
Abstract
Cholesterol is important for the growth and persistence of Mycobacterium tuberculosis in macrophages. The mce4 locus, which is conserved in both M. tuberculosis and Mycobacterium bovis, is thought to be responsible for cholesterol transport into the bacteria. However, the exact roles of specific genes within the sophisticated mce4 system remain poorly understood. In this study, Mce4A and Mce4E of M. bovis, two proteins that are encoded by the mce4 locus, were expressed in Mycobacterium smegmatis. The recombinant strain expressing the Mce4E protein (M. smeg::E) performed better than that expressing the Mce4A protein (M. smeg::A) in a minimal medium with and without glycerol or cholesterol, which may be the reason why M. smeg::E showed better survival in ANA-1 macrophages than did M. smeg::A. Cytokine expression profiles were similar in macrophages infected with either recombinant strain. We also investigated the role of CD36 in recognizing Mce4A and Mce4E proteins. However, CD36 did not appear to be specific for these proteins and showed little impact on the ultimate clearance of the recombinant strains. Reduced interleukin-1β, inducible nitric oxide synthase, and tumor necrosis factor-alpha mRNA expression at 6 h postinfection in macrophages infected with M. smeg::E was observed using a CD36-specific monocular antibody to block the receptor, whereas no obvious changes in the expression of these cytokines were observed in cells infected with M. smeg::A with or without exposure to the CD36 antibody. Conclusively, the different performances of the recombinant strains suggest that the Mce4A and Mce4E proteins enhance mycobacterial adaptation to the harsh environment within macrophages after phagocytosis.
Introduction
M
Some Mce proteins are thought to play a role in pathogenesis and immunity. Knockout mutations in the mce1, mce2, and mce3 operons in M. tuberculosis attenuated virulence (Gioffre et al., 2005). The mce2 and mce3 mutants induced less tissue damage (pneumonia) and with lower bacterial numbers (Gioffre et al., 2005). Expression of mce1 in M. tuberculosis has been demonstrated by both the antibody and real time-polymerase chain reaction (RT-PCR) in vitro and during natural infection in man (Ahmad et al., 1999). Levels of expression differed at different stages of growth and under varied culture conditions in vitro (Kumar et al., 2003, 2005).
Recent studies revealed that the mce4 system may be involved in the transport and metabolism of cholesterol. Mohn et al. (2008) found that the growth of Rhodococcus sp. strain RHA1 on cholesterol requires expression of the mce4 system and proposed that in M. tuberculosis it may serve a similar function. Microarray analysis revealed that all of the mce4 genes of cholesterol-fed RHA1 were upregulated (Van der Geize et al., 2007). The mce4 system was also identified in M. tuberculosis as an ABC-like transport system involved in cholesterol import with deletion of the operon resulting in a growth defect when cholesterol was the primary carbon source (Pandey and Sassetti, 2008).
The mce4 operon is essential for mycobacterial persistence in macrophages. However, the exact roles of specific genes in this sophisticated cholesterol import system remain poorly understood. In our previous studies, we demonstrated that recombinant Mce4A and Mce4E proteins exerted a profound impact on the host immune response by alteration of macrophage cytokine expression and the host inflammatory response (Xu et al., 2007, 2008).
M. tuberculosis mce4 homologs are present in M. bovis, Mycobacterium Avium, and Mycobacterium smegmatis. The Mce4 proteins in M. smegmatis (MSmce4) are almost identical in size as those in M. tuberculosis (MTmce4), although pairwise alignments show the MSmce4 proteins to be divergent from MTmce4 and MAmce4 (M. avium) (Haile et al., 2002). To study the function of Mce4A and Mce4E in M. bovis further, we cloned and expressed mce4A and mce4E from M. bovis in M. smegmatis and investigated the survival of the recombinant strains in ANA-1 murine macrophages. The relative mRNA expression levels of three cytokines, interleukin (IL)-1β, inducible nitric oxide synthase (iNOS), and tumor necrosis factor (TNF)-α, were measured by quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR) to evaluate the effects of the strains on macrophages. The results of our study conclusively demonstrate that differences in the expression profiles of Mce4A and Mce4E may be related not only to the gene organization but also to their involvement in adaptation to the harsh and changing conditions in the macrophages.
Materials and Methods
Bacterial strains and culture conditions
The M. bovis Beijing strain 93006, designated bovis 93006, a virulent strain, and M. smegmatis strain ATCC607, designated smeg ATCC607, were obtained from the China Institute of Veterinary Drug Control (CVCC, China) and cultured in Middlebrook 7H9 broth (Difco) supplemented with 10% albumin–dextrose–catalase (ADC) (vol/vol) and 0.05% Tween 80 (vol/vol) with shaking.
Cloning and expression of mce4A and mce4E
Genomic DNA from the bovis strain 93006 was extracted as described previously (Xu et al., 2007, 2008). Full length mce4A and mce4E genes were amplified from bovis strain 93006 genomic DNA by PCR. The primers and corresponding restriction enzyme sites used are listed in Table 1.
Restriction sites in primers are indicated in bold letters.
The amplicons were cloned in pmv261 (Stover et al., 1991) to generate pmv261/mce4A and pmv261/mce4E plasmids. M. smegmatis smeg ATCC607 was transformed with the recombinant plasmids by electroporation to generate M. smeg::A and M. smeg::E. The presence of the inserts was confirmed by restriction enzyme digestion and sequencing.
