
Abstract
DNA and Cell Biology
officially retracts the paper entitled, “HBx Elevates Oncoprotein AEG-1 Expression to Promote Cell Migration by Downregulating miR-375 and miR-136 in Malignant Hepatocytes,” by Jing Zhao, Wenjie Wang, Yuxian Huang, Jing Wu, Mingquan Chen, Peng Cui, Wenhong Zhang, and Ying Zhang (DNA and Cell Biology; doi: 10.1089/dna.2014.2376).
After the journal has published a post-publication correction to the article,1 additional considerations were made after revisiting the recently released guidelines by STM: Advancing Trusted Research.2 These recommendations indicate that replicative experiments should not be permitted if they were not generated at the time of the original experimentation.3[section A2(4)]
The Editor and Publisher of DNA and Cell Biology are committed to preserving the scientific literature and the community it serves, and abides by all standard guidelines for, and proper protocols of, scientific publishing.
References
1. [No author(s) listed]. Correction to: HBx elevates oncoprotein AEG-1 expression to promote cell migration by downregulating miR-375 and miR-136 in malignant hepatocytes by Zhao et al. Epub 2021 Sep 27. doi: 10.1089/dna.2014.2376.correx
2. Working Group on Image Alterations and Duplications. STM STEC Working Group on Image Alterations and Duplications.
3. STM Working Group on Image Alteration and Duplication Detection. STM Recommendations for handling image integrity issues.
Introduction
Hepatocellular carcinoma (HCC) is one of the most common worldwide malignancies with high morbidity and mortality. Chronic hepatitis B virus (HBV) infection is a major risk factor (Kew, 2010). The hepatitis B viral X (HBx) protein encoded by HBV plays important roles in hepatocarcinogenesis (Zhang et al., 2006) and high HCC incidence is observed in the HBx-transgenic mice (Kim et al., 1991). HBx is a multifunctional regulator involved in modulating the expression of many viral and cellular genes, which affect cell survival, apoptosis, and intracellular signal transduction, and stimulate invasion and metastasis of HCC (Zhang et al., 2006). For instance, HBx interacted with oncoprotein AIB1 and increased the stability of AIB1. Thus, HBx cooperated with AIB1 to promote matrix metallopeptidase 9 (MMP-9) expression and enhance HCC cell invasiveness (Liu et al., 2012). HBx can also affect some microRNA (miRNA) expression. Let-7a was the first established miRNA downregulated by HBx, which led to the upregulation of STAT3 to increase cell proliferation in HBx-expressing cells (Wang et al., 2010).
Astrocyte elevated gene-1 (AEG-1), also named as metadherin (MTDH) or 3D3/LYRIC, has been regarded as an important oncogene, which is located in 8q22 and this region is amplified in breast cancer (Hu et al., 2009) and HCC (Yoo et al., 2009). AEG-1 overexpression has been observed in various cancers and is associated with poor clinical outcomes (Yoo et al., 2011). The transgenic mouse expressing AEG-1 in specific hepatocyte developed multinodular HCC with steatosis when treating with N-nitrosodiethylamine (Srivastava et al., 2012). The positive feedback loop interplay existed between AEG-1 and oncogenic Ha-ras requiring PI3K-Akt pathway, and synergistically resulted in tumor aggressiveness (Lee et al., 2006, 2008). AEG-1 was the downstream target of Ha-ras. Ha-ras could activate PI3K-Akt pathway to induce transcription factor c-MYC recruitment in E-box elements of the AEG-1 promoter region, and consequently augmented AEG-1 expression and promoted cell proliferation and invasiveness (Lee et al., 2006). Additionally, AEG-1 overexpression enabled Akt phosphorylation activation to induce cell survival under serum starvation by inhibiting apoptosis (Lee et al., 2008). Further, AEG-1 upregulation could activate NF-κB, Wnt/β-catenin, and MAPK/ERK signaling cascades, which induced many malignant phenotypes, including chemoresistance, angiogenesis, metastasis, and invasion (Yoo et al., 2011). Hence, exploring the molecular mechanisms of AEG-1 overexpression is helpful to prevent tumor development.
