Singh et al., 2009), it remains unknown whether reduction/abolishment of in vivo DNase 1L1 activity may be involved in the etiology of these diseases. However, Shiokawa et al., (2007) have suggested that suppression of foreign gene transfer by DNase 1L1 on the cell surface is probably important for maintaining healthy cell communities in vivo, thereby reducing the risk of malignant transformation. Therefore, it seems plausible that a minor allele of p.Thr217Ile and p.Asp227Ala, producing a loss-of-function DNase variant, might be a genetic risk factor for malignant transformation.
We have been conducting genetic and expression analyses of nonsynonymous SNPs in the DNase I family genes to evaluate whether these SNPs are functional and potentially implicated in various diseases. As a result, we have demonstrated that almost all of the nonsynonymous SNPs in the genes encoding other members of the human DNase I family, DNase I (Yasuda et al., 2010; Ueki et al., 2013b), I-like 2 (Ueki et al., 2013a), and I-like 3 (Ueki et al., 2009; 2013b), together with DNase II (Ueki et al., 2010b; Kimura-Kataoka et al., 2012, 2013), exhibit a monoallelic distribution in the same study populations, similar to those of DNase 1L1. Thus, with regard to nonsynonymous SNPs potentially resulting in alterations of in vivo DNase activity, the human DNase I family genes show remarkably low genetic diversity. These facts allow us to conclude that the human DNase family including DNase II has, on the whole, been well conserved at the protein level during the evolution of human populations.
Footnotes
Acknowledgments
DNA samples of Bloodstain samples of the Ovambo and Turkish populations were kindly provided by Dr. B. Brinkmann. Blood samples of Korean and Mongolian populations were kindly provided by Dr. K. Shiwaku. This study was supported in part by Grants-in-Aid from Japan Society for the Promotion of Science (25460864 to M.U., and 22249023 to T.Y.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.