
Abstract
Natriuretic peptide precursor-A (NPPA) is an early and specific marker for functional myocardium of the embryonic heart. NPPA gene encodes for a precursor of atrial natriuretic peptide (ANP). So far, three alternatively spliced variants have been reported for NPPA in human. In mouse, no alternatively spliced transcript of NPPA has been reported. In the current study, we investigated the expression of NPPA gene during cardiac differentiation of mouse adipose-tissue-derived stem cells (ADSCs) and embryonic stem (ES) cells. As revealed by reverse-transcription polymerase chain reaction analysis, 2-week-differentiated cells expressed some cardiac-specific makers, including ANP. Three additional intron-retained splice variants of NPPA were also detected during cardiac differentiation of the ADSCs and ES cells. In addition, we detected three intron-retained splice variants of NPPA in 8.5-day mouse embryonic heart. In the mature cardiomyocytes of 1-week-old mice, only the correctly spliced isoform of NPPA gene was expressed. Freshly isolated stromal vascular fraction also expressed one intron-retained isoform of NPPA gene. In conclusion, our findings have provided evidence for the expression of intron-retained splices of NPPA mRNA during the early stages of mouse cardiogenesis as well as in the mouse adipose tissue.
Introduction
N
The expression pattern of NPPA during development of cardiac chamber is highly dynamic (Zeller et al., 1987; Houweling et al., 2005). So far, three alternatively spliced variants have been reported for NPPA mRNA in human, including a correctly spliced transcript and variants with either retained intron 1 or retained introns 1 and 2. The correctly spliced variant has its highest expression level in heart, while it is moderately expressed in some other tissues, including prostate, pancreas, and small intestine (Annilo et al., 2009). In mouse, no alternatively spliced isoform of NPPA gene has been reported previously (Annilo et al., 2009).
Stem cells have been used as valuable models to investigate patterns of gene expression during cellular differentiation. In the current study, we studied the expression profile of NPPA gene during cardiac differentiation of mesenchymal and embryonic stem (ES) cells. For this purpose, ES cells were differentiated at the presence or absence of bone morphogenetic protein-4 (BMP4). Cardiac differentiation of the adipose-tissue-derived stem cells (ADSCs) was induced by 5-azacytidine or BMP4, which are known cardiogenic factors (Schultheiss et al., 1997; Rangappa et al., 2003b; Zhang et al., 2005; Taha and Valojerdi, 2008), as well as by direct or indirect coculture with mouse cardiomyocytes.
As previously discussed (Rangappa et al., 2003a), chemical and physical components of the microenvironment play a critical role in determining the ultimate fate of developing cells. In direct coculture of ADSCs with viable cardiomyocytes, both chemical and physical factors can influence the differentiation of ADSCs (Rangappa et al., 2003a). However, when the ADSCs are cocultured with paraformaldehyde-fixed cardiomyocytes, just the physical contact between the ADSCs and cardiomyocytes can affect and direct the differentiation (Badorff et al., 2003). In an indirect coculture method using culture inserts, there is no physical contact between different cell types, and they can influence each other only by soluble factors (Liu et al., 2012). Using these models, we identified three alternatively spliced transcripts of NPPA for the first time. We further isolated the cardiac area of 8.5-day mouse embryos and showed the expression of intron-retained isoforms of NPPA mRNA at this early stage of embryonic heart development.
Materials and Methods
Isolation and culture of mouse ADSCs
All animal works conducted in this study were according to the regulations of animal care and use committee of the National Institute of Genetic Engineering and Biotechnology. Inguinal fat pads of 10–12-week-old NMRI mice were isolated and digested using 2 mg/mL collagenase A (Roche Applied Science), as described previously (Taha and Hedayati, 2010). Isolated cells were plated at 5×104 cells/mL in 6-cm tissue culture plates and cultured in a growth medium containing Dulbecco's modified Eagle's medium (DMEM) and 20% fetal bovine serum (FBS) (both from Gibco®, Life Technologies). The cells were daily observed and passaged after reaching 80–90% confluency. The culture media were changed every other day.
Cardiac differentiation of the ADSCs
Third-passaged ADSCs were cultured and propagated to a confluent state. Cardiac differentiation of the ADSCs was induced either by 5-azacytidine or BMP4 treatment or by coculture with mouse cardiomyocytes.
5-Azacytidine treatment
5-Azacytidine (Sigma) with a final concentration of 10 μM was added to the culture medium for 24 h. ADSCs were cultured in DMEM plus 10% FBS.
