
Abstract
Mood stabilizers lithium and valproates are widely used in the treatment of bipolar disorder. It has been shown that these drugs can affect the inositol monophosphatase activity and thus the inositol de novo biosynthesis. However, the molecular mechanism of this action has thus far been vague. As such, characterizing the regulation of the gene encoding inositol monophosphatase at the molecular level can help to understand the bipolar disorder. As the model organism, the inositol monophosphatase is encoded by INM1 in Saccharomyces cerevisiae. In this study, we showed, using real-time reverse transcriptase polymerase chain reaction analysis, that INM1 is expressed in the presence of inositol, suggesting that the presence of inositol is required for INM1 transcriptional activation. We also demonstrated, using chromatin immunoprecipitation, that Ino2p is present at the promoter under uninduced conditions. Upon induction, Ino2p dissociates from the INM1 promoter. Furthermore, chromatin remodelers Ino80p and Snf2p are recruited to INM1 promoter upon induction as well as histone acetylases Gcn5p and Esa1p. Altogether, we have provided the evidence which describes how the transcriptional activator and coactivators participate in INM1 activation.
Introduction
B
Mood stabilizers play a very important role in the treatment of bipolar disorder. Two conventional anti-bipolar drugs, lithium and valproic acid (VPA) have been widely used in the treatment of bipolar disorder. However, the response rate of monotherapy with these drugs can only yield the optimum results in about one-third of the patients (Rybakowski et al., 2001; Gould et al., 2007). Because of such poor response rates, research into bipolar disorder remains a hugely complex undertaking.
It has been demonstrated that lithium treatment can regulate the attenuation of inositol signaling and phosphoinositol signaling (Berridge et al., 1989; King et al., 2009). The therapeutic role of VPA has also been associated with the reduction of de novo inositol synthesis and phosphoinositol depletion (Berridge et al., 1989; Agam et al., 2002; Eickholt et al., 2005; Shaltiel et al., 2007; Shimshoni et al., 2007). As such, it has been proposed that inhibition of inositol monophosphatase (IMPase) by lithium results in depletion of brain inositol, an increase in inositol-3-phosphate, downregulation of the phosphoinositide signaling cycle, and subsequent dampening of overactive neurotransmission (Berridge, 1985; Berridge et al., 1989). However, little is known about the molecular mechanism of this regulation. Therefore, it is imperative to identify and characterize the changes in cell signaling caused by the lithium and VPA treatments. Furthermore, the efficacy of lithium and VPA treatments can be understood through the study of the molecular mechanism of the therapeutic actions. As a result, we can understand the cause of bipolar disorder.
As an excellent model system, Saccharomyces cerevisiae has been used to study the molecular mechanism of inositol biosynthesis mediated by the VPA and lithium therapeutic effect. It has been reported that VPA, like lithium, causes inositol depletion in yeast (Vaden et al., 2001). The depletion of inositol might suggest the decrease in activity of IMPase, which is encoded by INM1 in yeast. IMPase catalyze the conversion of inositol-1-phosphate to inositol. Although the regulation of INM1 has been studied at the cellular level to evaluate the effect of lithium and VPA in INM1 expression (Murray and Greenberg, 2000; Agam et al., 2002; Shaldubina et al., 2002; Shamir et al., 2003), the molecular mechanism of INM1 expression has not been established. As such, we were interested in understanding the molecular mechanism of INM1 expression. In this study, we performed biochemical analysis to examine how the transcriptional activator and transcriptional coactivators are involved in INM1 activation.
