
Abstract
Mucin 1 (Muc1) is a tumor-associated glycoprotein and has an important role in cancer progression and metastasis. The aim of the study was to explore the effects and mechanism of Muc1 silencing on proliferation and tumor-forming capacity of colon cancer cell lines. In the present study, we first screened the colon cancer cell lines with high expression of Muc1 by western blot analysis. Then, the effective siRNA was selected and used for silencing endogenous Muc1. The results by MTT and vitro scratch assay showed that interference of Muc1 could effectively inhibit HCC2998 proliferation and migration (p<0.01). Also, colony-forming ability in Muc1-siRNA groups was significantly decreased compared with the control group (p<0.01). Cell cycle is detected by flow cytometry that suggested that Muc1-siRNA1 induced cell cycle arrest at S phase in HCC2998. Next, the expression and distribution of β-catenin in cytoplasm and the nucleus was detected by western blot and the results revealed that the expression of β-catenin was redistributed in Muc1-siRNA group. A higher β-catenin level was detected in cytoplasm, while a lower β-catenin level was located in nucleus, compared with controls (p<0.05). The tumorigenicity experiments showed that inhibition of Muc1 could significantly suppress the growth of HCC2998 in nude mice models (p<0.01). This study would effectively provide new clues for colon cancer therapy.
Introduction
C
It has reported that the production of mucin by cancer cells affects metastasis in patients (Rachagani et al., 2012; Kim et al., 2013). Mucins are a family of high-molecular-weight, heavily glycosylated proteins. Mucin 1 (Muc1), cell surface associated (Muc1) or polymorphic epithelial mucin, is a membrane-bound protein that is a member of the mucin family (Sinn et al., 2013). The hyperglycosylated transmembrane protein Muc1 has a core protein mass of 120–225 kDa that increases to 250–500 kDa with glycosylation (Gao et al., 2005; Fatrai et al., 2008; Woenckhaus et al., 2008). It has been detected for high expression in a variety of tumor tissues, including breast cancer, lung cancer, pancreatic cancer, colon cancer, and ovarian cancer. Muc1 expression correlates with high grade, metastasis potential, and poorer survival rate in malignant cancers, and has been made the candidate of the therapeutic targets and biomarkers for cancers (Remmers et al., 2013). Although the hyperglycosylated transmembrane protein isoform of Muc1 is aberrantly overexpressed in colon cancer, the biological significance and the mechanisms on tumor progression are still unclear and incompletely understood.
Additionally, abnormal activation of Wnt/β-catenin signaling pathway has been considered to play a critical role in carcinogenesis and metastasis of malignant tumors (Arend et al., 2013; Mo et al., 2013; Yao et al., 2013). In normal conditions, cytosolic β-catenin is constitutively phosphorylated by glycogen synthase kinase 3β (GSK3β) and then ubiquitinated and degraded by ubiquitin-proteasome pathway (Kavitha et al., 2013). Actually, disorders of Wnt/β-catenin signaling pathway are found in colon cancers, resulting in the accumulation of unphosphorylated β-catenin in the cytoplasm. Then, the accumulated β-catenin translocates into the nucleus, in which, Wnt/β-catenin-mediated target genes, such as T-cell factor (TCF) and lymphoid-enhancing factor, are activated (Paez et al., 2013). In the present study, we studied the role of Muc1 in proliferation and metastasis of colon cancer cell line HCC2998, and further explored its potential mechanism. Finally, the in vivo experiments were performed and the results demonstrated that the lentiviral vector of Muc1 could effectively inhibit the progression of colon cancer in nude mice model. It would be useful and helpful to provide new clues for diagnosis and prognosis of colon cancer.
Materials and Methods
Cell lines
Colon cancer cell lines HCC2998, HCT15, HT29, Colo205, and HCT116 were cultured in DMEM supplemented with 10% fetal calf serum and antibiotics (100 units/mL penicillin G sodium and 100 μg/mL streptomycin).
MTT assay
MTT assay was performed as described previously (Sylvester, 2011; Li et al., 2012; Sarzaeem et al., 2012). Briefly, cell proliferation was evaluated over a 3-day time course. Ten microliters of 5 mg/mL MTT agent (Sigma) was added into the medium and incubated for 4 h. Then, 150 μL of DMSO was added to incubate for further 15 min. The plates were then read on a microplate reader using a test wavelength of 490 nm and a reference wavelength of 570 nm.
Rescue experiments
For rescue experiments, siRNA-resistant Muc1 overexpression cell line was constructed in our lab. Two nucleic acids within the siRNA-targeted regions were mutated to alleviate and abolish the siRNA effect. Mutations were generated using the QuikChange II Site-Directed Mutagenesis Kit (Stratagene), according to the manufacturer's protocol.
Cell transfection
Three pairs of siRNA specific to Muc1 were designed and synthesized by Jima Corporation. Transfection of cells was performed using Oligofectamine (Invitrogen) according to the manufacturer's protocol. Muc1 shRNA (h) Lentiviral Particles (sc-35985-V) were purchased from Santa Cruz.
Clone formation assay
For colony-formation assay, cells were transfected with Muc1-siRNA and negative control siRNA and plated into six-well plates and cultured for 12 days. The medium was refreshed every 3 days. Methanol was used to fix the surviving colonies (≥50 cells per colony). The colonies were stained with 1.25% crystal violet and counted under a light microscope.
Cell cycle analysis
Seventy-two hours after transfection, cells were harvested and washed twice in phosphate-buffered saline. Then, the cells were fixed in 70% ethanol on ice for more than 30 min. They were stained with propidium iodide solution (50 μg/mL propidium iodide, 50 μg/mL RNase A, 0.1 mM EDTA, and 0.1% Triton-X). Cell cycles were analyzed based on DNA contents by FACS using a flow cytometer (BD Biosciences).