AEG-1 expression is regulated by many factors. AEG-1 was originally identified as the inducible gene by human immunodeficiency virus and tumor necrosis factor α in primary human fetal astrocytes (Su et al., 2002, 2003). Recently, some miRNAs were demonstrated to reduce AEG-1 expression. For instance, miR-375 could target AEG-1 to stimulate G1 arrest and apoptosis, and inhibit cell migration and invasion in head and neck cancer (Hui et al., 2011), and HCC (Fan et al., 2012); miR-136 triggered apoptosis partially by targeting AEG-1 in human glioma (Yang et al., 2012); and miR-26a was found to negatively regulate AEG-1 to enhance apoptosis in breast cancer (Zhang et al., 2011). Previous study reported that AEG-1 expression displayed increasing trend from normal liver tissue to hepatitis B and HBV-related HCC samples (Gong et al., 2012), which suggests that HBV may be involved in augmenting AEG-1 expression. Considering that HBx can increase the levels of some oncoproteins, such as AIB1 (Liu et al., 2012) and c-Myc (Kalra and Kumar, 2006), we wonder whether HBx contributes to AEG-1 upregulation and collaborates with AEG-1 to promote HCC development.
In this study, we found that HBx could elevate AEG-1 protein level without affecting its transcription, which in turn was involved in HBx-induced cell migration. The miR-375 and miR-136 downregulated by HBx accounted for HBx-mediated AEG-1 elevation and cell migration.
Materials and Methods
Cell culture
The HepG2-pcDNA3-HBx cells had stable HBx expression in human HCC cell line HepG2, and the HepG2-pcDNA3 was its vector control cells. These cells were kindly provided by Dr. Zhongtian Qi (Changhai Hospital, the Second Military Medical University of China) and were constructed as described previously (Li et al., 2009). The control HepG2-pcDNA3 and HepG2-pcDNA3-HBx stable cells were named HepG2-Ctr and HepG2-HBx in this study. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) and 200 μg/mL G418 (Gibco) at 37°C in 5% CO2-humidified atmosphere. Another HCC cell line SMMC-7721 was also grown in DMEM with 10% FBS at 37°C in 5% CO2 atmosphere.
Antibodies
Specific antibodies used for western blot were as follows: anti-HBx (Santa Cruz), anti-AEG-1 (Abcam), anti-Myc and anti-β-Actin (Sigma), and anti-MMP-9 (Cell Signaling Technology).
Plamids, miRNA mimics, and siRNA oligos transfections
Transfection was conducted with Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions. Cells at 80% confluence were transfected with plasmids pCMV-Myc-HBx. When transfecting siRNA oligos and miRNA mimics (RiboBio), cell densities were about 60%. Then, 40 nM of siRNA oligos or 60 nM of miRNA mimics was, respectively, introduced into cells as indicated, and cells were harvested 48 h after transfection. The siRNA oligos sequences targeting AEG-1 were siRNA1-AEG-1: 5′ GGAGGAGGCUGGAAUGAAAdTdT 3′ and siRNA2-AEG-1: 5′ GGUCU CAGAUGAUGAUAAAdTdT 3′. The siRNA oligos sequences targeting HBx were siRNA1-HBx: 5′ CCUGGAUCGAAGAAUACAUTT 3′ and siRNA2-HBx: 5′ GGGAGGAGAUUAGGUUAAATT 3′. Negative control siRNA: 5′-AACUCCUGCCUCCUUAUGUAUUUdTdT-3′.
Western blot
Cell lysates were separated by 12% SDS-PAGE and transferred to nitrocellulose membranes (GE Healthcare). The membrane was blocked with 5% milk in TBS (20 mM Tris-HCl and 137 mM NaCl [pH 7.6]) for 1 h at room temperature, and incubated with specific primary antibodies at 4°C overnight, followed by incubation with horseradish-peroxidase-conjugated secondary antibody at room temperature for 1 h. Then, the protein bands of interest were visualized with enhanced chemiluminescence detection reagents (Santa Cruz Biotechnology).