BMP4 treatment
The confluent ADSCs were cultured in a medium containing 15% Knockout™ Serum Replacement and 10 ng/mL bone morphogenetic protein 4 (BMP4; Sigma) during the first 5 days of differentiation. After this period, BMP4 was removed, and differentiation of the ADSCs was continued in DMEM containing 10% FBS without any additives.
Coculture of the ADSCs with mouse cardiomyocytes
Ventricular cardiomyocytes were isolated from the hearts of 4–7-day-old mice. The hearts were surgically removed, collected in tissue culture plates, and rinsed in cold PBS to remove blood and cell debris. Ventricles were dissected, minced, and dissociated by 0.2% collagenase II in PBS containing 1% BSA (Sigma). Cell suspension was transferred to gelatinized tissue culture plates and cultured for 1–1.5 h to remove cardiac fibroblasts. The same procedure was repeated.
Monolayers of ventricular cardiomyocytes were prepared by culturing the cells in gelatinized tissue culture plates and cocultured with third-passaged ADSCs directly by cell–cell contact or indirectly using culture inserts. For the direct coculture, monolayers of the cardiomyocytes were fixed by incubation with 2% paraformaldehyde, and after three washes with PBS, third-passaged ADSCs were cultured on the top of these cells (Badorff et al., 2003). As mentioned by Badorff et al. (2003), paraformaldehyde-fixed cells cannot fuse with other cells but provide an intact cell surface for coculture. After coculture, differentiated ADSCs were isolated by a brief trypsinization. Then, the cells were cultured in gelatinized tissue culture plates for 24 h. Immediately after attachment of the ADSCs, the cultures were washed to remove all nonviable unattached cells. In this way, only the differentiated cells are retained and paraformaldehyde-fixed cardiomyocytes are eliminated.
For the indirect coculture, about 8×104 cardiomyocytes were seeded on 0.4-μm porous Transwell-Clear inserts (Greiner Bio-One GmbH). Culture inserts were placed in six-well tissue culture plates containing third-passaged ADSCs (Liu et al., 2012). Medium of both the coculture systems consisted of DMEM and 10% FBS (both from Gibco®, Life Technologies). Third-passaged ADSCs were cocultured with the same cells served as the control group.
Cardiac differentiation of ES cells
The mouse ES cell line Royan B1 (Royan Stem Cell Bank, Royan Institute, RSCB0001) was cultured on top of a feeder layer of mitomycin C-treated mouse embryonic fibroblast (MEF) at the presence of leukemia inhibitory factor (LIF; Chemicon, ESGRO), as previously described (Taha et al., 2007). The ES cells were dissociated from the feeder layer, and differentiation was initiated with embryoid body (EB) formation using hanging-drop method (Taha et al., 2007). In summary, 20 μL of hanging drops containing about 800 cells was cultured for 2 days and then EBs were maintained in suspension culture for 5 days. Seven-day-old EBs were plated in 0.1% gelatin-coated tissue culture plates and cultured for 3 weeks. Differentiation medium consisted of Knockout™ Dulbecco's modified Eagle's medium (Ko-DMEM, high-glucose, with sodium pyruvate; Gibco®, Life Technologies), 15% FBS (FBS, ES cell qualified; Gibco®, Life Technologies), 0.1 mM β-mercaptoethanol (Sigma), 1 mM L-glutamine, 1% nonessential amino acid stock, and 1% penicillin-streptomycin (all from Gibco®, Life Technologies). Some EBs were also treated with 10 ng/mL BMP4 during the suspension stage.
Reverse-transcription polymerase chain reaction
Total RNA was extracted from the cells using High Pure RNA Isolation Kit (Roche Applied Science) and treated with DNase I (Roche Applied Science) according to the manufacturer's instructions. For cDNA synthesis, 1 μg of total RNA was reverse transcribed to cDNA using oligo-dT primers and RevertAid™ H Minus First Strand cDNA Synthesis Kit (Fermentas, Thermo Fisher Scientific, Inc.). cDNA samples were used in PCR using primers described in Supplementary Table S1 (Supplementary Data are available online at
Sequencing analysis
PCR products were extracted from the agarose gel using Accuprep Gel Purification Kit (Bioneer) and sequenced (Macrogen, Inc.). DNA sequencing results were visually analyzed by Sequence Scanner v1.0 (Applied Biosystems) aligned to the mouse genome using BLAST® alignment tool (NCBI, NIH). The novel sequences detected in this study were submitted to GenBank database and were acquired accession numbers (KC526925, KC526926, and KC526927).