Real-time PCR analysis
Total RNA was extracted from HCC2998 cells using TRIzol (Takara, Inc.) according to the protocols. cDNA was generated using the PrimeScript® RT reagent kit (Takara Co., Ltd.) in a 20 μL final reaction volume according to the protocols. Real-time quantitative PCR assay was performed to evaluate Muc1 expression using the SYBR Premix Ex TaqTM II (Takara Co., Ltd.) on ABI 7500 system according to the instructions of manufacturer. The sequence of primers on the genes was as follows:
Muc1: 5′-CTGGTCTGTGTTCTGGTTGC-3′ and 5′-CCA CTGCTGGGTTTGTGTAA-3′
c-Myc: 5′-GCTGCTTAGACGCTGGATTT-3′ and 5′-CAC CGAGTCGTAGTCGAGGT-3′
Cyclin D1: 5′-AGACCTTCGTTGCCCTCTGT-3′ and 5′-AGTTGTTGGGGCTCCTCAG-3′
β-actin: 5′-GCACAGAGCCTCGCCTT-3′ and 5′-GTTG TCGACGACGAGCG-3′
Western blotting
The samples were prepared and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis as previously described (Chen et al., 2013; Murugan et al., 2013; Xie et al., 2013). The antibodies used included the anti-Muc1, anti-IκB-α, anti-Lamin B, anti-β-actin, and horseradish peroxidase-conjugated goat anti-mouse secondary antibody. They were all purchased from Santa Cruz Biotechnology Incorporation with an ECL detection kit (Amersham).
In vivo tumor xenograft study
The male BALB/c(nu/nu) mice were purchased from Experimental Animal Center of the Chinese Academy of Sciences Shanghai (Shanghai, China) and kept in pathogen-free environment with a 12-h light/dark cycle. All animal experiments and care were conducted in a manner conforming to the Guidelines of the Animal Care and Use Committee of Xinhua Hospital Affiliated to Shanghai Jiaotong University School of Medicine. HCC2998 cells were injected subcutaneously into the flanks of the mice (2×106 cells in 200 μL of medium). The mice were randomly divided into three groups: untreated control, negative control shRNA, and Muc1 shRNA group (n>5). Tumor volume was measured using calipers according to the following formula: tumor volume (mm3)=3.14×L×W×W/2, where L is the length and W is the width.
Statistical analysis
All the experiments were performed and repeated at least three times. The samples of real-time PCR were performed in triplicate. All statistical analyses were performed using the SPSS statistical package (11.5). Results are depicted as the mean±standard error of the mean. p-Values<0.01 were defined as significant statistical difference.
Result
Expression analysis of Muc1 in colon cancer cell lines