The fresh cultures were grown to OD600≈0.6, then heated at 45°C for 30 min, and maintained at 45°C with shaking for a further 1 h. Cells were harvested by centrifugation at 12,000 rpm for 10 min, washed twice, and resuspended in sterile phosphate-buffered saline (PBS).
To induce the expression of these proteins, the recombinant M. smeg::A and M. smeg::E strains were cultured in a shaking incubator in the LBGTW broth [5 g
Bacterial growth on cholesterol or glycerol
The minimal medium (MM) was used to assess bacterial growth on cholesterol or glycerol and contained 0.5 g asparagine, 1 g KH2PO4, 2.5 g Na2HPO4, 50 mg ferric ammonium citrate, 0.5 g MgSO4·7H2O, 0.5 mg CaCl2, 0.1 mg ZnSO4 per liter with 0.01% cholesterol (wt/vol) or 0.1% glycerol (vol/vol). Cholesterol was diluted in 1:1 (vol/vol) tyloxapol–ethanol before addition (Pandey and Sassetti, 2008). Bacterial growth in the MM was assessed by monitoring OD600. As control, the MM without cholesterol also contained 1:1 (vol/vol) tyloxapol–ethanol, whereas the MM without glycerol did not contain this combination of tyloxapol and ethanol.
For growth curves, bacterial cultures prepared as above were washed with PBS and diluted into MM to obtain a final optical density at 600 nm (OD600) of 0.1.
Cell infection with recombinant M. smegmatis strains
ANA-1 murine macrophages were obtained from the Committee on Type Culture Collection of the Chinese Academy of Sciences. Before infection, bacterial stocks were cultured as described above to induce recombinant protein expression. ANA-1 murine macrophages (2×105 cells/well in 24-well plate) were challenged with the recombinant strains (multiplicity of infection [MOI]=10:1) at 37°C in 5% CO2 for 2 h. To remove extracellular bacteria, 600 μL fresh warm PBS was added into each well and then removed, with this procedure repeated twice. Cells were incubated with fresh RPMI-1640 containing 2% fetal calf serum (FCS) and gentamicin (final concentration, 100 μg/mL) for 30 min, then washed twice with PBS, and incubated again with fresh RPMI-1640 containing 2% FCS for 6, 12, 24, 48, or 72 h. Macrophages challenged with M. smeg containing the empty plasmid pmv261 were treated using the same process as a control.
Effects of Mce4A and Mce4E on macrophage viability
ANA-1 macrophages (104 cells/well) were seeded in 96-well plates and infected as described above. Each sample was then incubated with 20 μL (5 mg/mL) MTT [3-(4,5-dimethylthiazol-2-y1)-2,5-diphenyltetrazolium bromide] solution. After 4 h of incubation, dimethyl sulfoxide (DMSO) was added to dissolve the formazan, and the samples were kept at 37°C for 10 min. Absorbance values were measured at 570 nm.
RNA extraction and cDNA preparation
ANA-1 macrophages were challenged as above, and total RNA extraction and transcription to cDNA were carried out for each sample using an RNA extraction kit (Aidlab) and the RevertAid first-strand cDNA synthesis kit (Fermentas) according to the manufacturer's protocol.
Real-time quantitative PCR
The expression levels of five genes (four selected genes and one control) were investigated using real-time quantitative PCR. The housekeeping gene β-actin was used as a control gene to normalize expression data for target genes. Gene expression levels were detected using the DNA Engine Opticon TM2 fluorescence detection system (MJ Research, Inc.) and SYBR Green (RealMasterMix; Tiangen). The specific gene primer pairs are listed in Table 2. Real-time quantitative PCR data were analyzed using the 2−ΔΔt method.
IL, interleukin; iNOS, inducible nitric oxide synthase; TNF, tumor necrosis factor.
Intracellular survival of recombinant M. smegmatis
The intracellular survival of recombinant M. smegmatis was assessed according to the number of colony-forming units (CFUs). At each time point, infected cells were lysed with 1% Triton X-100 solution, serially diluted in PBS, and plated onto 7H10 agar plates containing 10% ADC. CFUs were counted after incubation at 37°C for 3 days. One group of macrophages was stimulated with 100 U/mL murine IFN-γ for 24 h before infection, while the other was not stimulated.
Role of CD36 in recognition of Mce4A and Mce4E proteins
ANA-1 macrophages were incubated with the CD36 antibody for 1 h and then infected with either recombinant M. smegmatis strain, and the control group was infected without exposure to the CD36 antibody (GeneTex). Cytokine expression analysis and CFU assessment were conducted as described above.
All the experiments were repeated thrice. Differences between groups were assessed by one-way ANOVA and post hoc Turkey's test using SPSS 13.0 software. Data are expressed as mean±standard deviation. Differences with p<0.05 were considered statistically significant.