Materials and Methods
Yeast strains and growth conditions
Both wild type (WT; INO2-FLAG; MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 ino2Δ::pESC-URA-INO2FLAG) and ino2Δ (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 ino2Δ) yeast strains were used in this study. The pESC-URA-INO2FLAG PLASMID was constructed as described below. The INO2 ORF was cloned into NotI and SpeI sites of pESC-URA plasmid so the INO2 ORF carries the C-terminal FLAG (#217454; Agilent). The resulting pESC-URA-INO2FLAG was confirmed by the sequencing. Subsequently, the pESC-URA-INO2FLAG was introduced into ino2Δ strain (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 ino2Δ) and the resulting WT (INO2-FLAG) strain was confirmed by Southern blot analysis and polymerase chain reaction (PCR). The yeast culture was grown at 30°C in either synthetic complete media without uracil (SC-ura, #114410622; MPBIO) or synthetic complete media (SC, #114400022; MPBIO), containing 2% glucose (w/v) with 100 μM myo-inositol or without myo-inositol for WT (INO2-FLAG) and ino2Δ cells, respectively.
RNA preparation and real-time reverse transcriptase PCR analysis
The total RNA preparation, first strand cDNA synthesis and real-time reverse transcriptase PCR (qRT-PCR) analysis was performed as described previously (Esposito et al., 2010; Wimalarathna et al., 2011, 2012). Briefly, 250 mL cells were harvested and resuspended in 400 μL lysis solution (10 mM Tris HCl pH 7.5, 10 mM EDTA, 0.5% SDS) followed by acid phenol (pH 4.3) extraction. After 1 h incubation at 65°C, the mixture was subjected to centrifugation, and the aqueous phase was subjected to chloroform extraction, ethanol precipitation, and was resuspended in 50 μL DEPC-treated H2O.
Equal amounts (10 μg) of total RNA were treated with RNase-free DNase (cat#79254; Qiagen) at 37°C for 1 h, and purified by phenol/chloroform (3:1) extraction and ethanol precipitation. One microgram of pure RNA was used in the SYBR GreenER Two-Step qRT-PCR kit (cat#11765-100; Invitrogen) for the first strand cDNA synthesis and real-time PCR reaction preparation as described in the manufacturer's manual. All experiments were repeated three times, and in each experiment, PCR reactions were done in triplicate in a 7500 sequence detection system (Applied Biosystems). All qRT-PCR primers are listed in the Supplementary Materials and Methods (Supplementary Data are available online at
The target DNA sequence quantities were estimated from the threshold amplification cycle number (CT ) using the Sequence Detection System software (Applied Biosystems). Each DNA quantity was normalized to the ACT1 DNA quantity by taking the difference between each gene's CT and ACT1's CT value, which is a ΔCT value. Each relative RNA fold change was calculated with the following formula: 2(−ΔC T ) (Esposito et al., 2010; Wimalarathna et al., 2011, 2012).
ChIP and real-time PCR analysis
The preparation of cross-linked chromatin immunoprecipitation (IP) procedures, and the real-time PCR analyses are as described previously (Esposito et al., 2010; Wimalarathna et al., 2012). All real-time PCR primers, TaqMan MGB probe and the antibody used in the study are listed in the Supplementary Materials and Methods. Input DNA was obtained before IP, whereas mock DNA was obtained without antibody treatment. Mock DNA and all ChIP-derived DNA were precleared with Salmon Sperm DNA/Protein A agarose (50%) to reduce nonspecific background. All experiments were repeated three times, and in each experiment, PCR reactions were done in triplicate.
Target DNA sequence quantities were estimated as described below. A corrected CT was calculated by subtracting the CT value for the corresponding mock (negative control) from the CT value for the IP or for the input sample. Subsequently, the (IP/INPUT)% (normalized IP) was calculated for each sample as described below. First of all, the ΔCT value was calculated by subtracting the corrected CT value for the IP sample from the corrected CT value for the corresponding input DNA to normalize the differences in ChIP aliquots. Second, each IP quantity was then calculated with the following formula: 2(ΔC T ) ×100% to represent (IP/INPUT)×100% (normalized IP).