First, Muc1 expression was screened in a panel of 5 colorectal cancer cell lines. As shown in Figure 1, increased expression of Muc1 in HCC2998 and colo205 was confirmed by real-time PCR and western blot analysis. The expression of Muc1 in HCC2998 and colo205 was much higher than other colon cancer cell lines, such as HCT116, HCT15, and HT29. So, in the present study, we used the colon cancer cell line of HCC2998 as the cell model.

Expression analysis of Muc1 in colon cell lines.
siRNA1 of Muc1 is the most effective sequence for silencing Muc1 gene
To knock down the Muc1 in HCC2998, we designed three pairs of special siRNA of Muc1. To test the interference effects of siRNAs, we used the three pairs of siRNAs to transfect the colon cancer cell line HCC2998. The silencing effects were determined by western blot and real-time PCR, and the results were shown that siRNA1 and siRNA3 were specific to Muc1 gene and could effectively interfere the expression of Muc1 (Fig. 2). Here, the Muc1-siRNA1 was used in the following experiment.
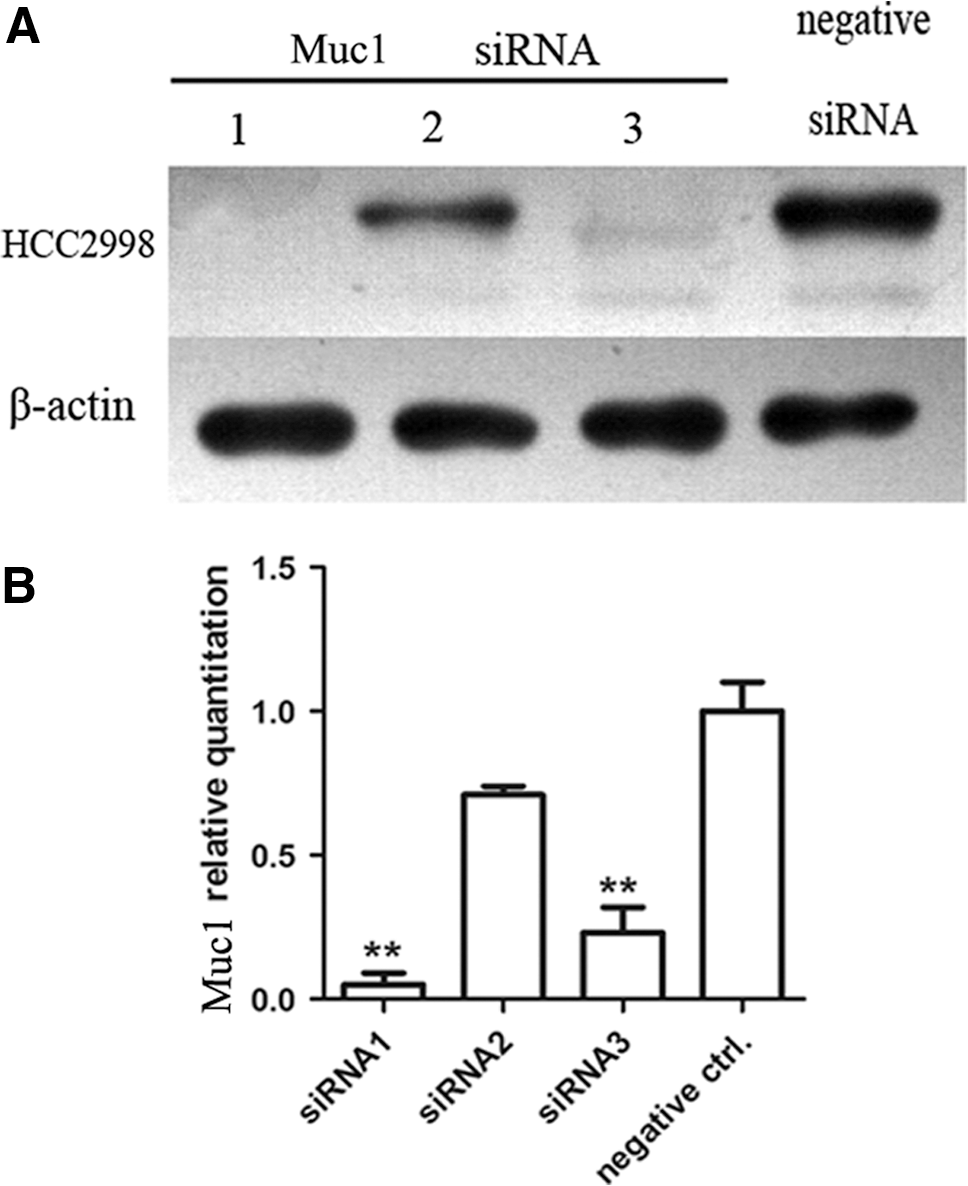
Muc1-siRNA is the most effective sequence for silencing Muc1 gene. The cells were plated in 48-well plates and three pairs of Muc1-siRNA were transfected into HCC2998. After 48 h, the expression of Muc1 gene was detected by western blot analysis
Muc1-siRNA1 effectively inhibits HCC2998 cell growth
To examine the effects of Muc1-siRNA on cellular proliferation, MTT assay was performed. The Muc1-siRNA and negative control siRNA were used to transfect colon cancer cell line HCC2998.
As demonstrated by MTT assays (Fig. 3), the HCC2998 transfected by Muc1-siRNA proliferated much slower than the controls. And the results of Muc1 rescue assay revealed that the Muc1-defective colon cell line HCC2998 could restore the higher proliferative capacity as control cell lines. Here, the untreated cells and the cells transfected by negative control siRNA were used as controls.

Interference of Muc1 effectively inhibits HCC2998 and Colo205 proliferation by MTT assay.
The inhibitory effect on HCC2998 cell growth is further confirmed by colony-formation assay
Compared with cells transfected with negative siRNA control, the number of colonies was significantly decreased in cells transfected with Muc1-siRNA (Fig. 4). Taken together, Muc1-siRNA significantly inhibits the growth of colon cancer cell line HCC2998 and acts as a potential tumor suppressor.
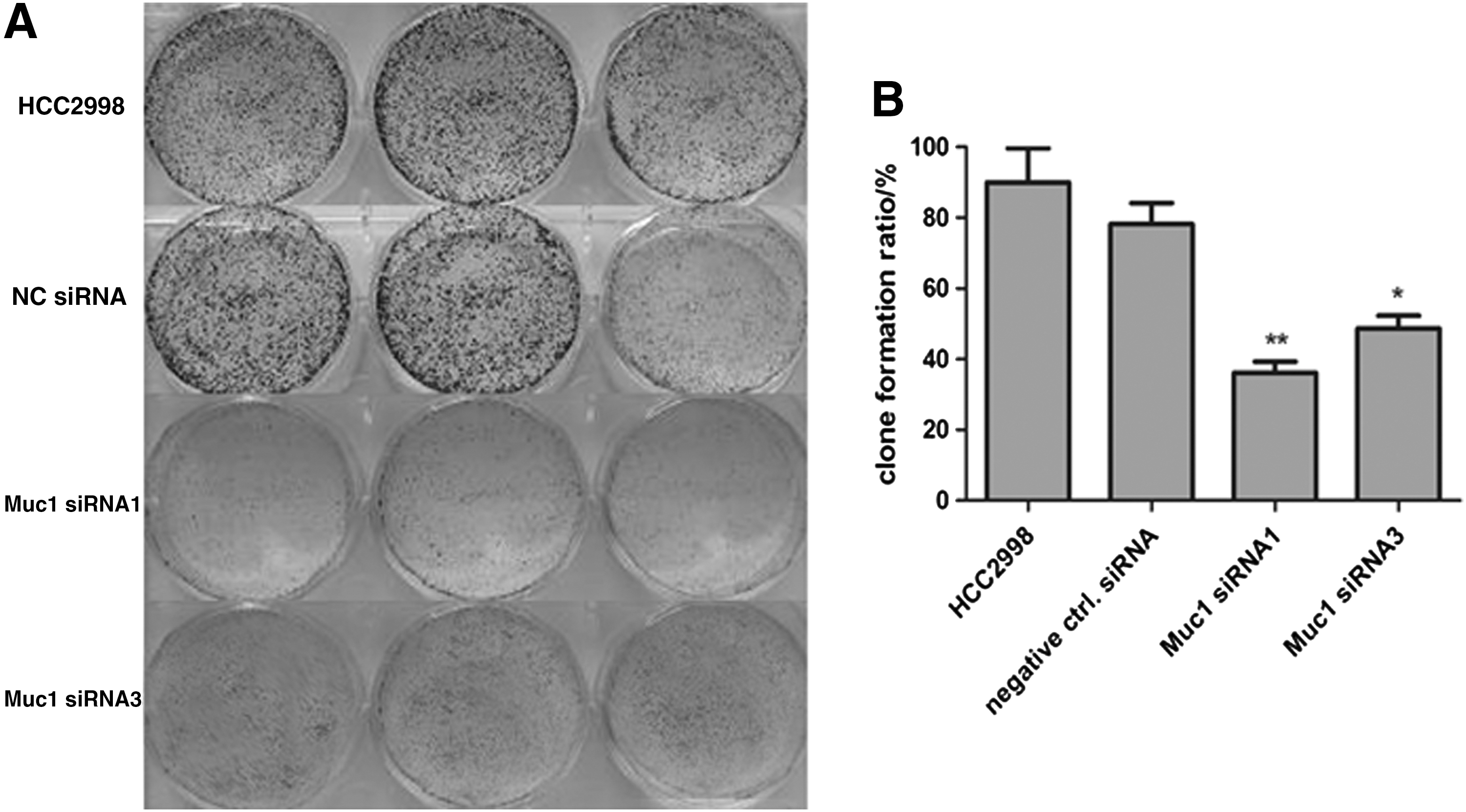
Transfection with Muc1-specific siRNAs inhibits the colony formation of HCC2998.