Results
Cloning and expression of Mce4A and Mce4E
The recombinant pmv261/mce4A and pmv261/mce4E plasmids were constructed and the inserts were confirmed by restriction enzyme digestion and sequencing. SDS-PAGE analysis confirmed the induction of the recombinant strains and the control (the result of SDS-PAGE analysis is shown in the Supplementary Fig. S1; Supplementary Data are available online at
M. smeg::A and M. smeg::E growth on cholesterol or glycerol
Growth of the two recombinant strains in the MM with or without 0.1% glycerol was assessed. Both recombinant strains experienced a logarithmic growth phase 24 h postinoculation and reached a stationary phase after 48 h. Beyond 96 h, the growth of M. smeg (control) and M. smeg::E dramatically slowed, whereas M. smeg::A did not experience a decline in growth until 120 h. Bacterial growth in the MM without a carbon source was observed as a control, and the MM without a carbon source did not support significant growth (Fig. 1A).
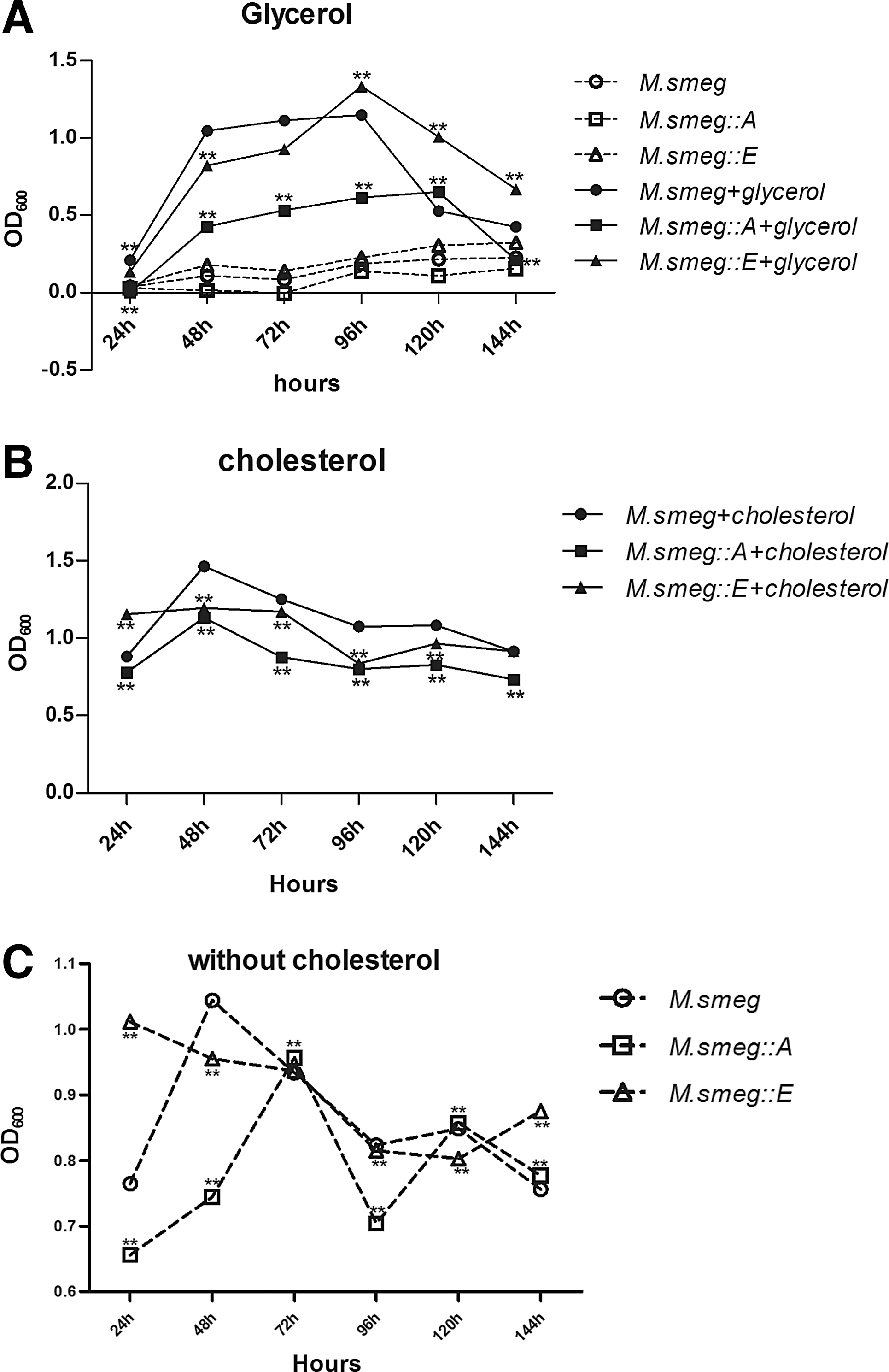
M. smeg::A and M. smeg::E growth status in cholesterol or glycerol. Bacterial growth status fed on cholesterol or glycerol. Bacterial growth in defined minimal medium containing 0.1% glycerol
The recombinant and control strains were cultured in the MM with or without 0.01% cholesterol, and the OD600 values of the cultures were recorded. For the cholesterol-fed strains, the OD600 values peaked at 48 h and then declined slowly (Fig. 1B). By contrast, the OD600 values of the strains kept in the MM without cholesterol changed dramatically during infection (Fig. 1C). The OD600 of M. smeg::E cultures was in a continuous decline until 120 h postinfection and then increased slightly until 144 h. The OD600 trends of M. smeg and M. smeg::A cultures were similar. For M. smeg and M. smeg::A, OD600 values peaked at 48 and 72 h, respectively, and then decreased progressively until 120 h when they increased again.