Results
INM1 is expressed in the presence of myo-inositol
To study how myo-inositol affects the INM1 expression, WT (INO2-FLAG) cells were grown at 30°C in SC-ura (synthetic complete media without uracil) containing 2% glucose (w/v) and 10 μM inositol. When the optical density reached 0.9–1.1, cells were harvested and washed twice with SC-ura-ino to remove inositol completely. Subsequently, one half of cells were resuspended in SC-ura with 100 μM inositol and incubated at 30°C for 2 h. On the other hand, the other half of cells were resuspended in SC-ura-ino and incubated at 30°C for 2 h. Subsequently, cells were collected by centrifugation for total RNA preparation followed by qRT-PCR analysis. Our results showed that the relative ratios of INM1/ACT1 were 0.08 and 0.55 under the absence or the presence of inositol, respectively (Fig. 1). The INM1 expression increased significantly, which is about seven-fold in the presence of 100 μM inositol. This observation suggests that INM1 is expressed in the presence of inositol. As such, INM1 is uninduced in the absence of inositol and it is induced in the presence of inositol.
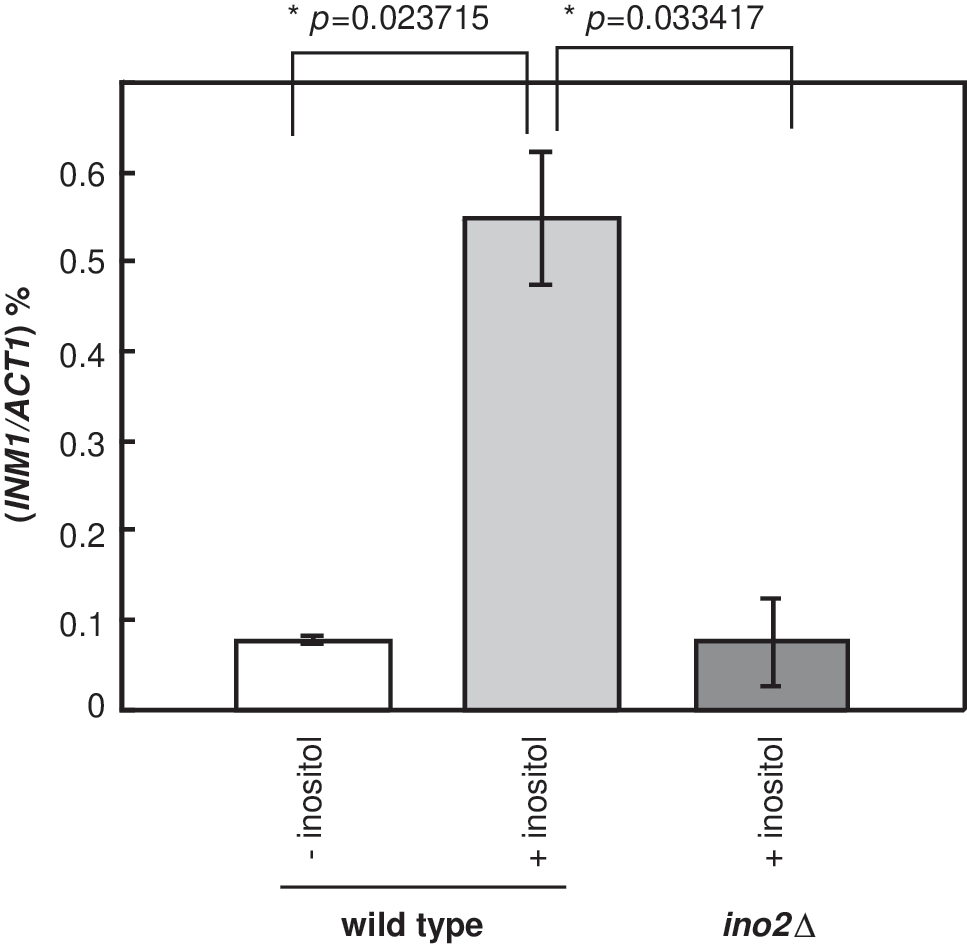
INM1 is expressed in the presence of inositol. INM1 and ACT1 mRNA were detected by qRT-PCR with INO1 and ACT1 probes, respectively. Mid-log phase WT (INO2-FLAG) cells or ino2Δ cells were grown 2 h at 30°C in synthetic complete media without uracil (SC-ura) or synthetic complete media (SC) containing 2% glucose (w/v) with 100 μM myo-inositol or without myo-inositol, respectively. qRT-PCR, real-time reverse transcriptase polymerase chain reaction; WT, wild type.