RNA extraction and qRT-PCR analysis
Total RNA, including miRNA, was extracted and purified with miRNeasy mini kit (Qiagen). The RNA concentration was measured by O.D. 260/280 nm absorbance with NanoDrop spectrophotometer (Thermo). Then, 500 ng of RNA was reverse-transcribed to cDNA with ReverTra Ace kit (Toyobo), and an aliquot of 500 ng RNA was converted to cDNA for miRNA analysis using the miScript RT kit (Qiagen) following the manufacturer's protocols. qRT-PCR for AEG-1 gene level was performed with standard SYBR-Green PCR kit protocol (Takara) and normalized by the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH). miRNA expression detection by qRT-PCR used the miScript SYBR Green PCR kit (Qiagen), the universal reverse primer was provided by manufacturer, and the forward primer was specific to the mature miRNA listed in the Sanger miRBase. The miRNA expression was normalized with U6 small RNA as internal control. All reactions were conducted in triplicate. Specific qRT-PCR primers were as follows:
AEG-1-forward: 5′ TGGGCGGACTGTTGAAGTGGC 3′, AEG-1-reverse: 5′ CCCAGGCTCCTTCATCAACTTCCT 3′; MMP-9-forward: 5′ TTGACAGCGACAAGAAGTGG 3′, MMP-9-reverse: 5′ GCCATTCACGTCGTCCTTAT 3′; and GAPDH-forward: 5′ AGGGCTGCTTTTAACTCTGGT 3′, GAPDH reverse: 5′CCCCACTTGATTTTGGAGGGA 3′.
Cell migration assay
The migration assay was performed by using Transwell insert chambers (8-μm pore size; Corning). About 2×105 cells were seeded into the upper chamber in serum-free medium in triplicate. The lower chamber was filled with 600 μL DMEM containing 10% FBS as chemoattractant. After incubation for 24 h, nonmigrating cells in upper chambers were removed by cotton swab, and cells migrating to the lower surface of membrane were fixed using methanol and stained with 0.1% crystal violet. The migrating cells were counted at least six visual fields per membrane under the light microscope.
Statistical analysis
Results were shown as mean and standard deviation (SD) from at least three independent experiments. Student's t-test was used and difference was considered significant when p-value was <0.05.
Results
HBx elevates AEG-1 expression without altering AEG-1 mRNA level
Previous studies found that AEG-1 expression levels increased from normal liver tissue to hepatitis B and HBV-related HCC samples (Gong et al., 2012), which implicated that HBV may be involved in controlling AEG-1 expression. Given HBx encoded by HBV is critical for HCC progression and HBx can enhance the levels of some oncoproteins (Kalra and Kumar, 2006; Liu et al., 2012). We wondered whether HBx could regulate AEG-1. To test this, increasing amounts of pCMV-HBx plasmids were transfected into HCC cell line SMMC-7721, and western blots were performed to detect AEG-1 protein level. As shown in Figure 1A, the level of AEG-1 protein was elevated with HBx expression in a dose-dependent manner. To address whether AEG-1 protein upregulation reflected changes in its mRNA level, we performed qRT-PCR analysis and found that the relative mRNA level of AEG-1 was not affected by HBx (Fig. 1B). To further confirm the effect of HBx on AEG-1 expression, AEG-1 protein levels and relative mRNA changes were also detected in HepG2 cells with stable HBx expression. The results indicated that AEG-1 protein level was significantly higher in HepG2-HBx than in HepG2-Ctr cells (Fig. 1C), but again the AEG-1 mRNA level had no obvious change (Fig. 1D). Then, two siRNA oligos were employed to deplete HBx and western blot indicated that AEG-1 protein level reduced with HBx knockdown (Fig. 1E). Next, we determined whether HBx-induced AEG-1 elevation was achieved by increasing AEG-1 protein stability. Cells were treated with cycloheximide (CHX) to block de novo protein synthesis, and AEG-1 protein level was examined at the indicated interval. As shown in Figure 1F, HBx expression had no significant effect on AEG-1 protein turnover. These findings suggest that HBx can upregulate AEG-1 expression via the post-transcriptional regulation.