Effects of M. smeg::A and M. smeg::E on macrophage viability
The MTT assay was used to assess the effects of the M. smeg::A and M. smeg::E strains on ANA-1 macrophage viability. Infection with both recombinant strains increased cell viability for the first 24 h, but after 24 h, cell viability was reduced (Fig. 2). The mean values for each group at different time points are given in Table 3. The differences in viability between M. smeg::A- and M. smeg-infected cells at 12 h and between M. smeg::E- and M. smeg-infected cells at 48 h were significant (p<0.05).

Effects of M. smeg::A and M. smeg::E on the viability of ANA-1. MTT assay was used to assess the effects of the indicated strains on ANA-1 cell viability. The mean value of each treatment at different time points is shown in Table 3. Asterisk (*) indicates a statistical significance between the M. smeg and the recombinant strains. *p<0.05.
p<0.05.
Intracellular survival of M. smeg::A and M. smeg::E strains
Viable bacterial numbers as CFU assessment were used to assess the intracellular growth and survival of the two recombinant strains. Resting ANA-1 macrophages were infected with the recombinant strains at a MOI of 10:1, and CFUs were counted at different time points. The numbers of CFU of the strains were very low during early infection, but steadily increased. After 24 h, a rapid increase in the multiplication of bacteria was observed. The number of M. smeg::E CFUs was significantly increased 24 h after infection compared to the control (p<0.01). The number of M. smeg::A CFUs also was increased 24 h after infection, but the numbers of M. smeg::A and M. smeg CFUs did not differ significantly at 48 and 72 h (Fig. 3A).
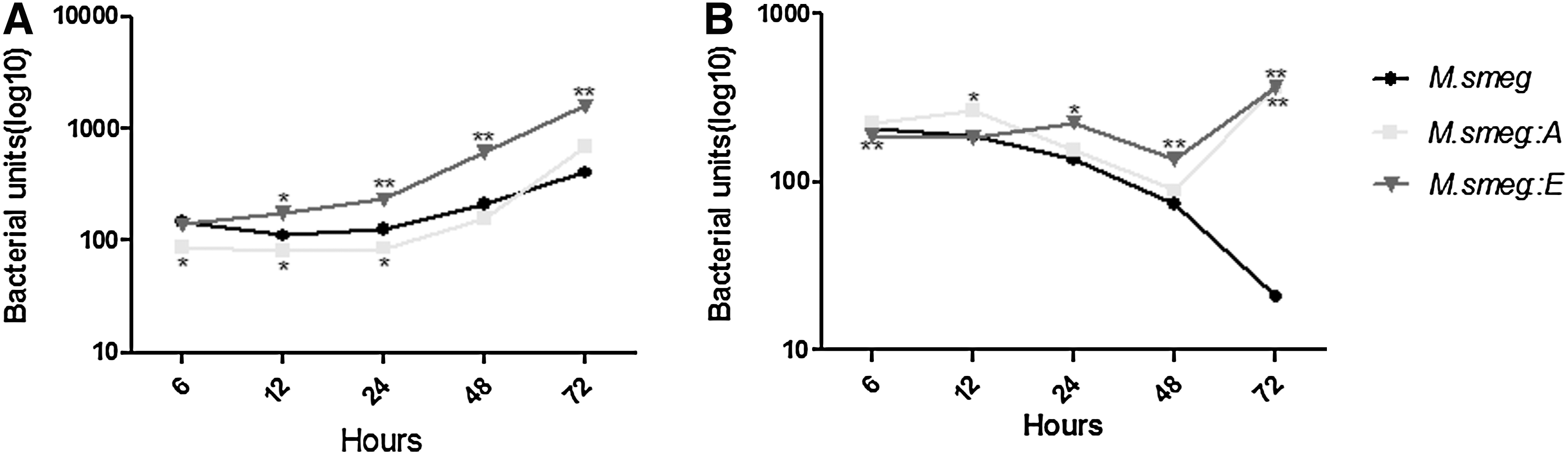
Intracellular survival of M. smeg::A and M. smeg::E. The ability of M. smeg::A and M. smeg::E to persist in resting macrophages
We also investigated the survival of the M. smeg::A and M. smeg::E strains in ANA-1 macrophages activated by 100 U/mL IFN-γ for 24 h before bacterial infection. During the infection, the CFU count of the control M. smeg continuously decreased, starting at ∼200 CFU/mL and dropping to less than 30 CFU/mL. Macrophage activation by IFN-γ also restricted the growth of the recombinant strains. The numbers of M. smeg::E and M. smeg::A CFUs were lower after 48 h than at the beginning. However, growth of the recombinant strains had partly recovered 72 h after infection (Fig. 3B).
Relative mRNA expression of IL-1β, iNOS, and TNF-α in infected macrophages
Quantitative RT-PCR was used to analyze the relative mRNA expression of IL-1β, iNOS, and TNF-α in ANA-1 macrophages infected with the recombinant strains. Infection with either M. smeg::A or M. smeg::E caused changes in the mRNA expression levels of the indicated cytokines. In macrophages infected with M. smeg::A, mRNA expression of all three cytokines gradually increased, peaking at 48 h, and then decreasing at 72 h. In macrophages infected with M. smeg::E, mRNA expression of IL-1β, iNOS, and TNF-α peaked at 48, 72, and 24 h, respectively. Phagocytosis of M. smeg::E caused greater increases in mRNA expression of these cytokines than did that of M. smeg::A, with the exception of TNF-α mRNA expression at 48 h (Fig. 4).