Previously, it was identified that INM1 promoter contains the UAS INO consensus sequences (Wimalarathna et al., 2012). It has also been demonstrated that the transcriptional activator Ino2p is required to bind to the UAS INO element for transcriptional activation of UAS INO -containing genes (Schwank et al., 1995). It is instructed to examine whether Ino2p plays a role in INM1 expression. In this study, the ino2Δ cells were grown in the presence of inositol. Our results showed that the expression levels of INM1 were similar to the WT (INO2-FLAG) cells in the absence of inositol, suggesting that INM1 is not expressed under the inducing conditions in the absence of Ino2p (Fig. 1). Therefore, INM1 induction was inhibited in ino2Δ cells, suggesting that Ino2p plays an important role in INM1 transcriptional activation.
Ino2p departs from the INM1 promoter upon induction
As INM1 expression was inhibited in the absence of Ino2p, we are interested in examining whether Ino2p regulates INM1 expression directly. ChIP coupled with real-time PCR (qPCR) analysis was performed. WT (INO2-FLAG) cells were grown in SC-ura until mid-log phase. For inducing conditions, cells were harvested and washed twice with a complete synthetic medium without inositol (SC-ura-ino) to remove inositol completely. Subsequently, cells were resuspended in SC-ura with 100 μM inositol and incubated at 30°C for 2 h. For repressing conditions, cells were resuspended in SC-ura-ino and incubated at 30°C for 2 h. Subsequently, cells were fixed and immunoprecipitated with antibodies against FLAG followed by qPCR analysis.
The IP signals were observed at the INM1 upstream activating sequences (UAS). The relative IP value represented the ratio of INM1 promoter IP DNA normalized to INM1 input DNA (Esposito et al., 2010; Konarzewska et al., 2012; Wimalarathna et al., 2012). Our results showed that Ino2p IP value was 1.7% under uninduced conditions (Fig. 2). The Ino2p IP value decreased to 0.7% under induced conditions. The decrease is more than 50% and is significant. As a control, we also examined the presence of Ino2p IP at the INO1 promoter, and our