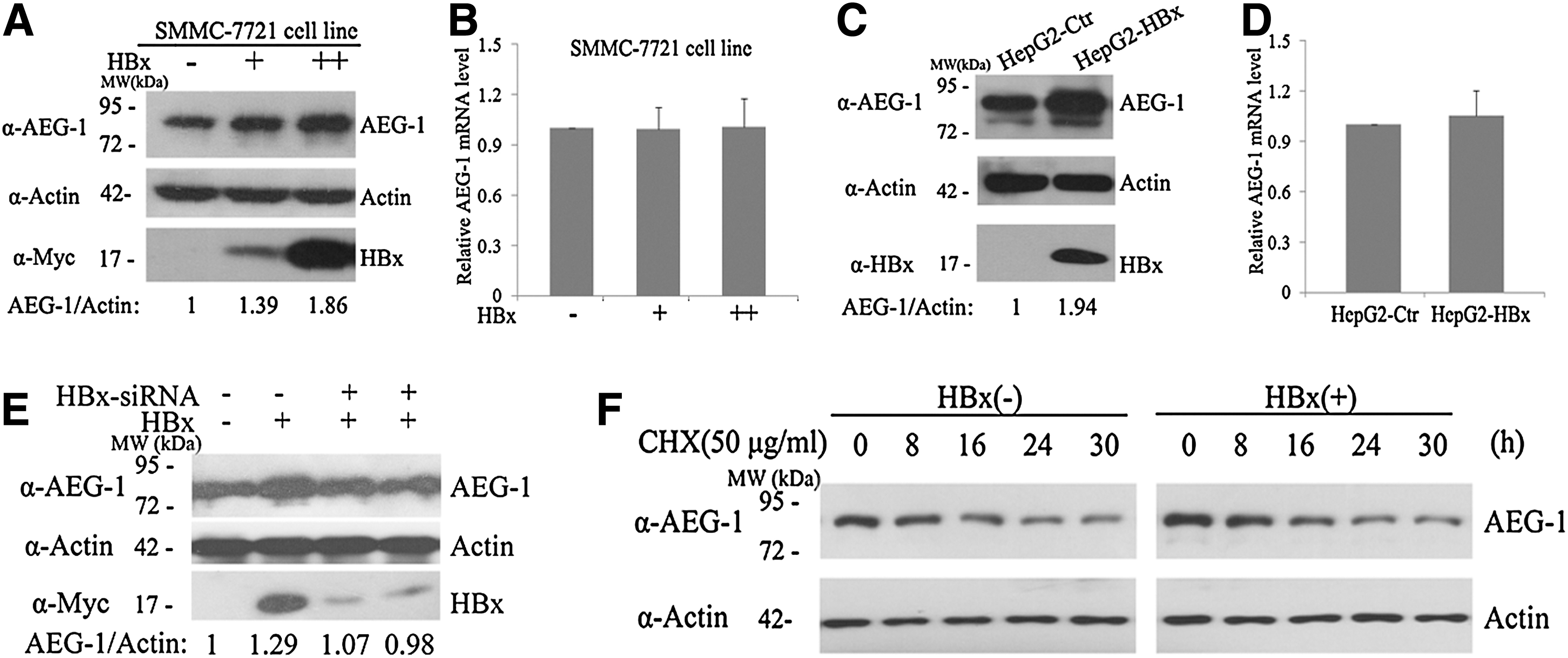
Hepatitis B viral X (HBx) elevates astrocyte elevated gene-1 (AEG-1) expression without altering AEG-1 mRNA level.
HBx-induced AEG-1 elevation is involved in HBx-mediated cell migration
The two siRNA oligos targeting AEG-1 were introduced into SMMC-7721 cells; qRT-PCR and western blot showed that they could markedly decrease AEG-1 mRNA level to about 70% and reduce AEG-1 protein expression to nearly 50% (Fig. 2A). However, the siRNAs were not successfully introduced into HepG2 cells because of the poor transfection efficiency. In addition, when SMMC-7721 cells were cotransfected with pCMV-HBx plasmid and AEG-1 siRNAs, the HBx-induced AEG-1 protein elevation was attenuated effectively (Fig. 2B). Since both HBx and AEG-1 were reported to enhance cell migration via upregulating MMP-9 (Chung et al., 2004; Liu et al., 2010, 2012; Sun et al., 2012), we detected the changes of MMP-9. Our result indicated that both MMP-9 mRNA and protein levels were increased with HBx expression. However, when pCMV-HBx plasmids were cotransfected with AEG-1 siRNAs, the HBx-mediated MMP-9 mRNA and protein elevation was blocked (Fig. 2C, D). Then, we detected whether AEG-1 participated in HBx-mediated cell motility with transwell assay. As shown in Figure 2E and F, there were more migrated cells in SMMC-7721 cells with HBx expression than in controls. In contrast, when decreasing AEG-1 expression with siRNA in the HBx-transfected cells, the HBx-mediated cell migrations were significantly reduced. These results implicate that the HBx-induced AEG-1 elevation is involved in HBx-mediated MMP-9 elevation and cell migration.