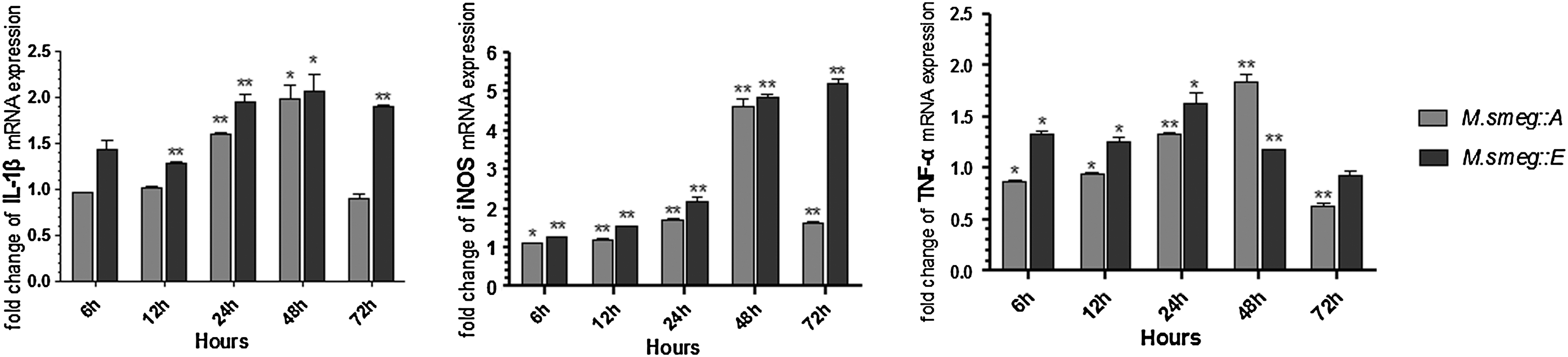
Relative mRNA expression of interleukin (IL)-1β, inducible nitric oxide synthase (iNOS), and tumor necrosis factor (TNF)-α at different time points in ANA-1 challenged by M. smeg::A or M. smeg::E. Asterisk (*) indicates a statistical significance between the M. smeg and the recombinant strains. *p<0.05, **p<0.01.
Role of CD36 in macrophage recognition of M. smeg::A and M. smeg::E
Quantitative RT-PCR was used to analyze the relative mRNA expression of CD36 in ANA-1 macrophages infected with the recombinant strains. In macrophages infected with M. smeg::A, CD36 mRNA expression gradually increased, peaked 48 h after infection, and then fell to the lowest level at 72 h. The CD36 mRNA expression pattern in macrophages infected with M. smeg::E was not as consistent over time as that for M. smeg::A except after 48 h. Overall, M. smeg::E infection stimulated more CD36 mRNA expression than M. smeg::A infection (Fig. 5).
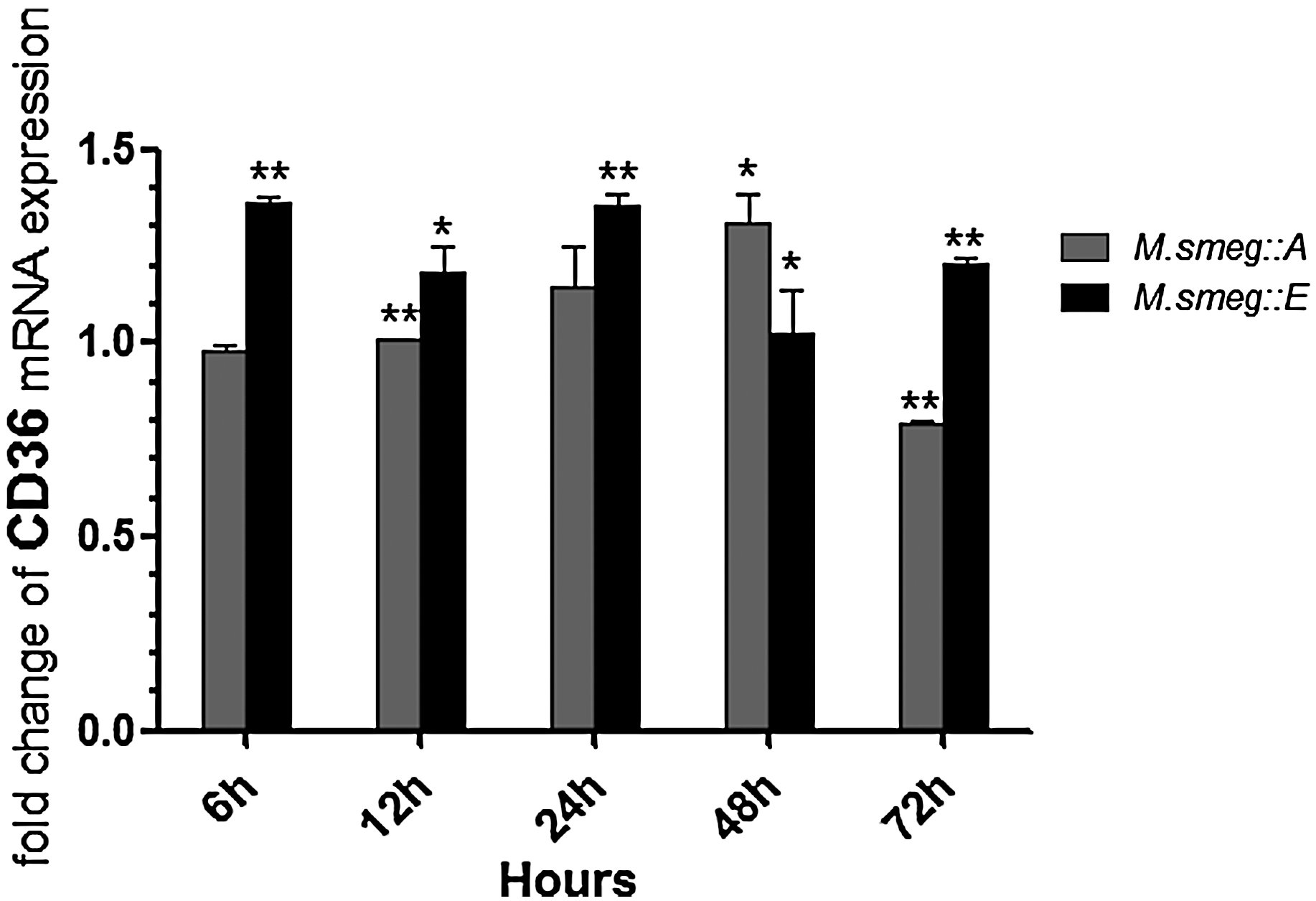
Role of CD36 in recognition of M. smeg::A and M. smeg::E. CD36 mRNA expression in ANA-1 changes at different time points challenged by M. smeg::A or M. smeg::E in real-time quantitative PCR. Asterisk (*) indicates a statistical significance between the M. smeg and the recombinant strains. *p<0.05, **p<0.01.