Immunocytochemistry
Differentiated cells were fixed by incubation with 4% paraformaldehyde, permeabilized by 0.5% Triton X-100 (Sigma), and blocked with 10% goat serum (Gibco®, Life Technologies). The cells were incubated with primary and secondary antibodies for 45 min each at 37°C. Antibodies used in this study included monoclonal antibodies for cardiac muscle troponin-I (CTnI; Chemicon) and anti-mouse FITC-conjugated IgG (Sigma), both with dilution of 1:100. Preparations were examined and photographed by an inverted fluorescence microscope connected to a high-resolution digital camera (Eclipse TE 2000U; Nikon).
Results
Isolation and culture of mouse ADSCs
After three passages, the ADSCs developed a homogenous fibroblast-like morphology (Supplementary Fig. S1A). We previously studied the molecular phenotype of the third-passaged ADSCs and showed that more than 98% of the cells express CD29 (beta-1 integrin), CD44 (hyaluronate receptor), and CD105 (endoglin) (Taha and Hedayati, 2010; Jafarzadeh et al., 2014), which are known mesenchymal stem cell markers (Dominici et al., 2006; Mitchell et al., 2006; Schaffler and Buchler, 2007).
Cardiac differentiation of the ADSCs
Cardiac differentiation of the ADSCs was induced by 5-azacytidine, BMP4, or by direct or indirect coculture with mouse cardiomyocytes. During the first week of differentiation, some round-shaped cells appeared in all the experimental groups (Supplementary Fig. S1B). During the next weeks, these cells grew in size and proliferated. Moreover, some cells developed a tube-like morphology (Supplementary Fig. S1C–G).
The expression of cardiac transcription factors and cardiac-specific genes was detected in the heart tissue of 1-week-old mice as the positive control. GATA binding protein-4 gene (GATA4) and myocyte enhancer factor 2C (MEF2C) were weakly expressed, due to maturation of the cardiomyocytes (Fig. 1).
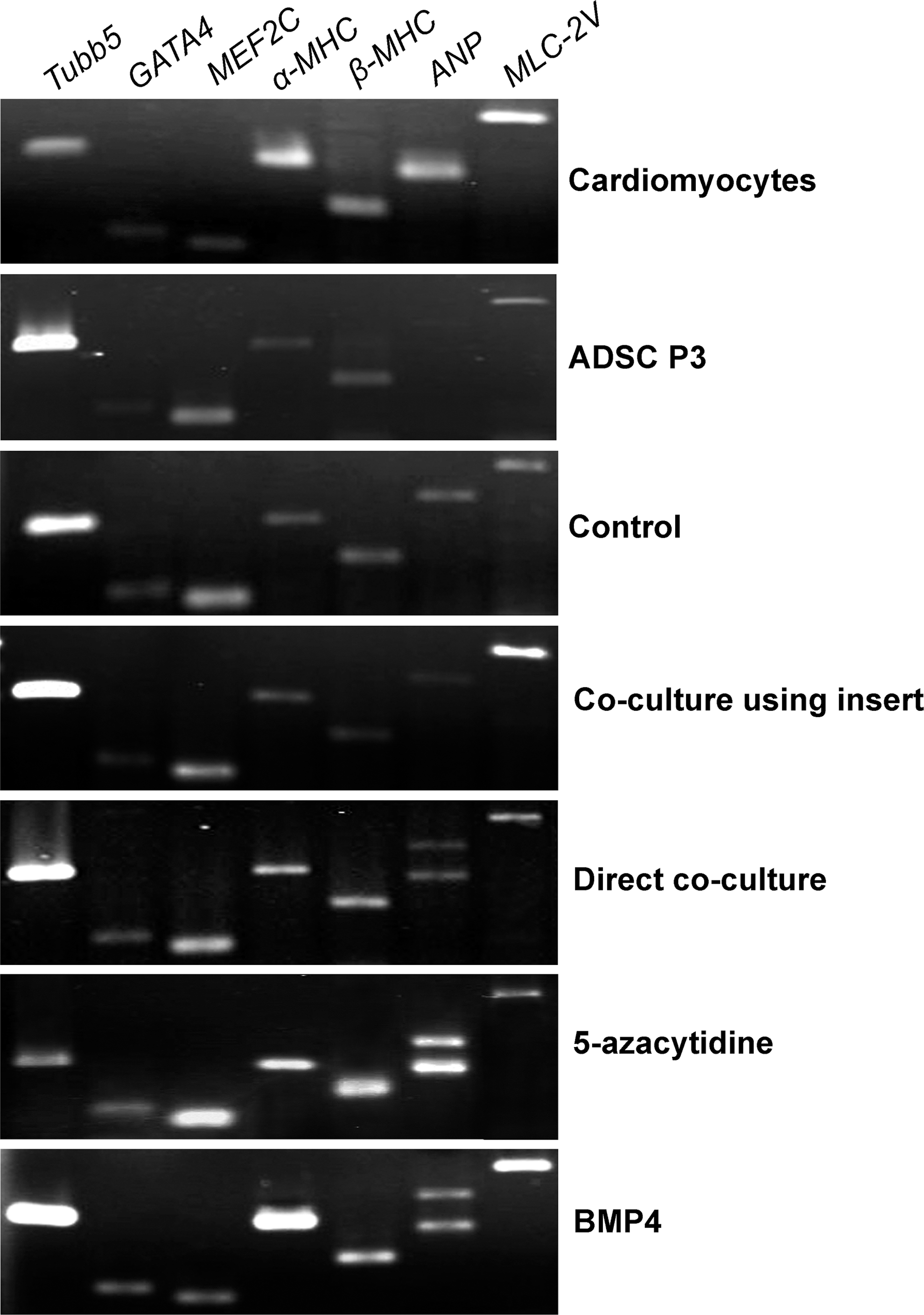