Muc1-siRNA1 induces cell cycle arrest of the colon cancer cell line
It has been proved that Muc1-siRNA inhibited the cell growth of colon cancer cells. Next, we examined the contribution of cell cycle arrest in different group of cells. As shown in Figure 5, compared with scramble siRNA control, cell cycle was arrested at S phase when cells were transfected with Muc1-siRNA. The percentage of S phase was increased from 22.48% to 40.63%.
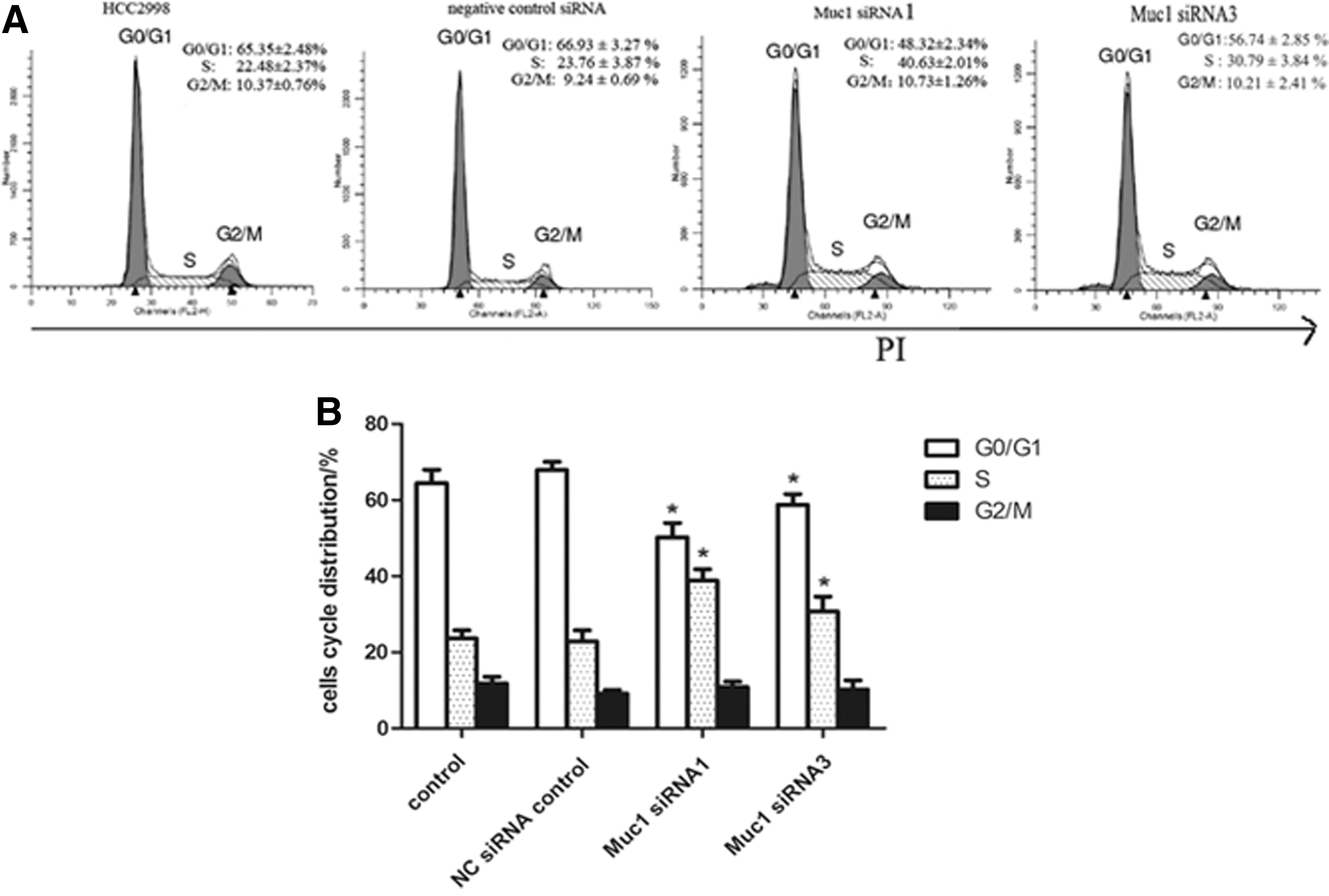
Muc1-siRNA1 and Muc1-siRNA3 induce cell cycle arrest of HCC2998 and the cell cycle is detected.