The numbers of CFUs for each strain in macrophages pretreated with the CD36 antibody were also assessed (Fig. 6). After 12 h, the numbers of CFUs of the recombinant strains were comparable to that of the M. smeg control. Beyond 24 h though, the recombinant strains grew much more rapidly than M. smeg, with significantly more CFUs observed for the recombinant strains than the control. In addition, when the CD36 antibody was used to block the receptor within macrophages before infection with M. smeg::A or M. smeg::E, the relative increases in mRNA expression of the tested cytokines were less than onefold, much lower than in macrophages not treated with the CD36 antibody (Fig. 7).
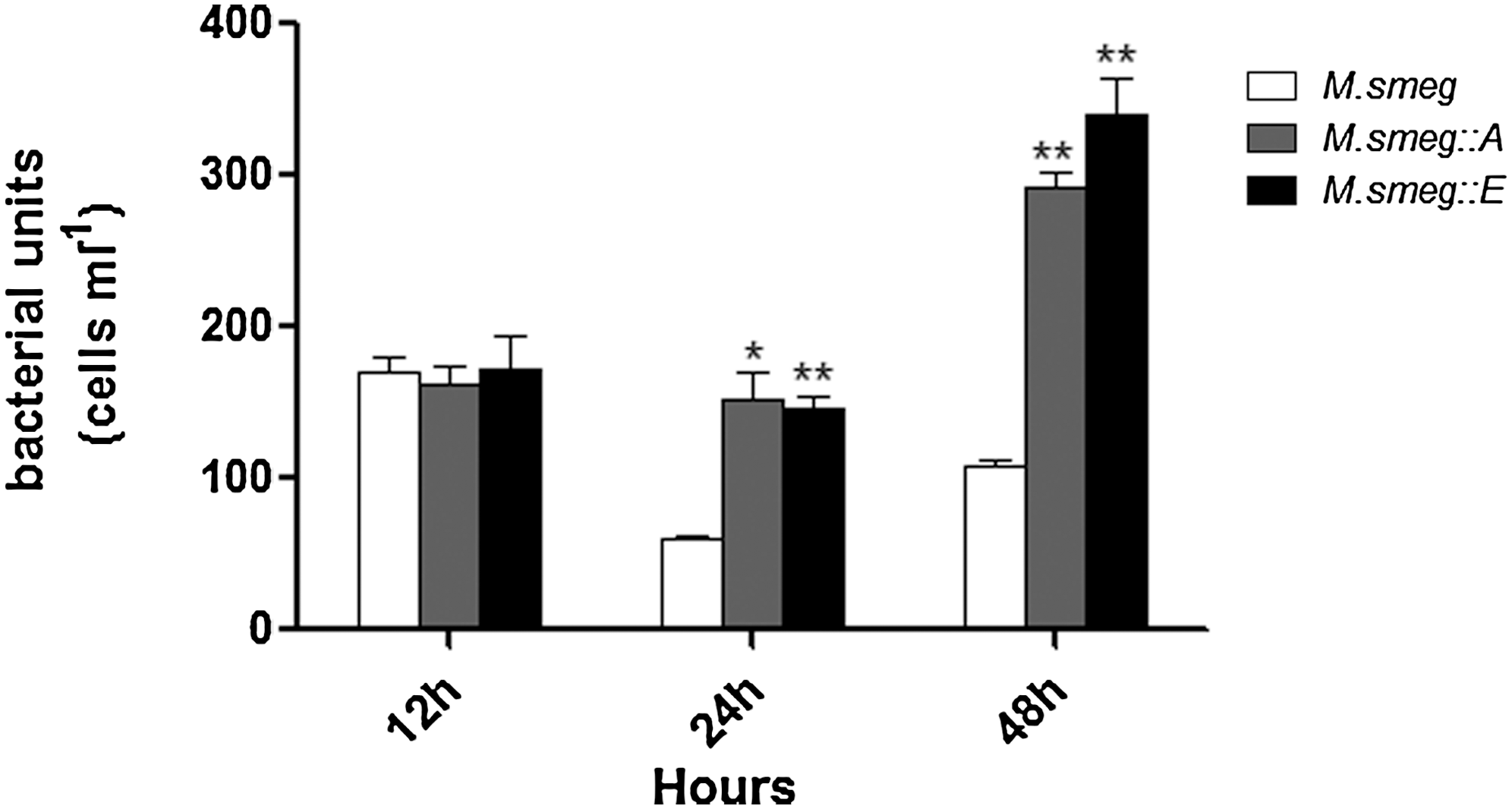
The role of CD36 in recognition of Mce4A and Mce4E proteins and ultimate clearance of indicated strains. Cells were pretreated with the CD36 antibody for 1 h before infection. Asterisk (*) indicates a statistical significance between the M. smeg and the recombinant strains. *p<0.05, **p<0.01.