The expression of some cardiac transcription factors and cardiac-specific genes in 2-week-differentiated, adipose-tissue-derived stem cells (ADSCs). The cardiomyocytes isolated from the hearts of 1-week-old mice were used as the positive control (first upper row). Third-passaged ADSCs (ADSC P3) were used as the initiating cells before induction (second upper row). The cells differentiated in 10% fetal bovine serum (FBS)–containing medium without any additional factors were used as the control group of differentiation (third upper row). Cardiac differentiation of the ADSCs was induced by direct or indirect coculture with mouse cardiomyocytes, as well as by 5-azacytidine and BMP4 treatment (the last four rows).
Third-passaged ADSCs expressed cardiac transcription factors and cardiac-specific genes, including α- and β-myosin heavy chain genes (α- and β-MHC), ANP, and myosin light chain-2V gene (MLC-2V) (Fig. 1). This may reflect potential of the ADSCs for cardiac differentiation. Two weeks after cardiac induction, the expression of GATA4, MEF2C, α-MHC, β-MHC, and MLC-2V was detected in all the experimental groups (Fig. 1).
As revealed by immunocytochemistry, ADSC-derived cardiomyocyte-like cells of the control, 5-azacytidine, BMP4, and coculture groups showed positive immunostaining for cardiac troponin-I, which is a cardiac-specific protein (Fig. 2A–E). Cardiomyocytes isolated from the hearts of 1-week-old mice were used as the positive control (Fig. 2F). Immunostaining of the third-passaged ADSCs for cardiac troponin-I protein was negative.

Immunostaining of the ADSC-derived cardiomyocytes for cardiac troponin-I protein.
Functionality of the ADSC-derived cardiomyocytes
None of the experimental conditions we used were sufficient for differentiation of the ADSCs into functional beating cardiomyocytes.
The expression of alternatively spliced transcripts of NPPA during cardiac differentiation of ADSCs
PCR analysis using the ANP primers (F1 and R1 in Fig. 3A) in the control and indirect coculture groups amplified a product longer than the correctly spliced ANP mRNA (385 bp instead of 282 bp) (Fig. 1). In the direct coculture, 5-azacytidine, and BMP4 groups, two fragments with the different sizes of 282 and 385 bp were detected (Fig. 1). Sequencing of the 385-bp fragment and BLAST analysis revealed a NPPA splice variant with retained intron 1 (Fig. 3B).