Muc1-siRNA inhibits HCC2998 cell migration
As colon cancer is a type of highly malignant tumor with local invasion and distant metastasis, we next explored the effects of Muc1-siRNA on cell invasion by in vitro scratch assay. As shown in Figure 6, the migrated cells of HCC2998 were significantly decreased after transfection with Muc1-siRNA, compared with the untreated cells and the cells transfected with negative control siRNA (p<0.01), suggesting that the siRNA of Muc1 inhibited the invasive potential of HCC2998 cells.
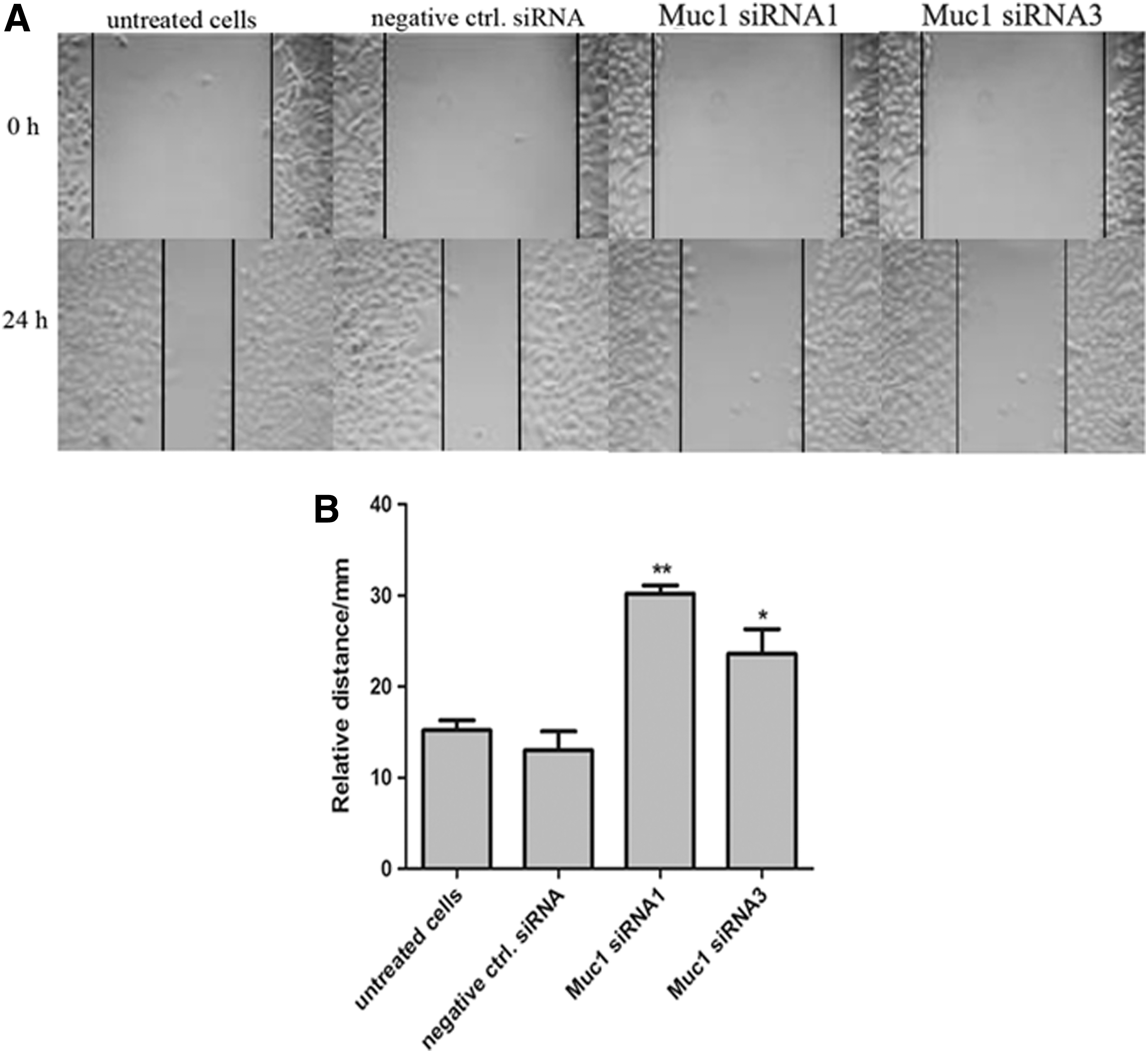
Muc1-specific siRNAs inhibit HCC2998 cell invasion by scratch assay.
Muc1-siRNA promotes redistribution of β-catenin and reduces the expression of cyclin D1 and c-Myc
The migratory activities of colon cancer cells were partly promoted through cellular translocation of β-catenin into nucleus. Then, we detected the expression and distribution of β-catenin in cytoplasm and the nucleus, respectively. As shown in Figure 7A and B, in Muc1-siRNA-transfected colon cells, the expression of β-catenin significantly increased in cytoplasm and decreased in nucleus compared with that in untreated cells or the cells transfected by negative control siRNA. It demonstrated that Muc1-siRNA inhibited the translocation of β-catenin from cytoplasm into nucleus and induced the redistribution of β-catenin. Under pathological conditions, stabilized β-catenin translocated to the nucleus where it interacted with TCF/LEF transcription factors resulting in transcription of target genes, such as cyclin D1 and c-Myc. Thus, we investigated whether Muc1-siRNA-mediated inhibition of colon cancer cell growth is due to repressed expression of cyclin D1 and c-Myc. The results by real-time PCR and western blot demonstrated that Muc1-siRNA was able to decrease the expression of cyclin D1 and c-Myc, which were involved in cell cycle progression.