Relative mRNA expression of IL-1β, iNOS, and TNF-α in ANA-1 after incubation with a CD36-specific antibody before infection with M. smeg::A or M. smeg::E. The samples were collected at 6 h postinfection. Asterisk (*) indicates a statistical significance between the M. smeg and the recombinant strains. *p<0.05, **p<0.01.
Discussion
Mce proteins are important virulence factors in M. tuberculosis (Rengarajan et al., 2005), and mce4 is one of the four mce loci. The complete mce4 system is believed to be responsible for cholesterol import into M. tuberculosis (Pandey and Sassetti, 2008), but the roles of individual genes within this sophisticated cholesterol import system remain poorly understood. They may be linked to the dramatically changing environment to which the bacteria are exposed after phagocytosis. First, we investigated the growth of M. smeg::A and M. smeg::E in the MM containing glycerol or cholesterol. Independent of the addition of glycerol or cholesterol, M. smeg::E performed better than M. smeg::A. Although the MM without glycerol did not support significant bacterial growth, M. smeg::E performed best among the tested strains. Therefore, the Mce4E protein likely enabled the bacteria to survival in the poor nutrient medium, which resulted in M. smeg::E persisting longer intracellularly, especially within IFN-γ-activated macrophages. Our results agree with those of a previous study by Pandey and Sassetti (2008), which demonstrated increased bacterial growth in the MM with added cholesterol. The peak OD600 value for each strain in the MM with cholesterol was higher than that in MM with glycerol, and at later culture times, the cell multiplication rates decreased more slowly. The bacterial growth pattern in the MM without cholesterol was not similar to that in MM without glycerol. These data suggest that the mce4 transport system in M. smegmatis may play a role in the utilization of ethanol as the sole carbon source, a finding also supported by inhibited growth of a mce4 M. tuberculosis mutant in the MM supplemented with ethanol (Pandey and Sassetti, 2008). Second, a higher OD600 for M. smeg in MM with cholesterol was observed compared to the OD600 values for the recombinant strains. Because M. smegmatis contains an ortholog of mce4 (Casali and Riley, 2007), the extra expression of Mce4A or Mce4E could interfere with the cholesterol import by its own mce4 system. However, in the long run, the recombinant M. smeg::E grew arriving at the level of nonrecombinant M. smeg, which demonstrates that Mce4E became of utmost important at later time points. However, all three strains multiplied much more stably in MM with cholesterol than they did in MM without cholesterol; thus, the effect of ethanol on bacterial growth can be disregarded.
When macrophages are activated by T-cell-derived cytokines, specifically interferon IFN-γ, a variety of antibacterial effectors, such as phagosomal maturation and nitric oxide production, are initiated. The pathogen must adapt to these harsh and continually changing conditions, and cholesterol plays an important role in that adaptation. Although the precise mechanism remains unclear, evidence suggests that the mce4 system is involved. Mutations in mce4 confer limited growth ability in IFN-γ-activated macrophages (Pandey and Sassetti, 2008). Additionally, upregulated Mce4E protein expression persists for 72 h, even though expression levels of other proteins have already returned to low levels (Rengarajan et al., 2005). Our data on the bacteria CFU showed the same tendency. Whether in intact macrophage or IFN-γ-activated macrophages, the recombinant M. smeg::E grew the best of the strains tested. The Mce4A protein is present in the cell wall fraction of stationary phase M. tuberculosis H37Rv cultures. The recombinant Mce4A protein promotes the invasion of a nonpathogenic strain of Escherichia coli into nonphagocytic Hela cells (Saini et al., 2008). The intracellular number of recombinant M. smeg::A became highest but decreased after 12-h infection in our experiment. The bacteria proliferated greatly after 48-h infection in IFN-γ-activated macrophages, which indicated that the Mce4A facilitated the invasion of bacteria and adaptation to the harsh environment of macrophages. Among products of the mce operons, Mce1A was first shown to confer on recombinant E. coli the capacity to invade and survive within macrophages (Casali and Riley, 2007). Later, mce3A-F was also reported to be involved in entry and survival inside macrophages (Ahmad et al., 2004, 2005).