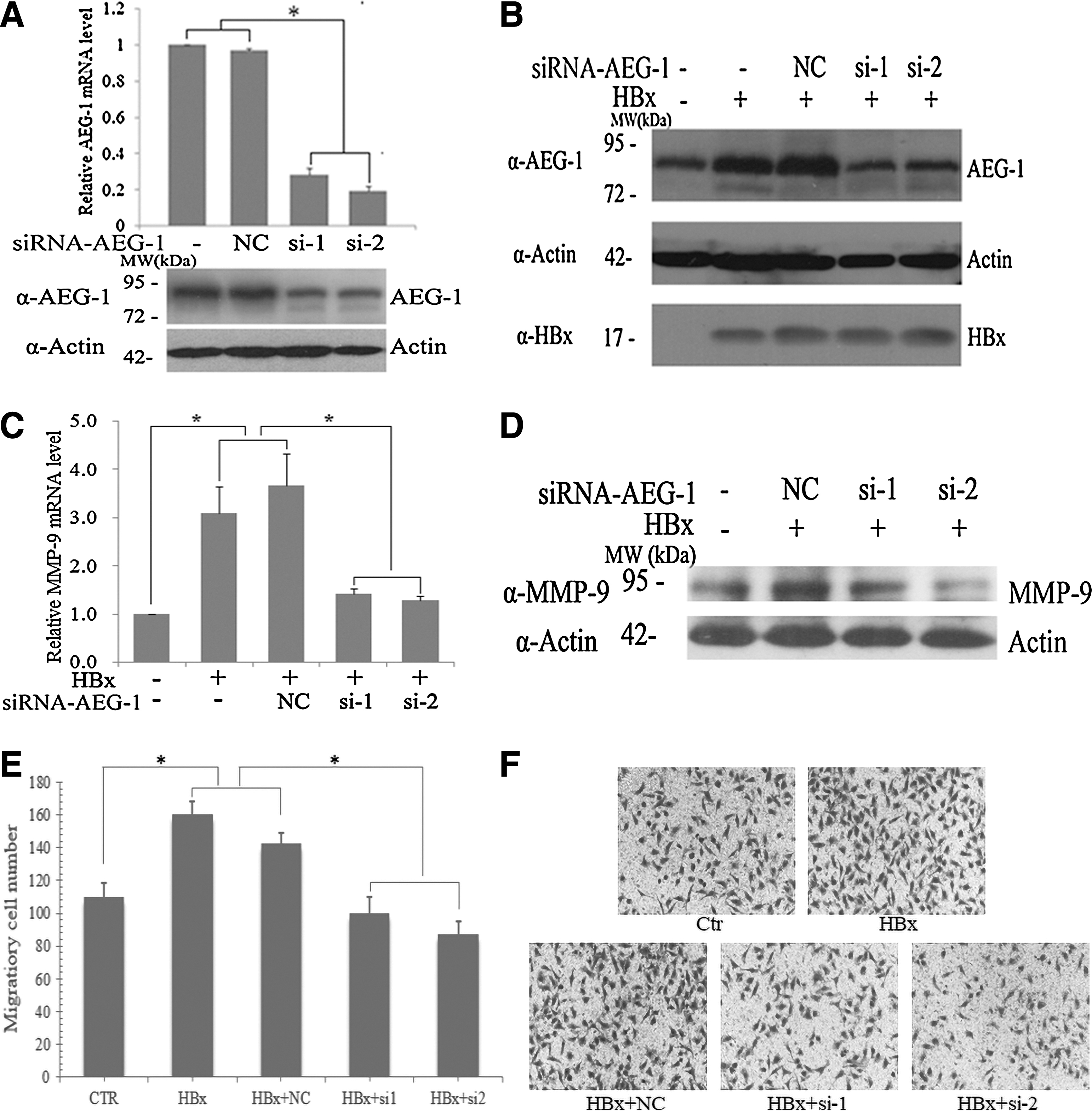
HBx-induced AEG-1 elevation is involved in HBx-mediated cell migration.