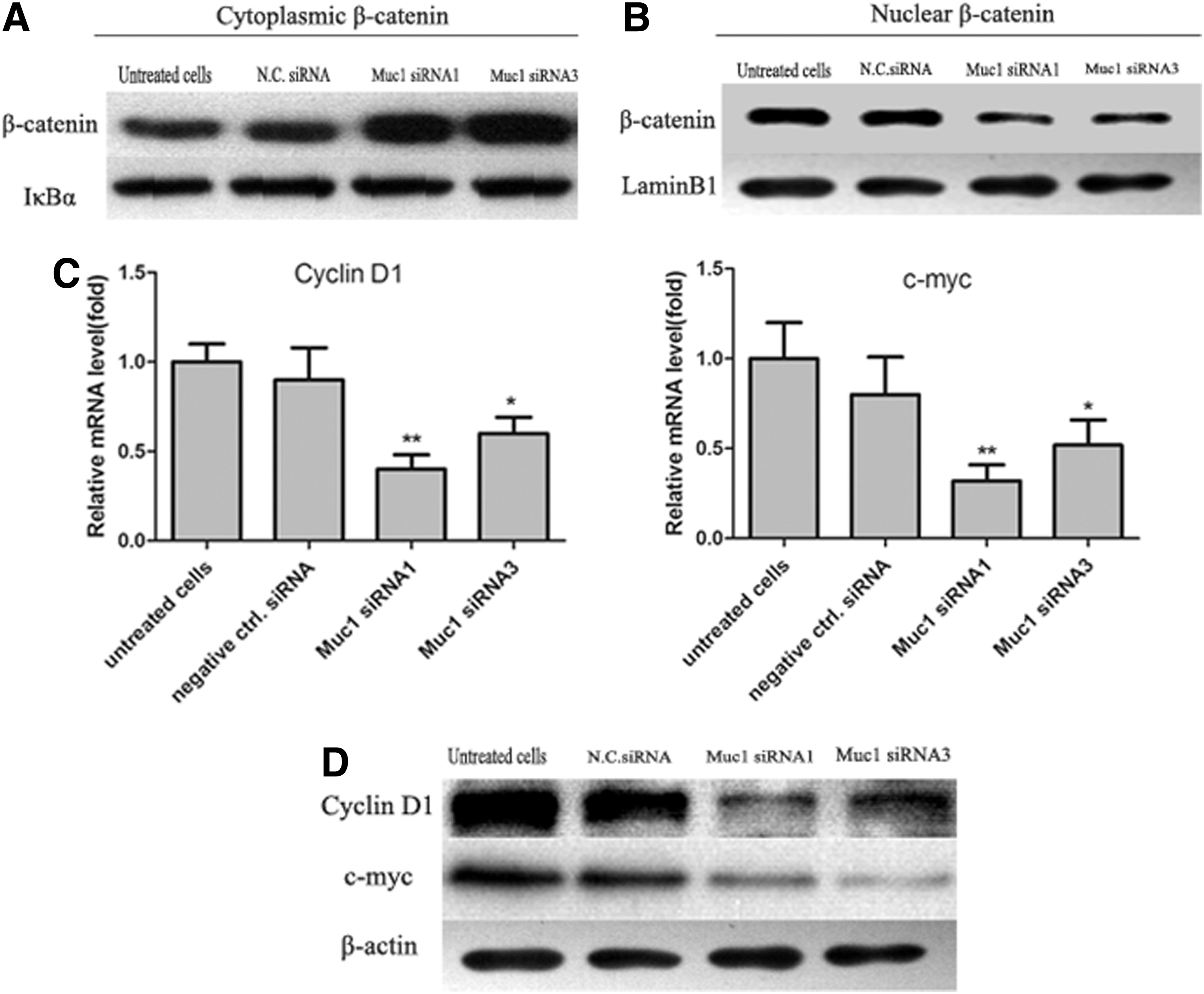
Muc1-siRNA1 and Muc1-siRNA3 promote redistribution of β-catenin and reduce the expression of cyclin D1 and c-Myc. Cell lysates were used for detection of β-catenin in cytoplasm
Muc1 cytoplasmic tail interacts with β-catenin
We have known that Muc1-specific siRNA inhibited the translocation of β-catenin from cytoplasm to nucleus and reduced the expression of cyclin D1 and c-Myc. Next, we explored how the membrane-bound protein Muc1 interacted with the proteins on β-catenin signaling pathway. Thus, the co-immunoprecipitation assay was performed, and the results demonstrated that the Muc1 cytoplasmic tail (Muc1-CT) could directly interact with β-catenin in HCC2998 cell line (Fig. 8).