Using expression of Mce4A and Mce4E, our previous studies showed that both proteins may induce the body's inflammation response to M. bovis and, therefore, may play an important role in the immune response (Xu et al., 2007, 2008). In this study, infection with each of the recombinant strains significantly upregulated the mRNA expression of three cytokines (IL-1β, iNOS, and TNF-α) in ANA-1 macrophages. TNF-α and IL-1β are involved in the Th-1 response, and upregulation of these cytokines indicates that both the Mce4A and Mce4E proteins can activate the Th1 response. iNOS mRNA expression in macrophages infected with either recombinant strain was much higher compared with the other cytokines. iNOS generates NO, and high concentrations of NO can result in cell damage and death. Specifically, NO production inhibits respiration and induces dormancy of M. tuberculosis (Voskuil et al., 2003). Dormant ovoid M. tuberculosis exhibits a low metabolic activity (Shleeva et al., 2011). A readily available carbon source for energy generation (Pandey and Sassetti, 2008) and a lower metabolic activity ensure that the recombinant strains adapt more effectively to changes in the intracellular environment and lead to better survival of the strains in activated macrophages. Compared to the mRNA expression levels of IL-1β and iNOS that of TNF-α seemed low. TNF-α is a primary proinflammatory and proimmune cytokine that plays an important role in the host immune response against M. bovis (Roach et al., 1999; Sedgwick et al., 2000; McDermott, 2001). TNF-α is involved in the formation of protective tuberculosis granulomas and regulates the expression of additional cytokines that contribute to a protective immune response (Kaneko et al., 1999; Mohan et al., 2001; Ehlers, 2003). From this point of view, expression of Mce4A or Mce4E in M. smegmatis was not expected to exert a strong effect on the virulence of the parental M. smegmatis but to induce a weak host immune response and protection.
CD36 belongs to the class B scavenger family and presents as a pattern recognition receptor mediating innate immune responses to several pathogens, including mycobacteria (Febbraio et al., 2001). Peste, a CD36 family member, is one specific factor that is required for the uptake of mycobacteria, but not for E. coli or Staphylococcus aureus (Philips et al., 2005). The mycobacterial burden in CD36−/− mice infected by Bacillus Calmette-Guérin is less than that observed in CD36+/+ mice. In this study, we used qRT-PCR to test CD36 mRNA expression in ANA-1 macrophages treated with M. smeg::A or M. smeg::E. We also assessed CFU counts for each strain in the cells preincubated with the CD36 antibody. However, the observed change in CD36 mRNA expression was not consistent with changes in intracellular CFU counts. On one hand, serum components directly compete with bacteria for the receptor, leading to an inhibition effect on bacterial binding to the cell surface (Schafer et al., 2009). On the other hand, there are a number of other receptors on the cell surface that are more sensitive and specific to the recognition of bacterial products or envelope components expressed during the infection process (Flannagan et al., 2012). CD36 appeared to have little impact on the clearance of the recombinant bacteria. Therefore, we focused on changes in the mRNA expression of IL-1β, iNOS, and TNF-α during the early infection period when a CD36-specific antibody was used to block the receptor. Interestingly, IL-1β, iNOS, and TNF-α mRNA expression levels in macrophages infected with M. smeg::E at 6 h postinfection were lower than in macrophages infected with M. smeg::E, but not pretreated with the CD36 antibody. However, in macrophages infected with M. smeg::A, no differences in cytokine mRNA expression profiles were observed between CD36 antibody-treated macrophages and those not treated with the CD36 antibody. Given that the Mce4A and Mce4E proteins belong to the mce4 cholesterol transport system and considering the function of CD36 in recognizing the bacterial products, we hypothesize that the Mce4E protein is more effective at facilitating bacterial persistence in macrophages than Mce4A, thus leading to better survival after phagocytosis by macrophages.
A previous microarray study reported the expression profiles of the mce4A and mce4E genes in M. tuberculosis (Van der Geize et al., 2007). Our study demonstrated that the differences in the expression profiles of Mce4A and Mce4E may be related not only to gene organization but also to their function in adaptation to the harsh environment within the cells. CD36 could facilitate the phagocytosis of wild-type and recombinant M. smegmatis. Blocking CD36 could decrease the bacterial numbers phagocyted by macrophages. Survival of the parent M. smegmatis decreased, but for the both recombinant M. smegmatis strains, the proliferation of the bacteria increased greatly, showing that Mce4A and Mce4E assist in bacterial adaptation to the acidic and bactericidal environment after the cells were activated. Moreover, after blocking CD36, mRNA expression of IL-1β, iNOS, and TNF-α reduced at 6 h postinfection in macrophages infected with M. smeg::E, whereas no obvious changes in the expression of these cytokines were observed in cells infected with M. smeg::A., suggesting that Mce4E is more effective for initiation of the host immunological response than is Mce4A.
Footnotes
Acknowledgments
This work was supported by the MoSTRCUK international cooperation project (Project No. 2013DFG32500), National High Technology Research and Development Program of China (863 Program, Project No. 2012AA101302), National Natural Science Foundation of China (Project No. 31172293, No. 31272532), Funding of State Key Lab of Agrobiotechnology (Project No. 2012SKLAB06-14), and 2014 CAU Foreign Experts Major Projects (Project No: 2012z018).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.