results showed that the quantity of Ino2p remained constant at the INO1 under both conditions (Fig. 2). The result that Ino2p presents at the INO1 promoter constitutively is consistent with previous observations (Brickner and Walter, 2004; Dasgupta et al., 2005; Ford et al., 2007; Ford et al., 2008). As such, our result suggests that Ino2p is present at the INM1 promoter under uninduced conditions, and it departs from the promoter after induction.
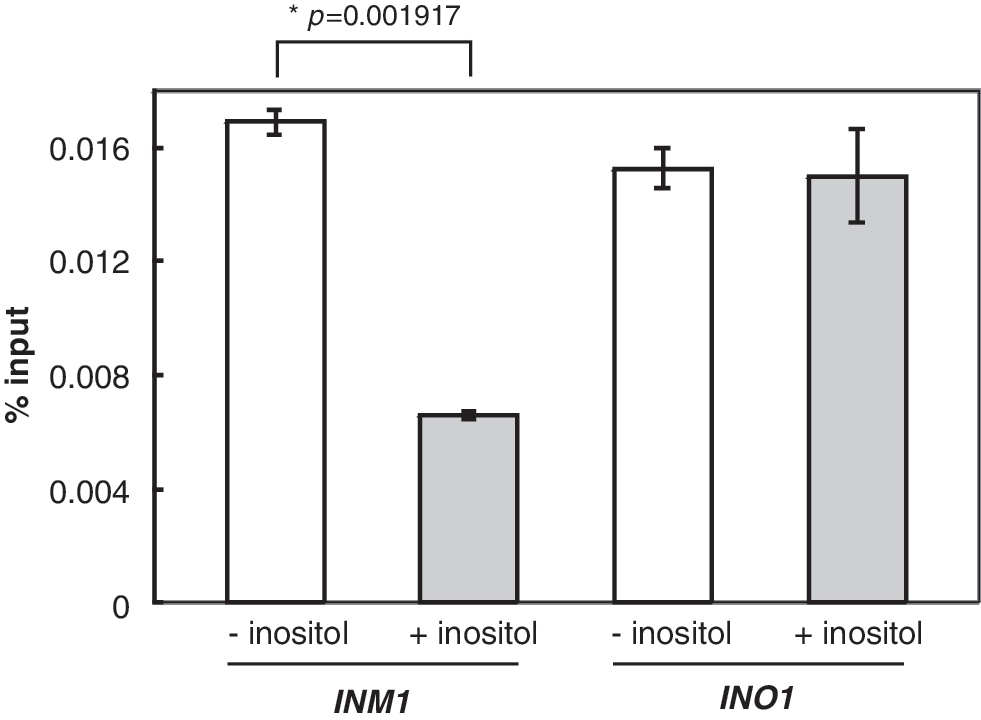
Ino2p departs from the INM1 promoter in induction. Real-time PCR of DNA immunoprecipitated with antibodies against Ino2p-FLAG (α-FLAG) at INM1 promoter and INO1 promoter. All experiments were repeated at least three times and, in each experiment, PCR reactions were done in triplicate. The % of input is graphed as mean±standard deviation.
The recruitment of chromatin remodelers in INM1 induction
It has been shown that both Ino80p and Snf2p are directly involved in INO1 expression at the transcriptional levels (Ford et al., 2007; Ford et al., 2008; Konarzewska et al., 2012). As one of the UASINO -containing genes, it is instructed to examine whether these two chromatin remodelers also participate in the INM1 gene expression. ChIP coupled with qPCR analysis was again performed here. Antibodies against both Arp8p subunit of INO80 (Arp8-IP) and Snf2p subunit of SWI/SNF (Snf2-IP) were used in the ChIP. The relative IP value represented the ratio of promoter IP DNA normalized to input DNA. Our results showed that the quantity of Arp8-IP is 1.3% and 2.5% for uninduced and induced WT (INO2-FLAG) cells, respectively (Fig. 3). The increase of INO80 upon induction is about two-folds. For Snf2-IP, the quantity of Snf2p is 0.6% and 1.8% for uninduced and induced WT (INO2-FLAG) cells, respectively. The increase of Snf2p is more than two-folds upon induction. As such, our results demonstrated that both Ino80p and Snf2p remodelers are recruited to INM1 promoter upon induction and the presence of both remodelers at the INM1 promoter suggests that they are directly involved in INM1 expression.
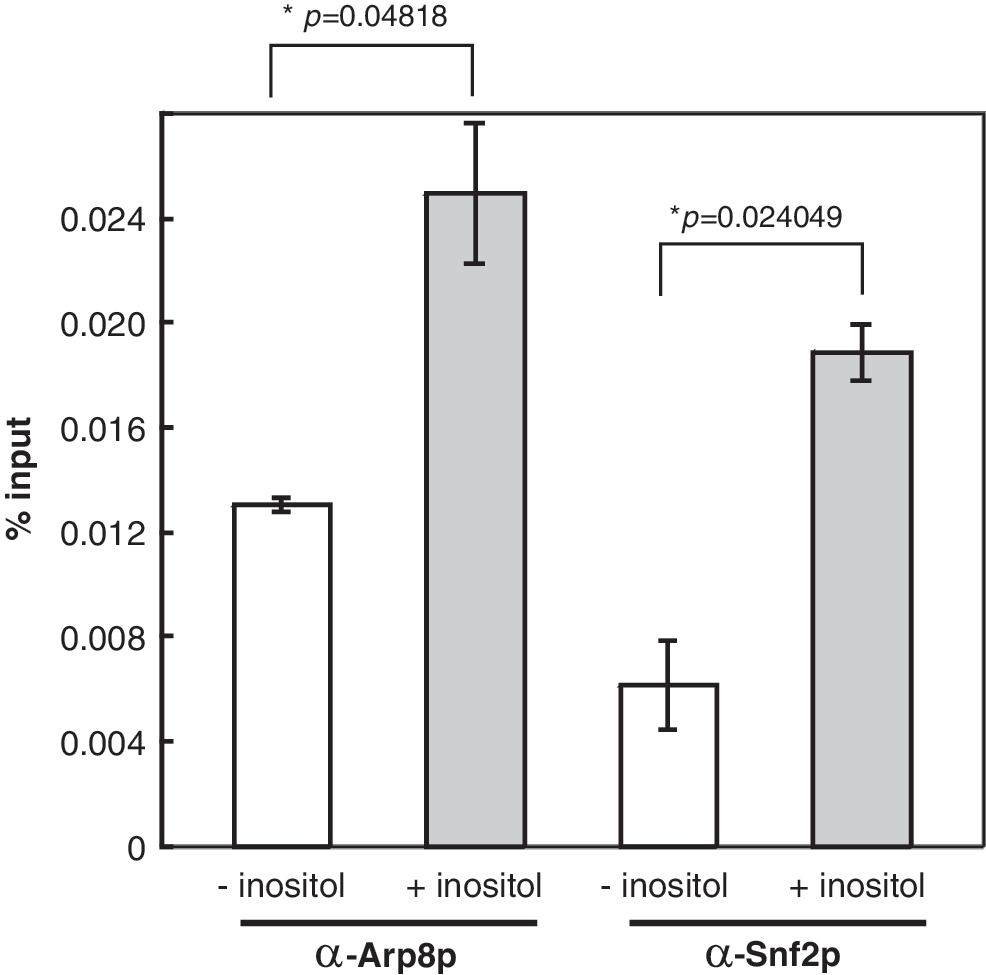