*
HBx increases AEG-1 expression by downregulating miR-375 and miR-136
Recent studies have manifested that miR-375 (Fan et al., 2012), miR-136 (Yang et al., 2012), and miR-26a (Zhang et al., 2011) could bind directly to the 3′-untranslated region of AEG-1 and suppressed cell growth, migration, or apoptosis via decreasing AEG-1 expression. In addition, HBx has been found to regulate miRNA expression in previous studies (Qiu et al., 2013). To further explore the underlying mechanism of HBx-mediated AEG-1 elevation, we first assessed whether HBx has any influence on the expression of miR-375, miR-136, and miR-26a by qRT-PCR. The results showed that miR-375 and miR-136 expression was significantly downregulated in SMMC-7721 cells that expressed HBx (Fig. 3A) and in HepG2-HBx cells (Fig. 3B), but miR-26a level did not change in SMMC-7721 and HepG2 cells expressing HBx (Fig. 3A, B). We also confirmed that miR-136 and miR-375 mimic introduction could reduce AEG-1 protein levels in SMMC-7721 cells (Fig. 3C). When the miR-136 and miR-375 mimics were cotransfected with pCMV-HBx into SMMC-7721 cells, we found that HBx-mediated AEG-1 protein upregulation was abrogated (Fig. 3D). Meanwhile, AEG-1 mRNA levels were not changed significantly with the high-dosed miR-136 and miR-375 mimic introduction (Fig. 3E), which was consistent with the result that HBx had no effect on AEG-1 transcription as shown in Figure 1B and D. These results indicate that HBx may increase AEG-1 protein level by downregulating miR-375 and miR-136 expression.
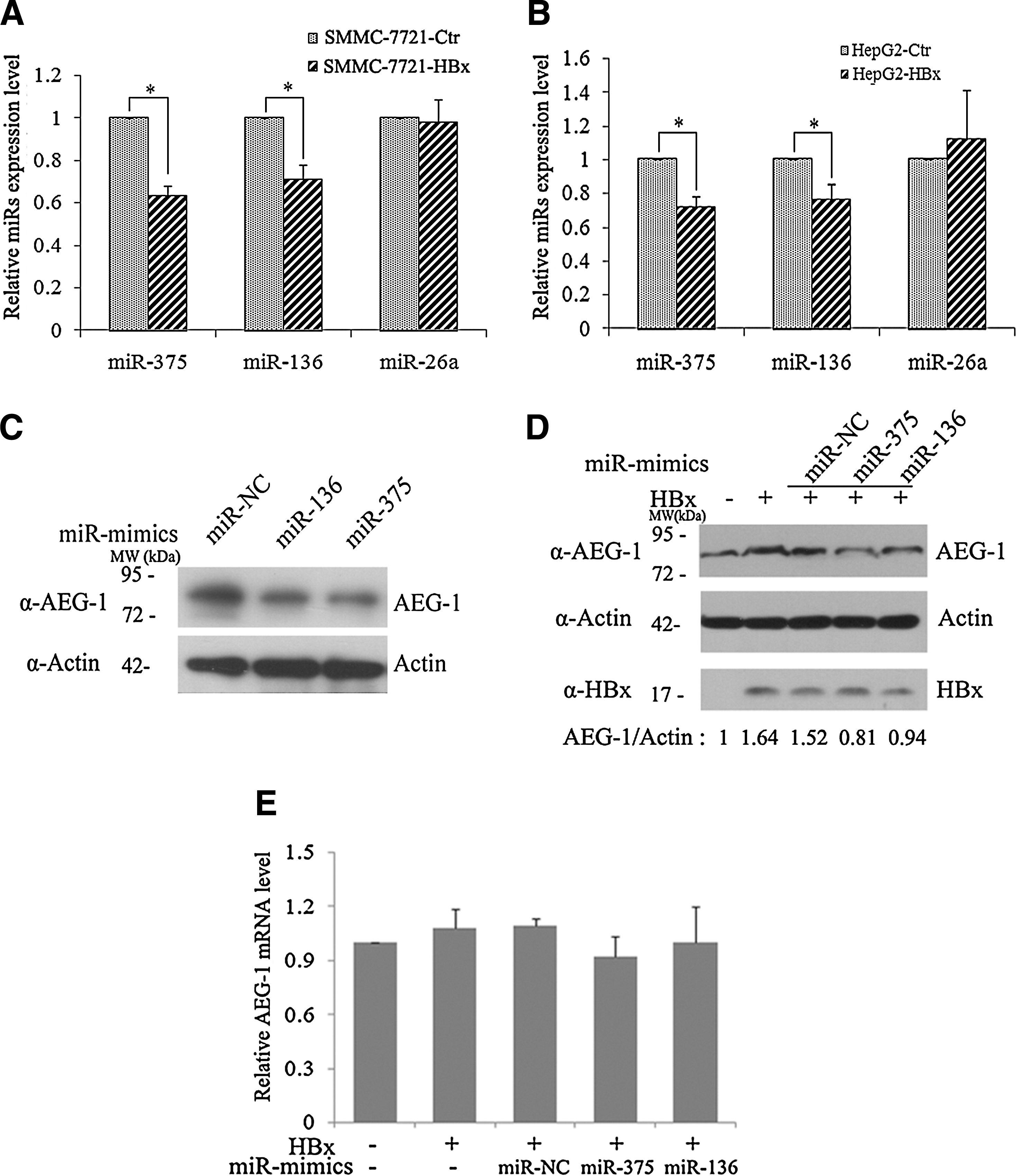
HBx increases AEG-1 expression by downregulating miR-375 and miR-136.
The introduction of miR-375 and miR-136 mimics attenuates HBx-mediated MMP-9 elevation and cell migration