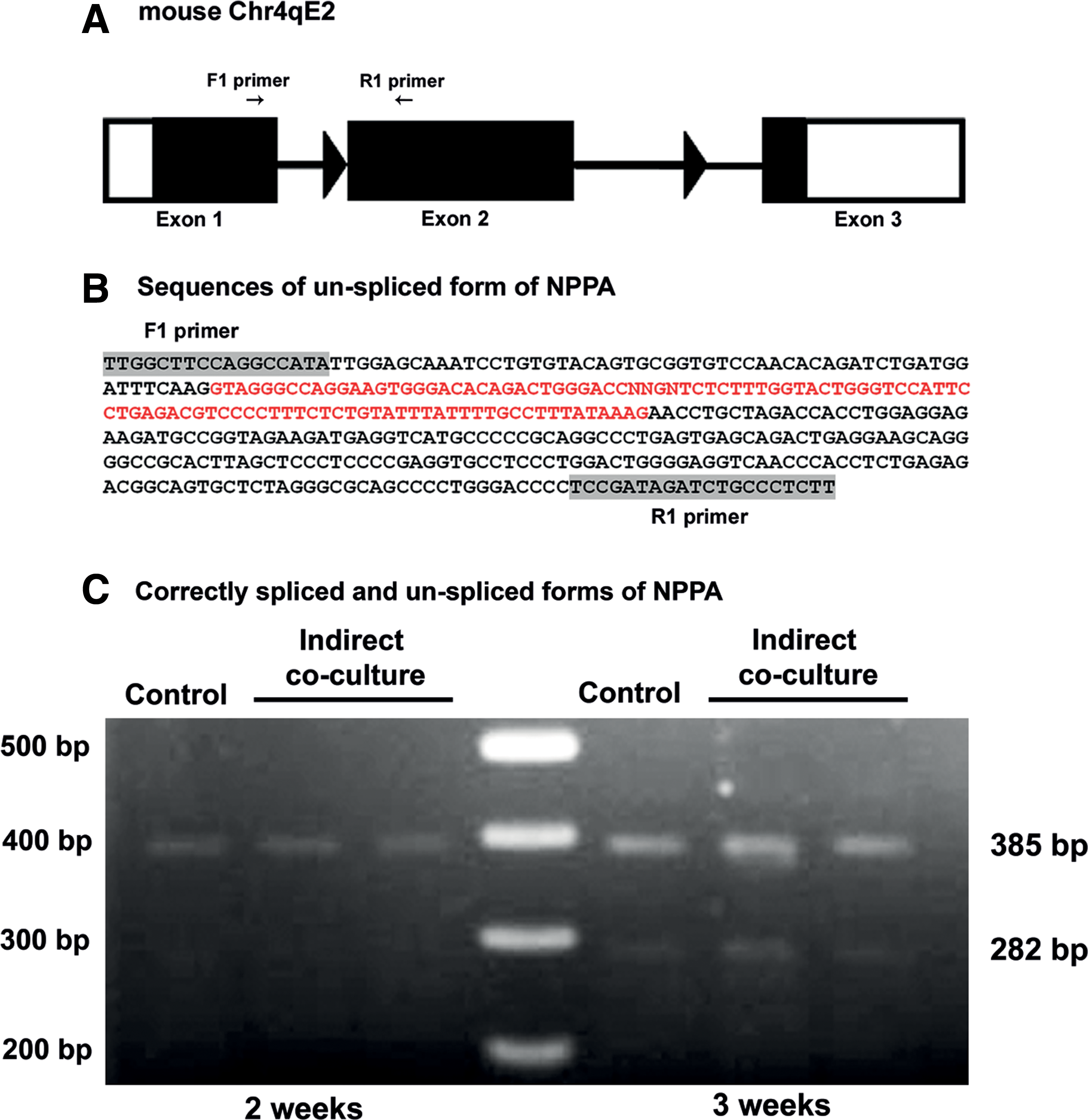
Based on these findings, we asked whether NPPA splicing depends on the stage of cardiac development. To answer this question, we extended differentiation of the ADSCs in the control and indirect coculture systems for one additional week. At this time, RT-PCR analysis revealed the expression of both correctly and alternatively spliced isoforms of NPPA (Fig. 3C).