Both INO80 and SWI/SNF chromatin remodelers are recruited to the INM1 promoter in induction. Real-time PCR of DNA immunoprecipitated with antibodies against INO80 (α-Arp8p) and SWI/SNF (α-Snf2p) at INM1 promoter. Annotation was as described in the legend of Figure 2.
Both Gcn5p and Esa1p histone acetylases are directly involved in INM1 induction
It has been demonstrated that both Gcn5p and Esa1p are responsible for histone acetylation during INO1 activation (Esposito et al., 2010; Konarzewska et al., 2012). To examine whether these histone acetylases are also recruited to the INM1 promoter, WT (INO2-FLAG) strain was used in the ChIP analysis. Antibodies against both Gcn5p (Gcn5-IP) and Esa1p (Esa1-IP) were used in the ChIP. Results showed that the relative IP values for Gcn5-IP were 2.3% and 3.4% of input under repressing and inducing conditions, respectively (Fig. 4). These results suggested that Gcn5p is recruited to the INM1 promoter upon transcriptional activation. Similar results were also observed for Esa1p, which showed the relative IP values were 1.9% and 3.0% of input under repressing and inducing conditions, respectively (Fig. 4). As such, both histone acetylases are recruited to the INM1 promoter during induction.

Both Gcn5p and Esa1p histone acetylases are recruited to the INM1 promoter in induction. Real-time PCR of DNA immunoprecipitated with antibodies against Gcn5p (α-Gcn5p) and Esa1p (α-Esa1p) at INM1 promoter. Annotation was as described in the legend of Figure 2.
Discussion
RNA analysis was performed to understand how phospholipid precursor inositol regulates the INM1 expression. We observed that INM1 is expressed in the presence of inositol and it is repressed in the absence of inositol (Fig. 1). Previously, it has been shown that INO1 is repressed in the presence of inositol and it is derepressed in the absence of inositol (Ford et al., 2007; Ford et al., 2008; Wimalarathna et al., 2011). As such, the regulatory pathway of INM1 is completely opposite from INO1. Furthermore, our results showed that INM1 induction is inhibited in ino2Δ cells, suggesting that Ino2p is required for INM1 induction.