To determine whether the restoration of miR-375 and miR-136 had any effect on HBx-induced cell migration, miR-375 and miR-136 mimics were re-expressed when SMMC-7721 cells were transfected with pCMV-HBx. The qRT-PCR results indicated that the HBx-induced MMP-9 mRNA level elevation was attenuated upon introducing miR-375 and miR-136 mimics (Fig. 4A). Cell migration assay was performed by transwell detection. The results showed that miR-375 and miR-136 restoration could partially suppress HBx-induced migration compared with control (Fig. 4B, C). These findings suggest that HBx upregulates AEG-1 protein level to promote cell migration by decreasing miR-375 and miR-136 expression.

miR-375 and miR-136 mimics attenuate HBx-mediated MMP-9 elevation and cell migration.
Discussion
In this study, we found that HBx overexpression could elevate AEG-1 protein level without affecting AEG-1 mRNA transcription. Similar results were also observed in the regulation of HBx on AIB1 (Liu et al., 2012). Further study revealed that HBx could interact with AIB1 protein and protected AIB1 from ubiquitin-proteasomal degradation. However, we found that HBx had no obvious effect on AEG-1 stability. When treated with CHX to block novel protein synthesis, HBx expression did not prolong AEG-1 half-life (Fig. 1F). Previous studies might provide some clues to this result. AEG-1 was not subjected to degradation through polyubiquitin-proteasome pathway (Thirkettle et al., 2009) and it was the longevous protein with half-life about 20 h (Lee et al., 2006). Therefore, it implies that except for inhibiting oncoproteins from polyubiquitinated degradation, there exists a novel mechanism for HBx to upregulate AEG-1 protein level.