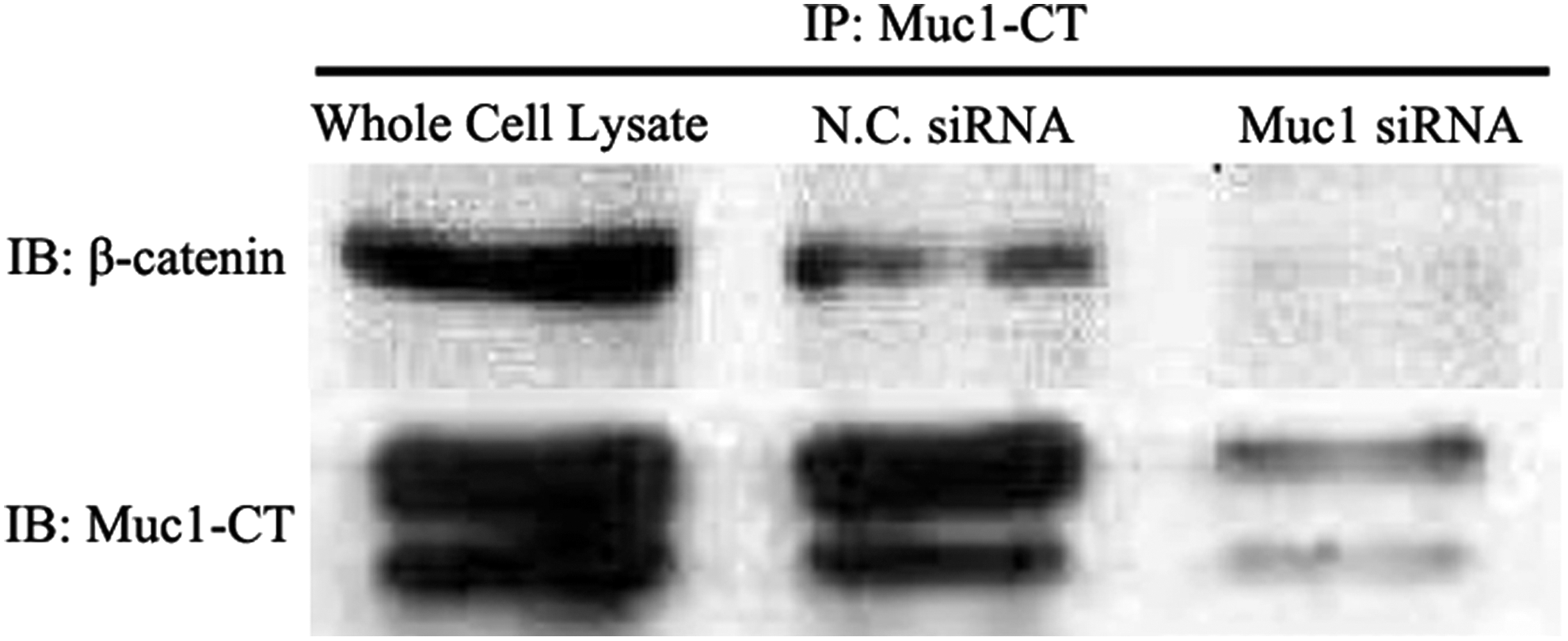
Muc1 cytoplasmic tail (Muc1-CT) interacts with β-catenin. Cell lysates of HCC2998 were used for immunoprecipitation with anti-Muc1-CT antibodies. And the immunoblotting assay was performed with β-catenin and anti-Muc1-CT antibodies. Here, the untreated cells or the cells transfected by scramble siRNA were used as controls.
Muc1-siRNA exhibited antitumor activity in a xenograft nude mouse model
The Muc1-siRNA effectively inhibited the proliferation and invasion of colon cancer cell line HCC2998 in vitro. To detect the antitumor activity of Muc1-RNAi in vivo, we used a nude mouse xenograft model. First, a lentiviral vector was used to construct the cell line HCC2998, which was stably transfected with Muc1-shRNA or NC-shRNA. Then, the untreated HCC2998, Muc1-shRNA-transfected HCC2998, and negative control shRNA-transfected HCC2998 were subcutaneously implanted into nude mice. As shown in Figure 9, compared with other two control groups, the tumors grew much lately and slowly in Muc1-shRNA group. However, body weight had not significantly changed among three groups (p>0.05), which meant that the treatment of Muc1-shRNA and NC-shRNA had no toxicity in the experiment.
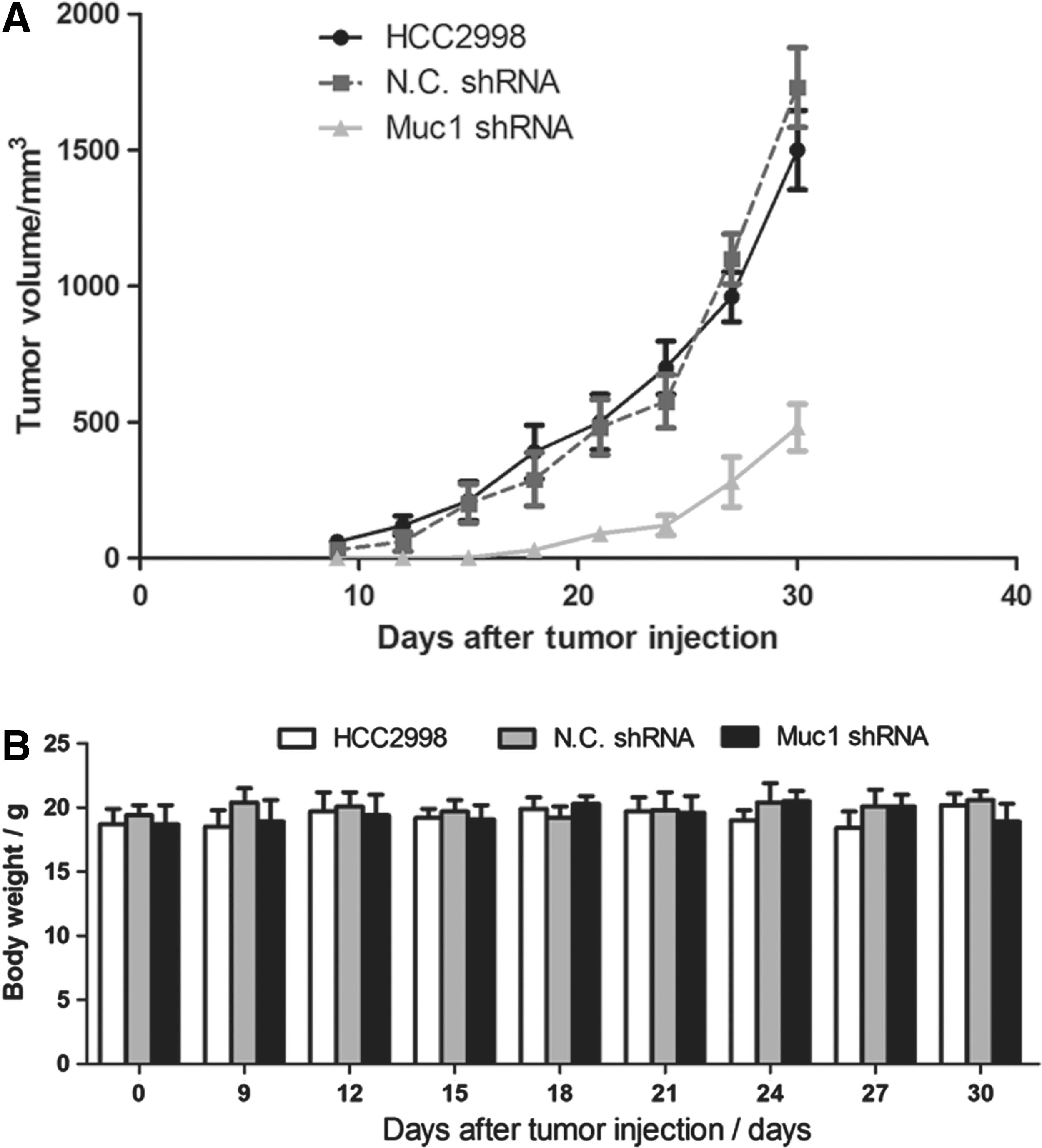
Muc1-shRNA exhibits antitumor activity in a xenograft nude mouse model. The nude mice were challenged subcutaneously with 5×105 colon cancer cells to the back area. Here, each group contained more than six mice.