We have shown that the Ino2p is present at the INM1 promoter in the repressing conditions and a significant amount departs from the INM1 promoter in the inducing conditions (Fig. 2). It is possible that Ino2p brings in transcriptional coactivators under inducing conditions and then leave the promoter when they complete the activation process. As such, we observed less amount of Ino2p after induction. On the other hand, many studies have demonstrated that Ino2p expression is decreased in the presence of inositol (Ashburner and Lopes, 1995; Eiznhamer et al., 2001). As such, Ino2p occupancy is decreased under the INM1 inducing conditions because less Ino2p is available in the presence of inositol. Since we observed that Ino2p occupancy remains constant at the INO1 promoter, the lower Ino2p occupancy at INM1 promoter might be irrelevant to the Ino2p quantity.
It is possible that the departure of Ino2p from INM1 promoter is to prevent the binding of the repressor. Many studies have shown that Ino2p constitutively binds to INO1 promoter (Brickner and Walter, 2004; Dasgupta et al., 2005; Ford et al., 2007; Ford et al., 2008), and the regulation of INO1 expression depends on the presence of Opi1 repressor in INO1 promoter. The Opi1p is released from the endoplasmic reticulum (ER) in the presence of inositol. Subsequently, the Opi1 repressor enters the nucleus and interacts with Ino2p to repress INO1 expression. In the absence of inositol, the Opi1p is anchored by the phosphatidic acid on ER. At this time, Ino2p departs from the INM1 promoter upon inducing conditions, which is in the presence of inositol. Since Opi1p is released from ER and can interact with Ino2p in the presence of inositol, it is possible that the dissociation of Ino2p from INM1 promoter is to prevent the negative effect of Opi1p on INM1 expression.
In an attempt to examine how chromatin remodelers are involved in INM1 expression, we found that INO80 and SWI/SNF were absent from the INM1 promoter under uninduced conditions. Both INO80 and SWI/SNF were largely recruited to the INM1 promoter (Fig. 3). Although this recruitment pattern is different from INO1 expression, many studies have shown similar recruitment patterns, including ARG1, CUP1, and GAL1 (Bryant and Ptashne, 2003; Govind et al., 2005; Wimalarathna et al., 2014). Since we observed the departure of Ino2p from INM1 promoter upon induction, it is possible that the Ino2p recruits these remodelers before it departs from the promoter. However, our current system cannot examine this possibility. Further biochemical analysis such as kinetic ChIP assay might be able to identify whether Ino2p is required to recruit these remodelers.
Our data also showed that both histone acetylases Gcn5p and Esa1p are recruited to INM1 promoter upon induction. This observation indicates that both Gcn5p and Esa1p are involved in the INM1 induction. Recent studies have demonstrated that the recruitment of histone acetylases might not play an important role in the initial activation, but might be important for the departure of chromatin remodelers in the late stage of transcription (Konarzewska et al., 2012). It is not clear whether both histone acetylases are essential for the initial stage of INM1 activation. As such, further analysis in the recruitment dependency would help to clarify the issue.
In this study, we have defined the inducing conditions for INM expression and we have also examined the molecular mechanism of INM1 induction. It has been shown that the anti-bipolar drugs cause the inositol depletion in yeast (Vaden et al., 2001). In this study, we demonstrated that INM1 expression is inhibited in the absence of inositol, suggesting that INM1 expression is inhibited in the presence of anti-bipolar drugs. We also showed both chromatin remodelers and histone acetylases are recruited to INM1 promoter upon induction. However, further analysis is necessary to uncover the specific role of Ino2p and the interplay within the transcriptional activator, remodelers, and histone modifying enzymes. As such, we will be able to understand the molecular mechanism of INM1 expression, and understand how anti-bipolar drugs influence its mechanism, which will provide a key insight into the molecular mechanism of bipolar disease.
Footnotes
Acknowledgments
This work was supported by the Science and Technology Grant of the Department of Education, Liaoning, China (L2010657) to L.Z.
Disclosure Statement
There are no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.