Recently, miRNAs have been extensively investigated, which function to repress gene expression by base-pair complementation with the 3′-untranslated region (3′-UTR) of target mRNAs (Bartel, 2004). When the association between the miRNA and target mRNA is complete, the mRNA would be cleaved and direct degradation. Since most complementary pairs are imperfect, miRNA-mediated silence is frequently accomplished by translational suppression (Bartel, 2009). Aberrant miRNA expression has been implicated in numerous cancers, including HCC (Murakami et al., 2006). Many studies have evaluated the potential effects of HBx on miRNA dysregulations (Wang et al., 2010; Wei et al., 2013). HBx could decrease miR-101; consequently, the miR-101-target DNA methyltransferase 3A (DNMT3A) was upregulated, which induced aberrant DNA methylation and expression inhibition of some tumor suppressors, including GSTP1, CDKN2A, and APC (Wei et al., 2013). Currently, some microRNAs have been identified to target AEG-1 and block its oncogenic roles, such as miR-375 (Fan et al., 2012; Yang et al., 2012), miR-136 (Yang et al., 2012), and miR-26a (Zhang et al., 2011). Our results found that HBx could inhibit miR-375 and miR-136 expression to elevate AEG-1 protein level (Fig. 3). The restoration of miR-375 and miR-136 mimics not only attenuated HBx-induced AEG-1 elevation (Fig. 3D) but also repressed HBx-mediated cell migration (Fig. 4). MiR-375 has potential tumor-suppressive function, and miR-375 downregulation is established in HCC (Furuta et al., 2010; Fan et al., 2012) and the head and neck squamous cell carcinoma (HNSCC) (Hui et al., 2011). The hypermethylation within the promoter or coding regions of miR-375 may account for its suppression (Furuta et al., 2010; Hui et al., 2011). Then, whether the miR-101 downregulated by HBx is involved in HBx-mediated miR-375 or miR-136 reduction through DNMT3A pathway needs further investigation.
MMP-9 can facilitate the degradation of environmental barriers, such as extracellular matrix and basement membranes, which has important roles in the migration and invasion of tumors (Woessner, 1991). Our findings indicated that both MMP-9 mRNA and protein levels, and cell migration were increased with HBx overexpression. In contrast, when AEG-1 was knocked down by siRNAs, or miR-375 and miR-136 mimics were introduced in HBx-expressing HCC cells, both the HBx-induced MMP-9 upregulation and cell migratory were repressed significantly (Figs. 2 and 4). MMP-9 promoter region has the AP-1 and NF-κB binding sites (Sato and Seiki, 1993). HBx was reported to stimulate MMP-9 expression mainly through activating ERK and PI3K-Akt/PKB signaling cascades to promote AP-1 and NF-κB transcriptional activity (Chung et al., 2004). Moreover, AEG-1 overexpression was also found to positively correlate with MMP-9 elevation in gliomas and in nonsmall cell lung cancer with aggressive clinicopathologic outcomes (Liu et al., 2010; Sun et al., 2012). The interaction between AEG-1 and p50/p65 complex of NF-κB could stimulate the nuclear translocation of NF-κB to enhance MMP-9 expression (Sarkar et al., 2008). Study in gliomas also illustrated that AEG-1 was able to physically bind MMP-9 promoter region (from −690 nt to −483 nt, from −164 nt to −3 nt) cooperating with NF-κB p65 and/or AP-1 (Liu et al., 2010). In addition, there are many crosstalk and overlaps in regulating some oncogenic signaling networks between HBx and AEG-1, and the precise mechanisms by which HBx and AEG-1 control MMP-9 expression and cell migration remain to be determined.
Conclusions
In summary, our results demonstrated that HBx could elevate AEG-1 protein level by inhibiting miR-375 and miR-136 expression, and the HBx-induced AEG-1 upregulation enhanced HBx-mediated cell migration. These findings provide a novel mechanism for AEG-1 upregulation in HCC and shed new light on HBx promoting HCC development and progression. Additionally, our results also implicate the therapeutic potential of miR-375 and miR-136 in HBV-infected HCC.
Footnotes
Acknowledgments
The present study was supported by the Natural Science Foundation of China (81201521, 81373064, and 30972635), China Postdoctoral Science Foundation Funded Project (2013 M541467), the Natural Science Foundation of Shanghai Municipal Science and Technology Commission (12ZR1441500 and 12ZR1404300), and the Key Technologies Research and Development Program for Infectious Diseases of China (2013ZX10003001-002).
Disclosure Statement
No competing financial interests exist.