Discussion
Colorectal cancer is one of the malignant tumors with high incidence and mortality. It is more and more commonly seen in the world. In this article, we focused on Muc1, a highly tumor-associated glycoprotein. It is reported that the abnormal expression of Muc1 can induce malignant transformation of cells, resist apoptosis, and increase the tumorigenicity activity of cancer cells (Huang and Chen, 2011; Karamitopoulou et al., 2011). Thus, a pair of siRNAs specific to Muc1 were selected and used to silence the endogenous Muc1 in colon cancer cell lines with high expression of Muc1. Our results demonstrated that interference of Muc1 effectively inhibited the proliferation and progression of colon cancer cells. The in vitro scratch assay results showed that cell growth of HCC2998 was significantly decreased after transfection of Muc1-siRNA, suggesting that silencing of Muc1 could markedly reduce cell migration and proliferation, which was consistent with the results of MTT assay. These were also consistent with that detected by colony formation. The colony-formation activities were obviously suppressed in Muc1-siRNA group. All of the data from in vitro assays definitely demonstrated that transfection of Muc1-siRNA can significantly inhibit the growth of colon cancer cells.
It has been well established that Wnt/β-catenin signaling pathway is causally involved in the progression and metastasis of colorectal cancers (Brabletz et al., 1999; Brabletz et al., 2000; Fang et al., 2009; Sack et al., 2011). The progression of cancer is accompanied by abnormal accumulation of β-catenin in nucleus of cells (Zhai et al., 2002; Park et al., 2013). We have identified that interference of Muc1 suppressed HCC2998. Next, we tried to explore whether interference of Muc1 was associated with Wnt/β-catenin signaling pathway and further to clarify the mechanism of regulating the progression of cancer. Indeed, a higher β-catenin level was detected in cytoplasm, while a lower β-catenin level was located in nucleus, compared with controls, which revealed that redistribution of β-catenin was induced by Muc1-RNAi technology. Additionally, β-catenin/TCF-targeted downstream genes were also downregulated in HCC2998 and colo205 human colon cancer cells, such as c-Myc and cyclin D, which were involved in cell cycle regulation (Xu et al., 2010; Park et al., 2013). These results demonstrated that the antiproliferative effects of Muc1-RNAi in colorectal cancer cells might partly be associated with the downregulation of Wnt signaling.
It is reported that the Muc1-CT interacts with β-catenin through a conserved -S-X-X-X-X-X-S-S-L- motif (where X represents any amino acid), which also interacts with other β-catenin-binding partners, including the cadherins (Schroeder et al., 2003; Molock and Lillehoj, 2006). The possible mechanism should be that Muc1-CT interacting with β-catenin could make it to be undegraded by GSK3β, stabilize the structure of β-catenin and promote β-catenin translocating to nucleus, and activate the target genes in the downstream of Wnt/β-catenin signaling pathway (Li et al., 2001; Aragaki et al., 2008). Additionally, Muc1-CT binding with β-catenin decreased the amount of E-cadherin-catenin, to a certain extent, which decreased cell adhesion and promoted cell migration of the tumor cells (Yuan et al., 2007; Niv, 2008). Our results by co-immunoprecipitation assay also proved that Muc1-CT could directly interact with β-catenin in the colon cancer cell HCC2998. However, interference of Muc1 with specific siRNA would weaken the interaction between Muc1-CT and β-catenin, which inhibited the nuclear translocation of β-catenin and the expression of the target genes in the course of tumorigenesis. Thus, interference of Muc1 played an antitumor role in colon cancer progression.
To be consistent with in vitro findings, we successfully constructed a xenograft nude mouse model by lentiviral delivery systems. As expected, tumor growth was also suppressed in a nude mouse xenograft model bearing HCC2998 cells by interferencing Muc1. No overt toxicity was found under these experimental conditions.
In summary, our data show that interference of Muc1 significantly inhibits the proliferation and migration of colon cancer cell HCC2998 partly by downregulating Wnt/β-catenin signaling pathway. It is also proved that Muc1 is a potential biomarker for colon cancer diagnosis and serves as a new target for colon cancer therapy. This study would effectively provide new clues for colon cancer therapy.
Footnotes
Disclosure Statement
No competing financial interests exist.