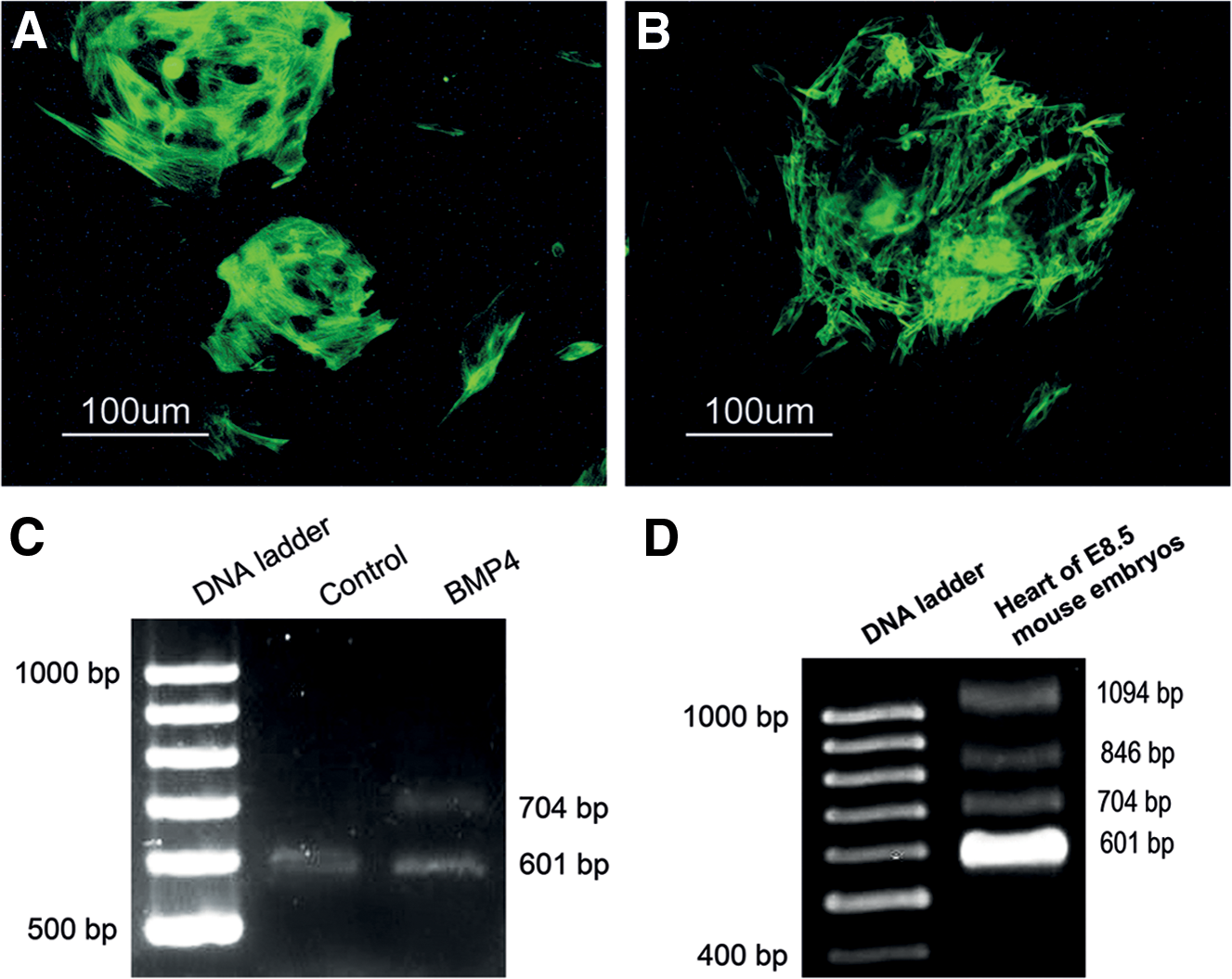
We also performed additional PCR experiments with the second set of primers, F2 and R2 (Fig. 4A), which detects NPPA transcripts with retained introns 1 and 2 (Annilo et al., 2009). PCR using this primer set resulted in the amplification of four products with different sizes. As shown in Figure 4B, the smallest band with the size of 601 bp represents the expression of correctly spliced variant, and the other bands show alternatively spliced variants of NPPA gene. Interestingly after 3 weeks of cardiac induction, differentiated cells expressed different splice variants of the NPPA gene. When the culture period was extended for another week, the variants of NPPA gene were disappeared and only a weakly expressed intron 1-retained variant was observable (Fig. 4C). Beating cardiomyocytes isolated from the hearts of 1-week-old mice only expressed the correctly spliced isoform of NPPA (Fig. 4C).

To develop more convincing evidence concerning the expression of NPPA splice variants during cardiac differentiation of mouse ADSCs, we assessed the expression of these splices after cultivation of the ADSCs in a noncardiogenic condition. The NPPA splice variants were not expressed in the ADSCs cultured in neurogenic differentiation media (Fig. 4C).
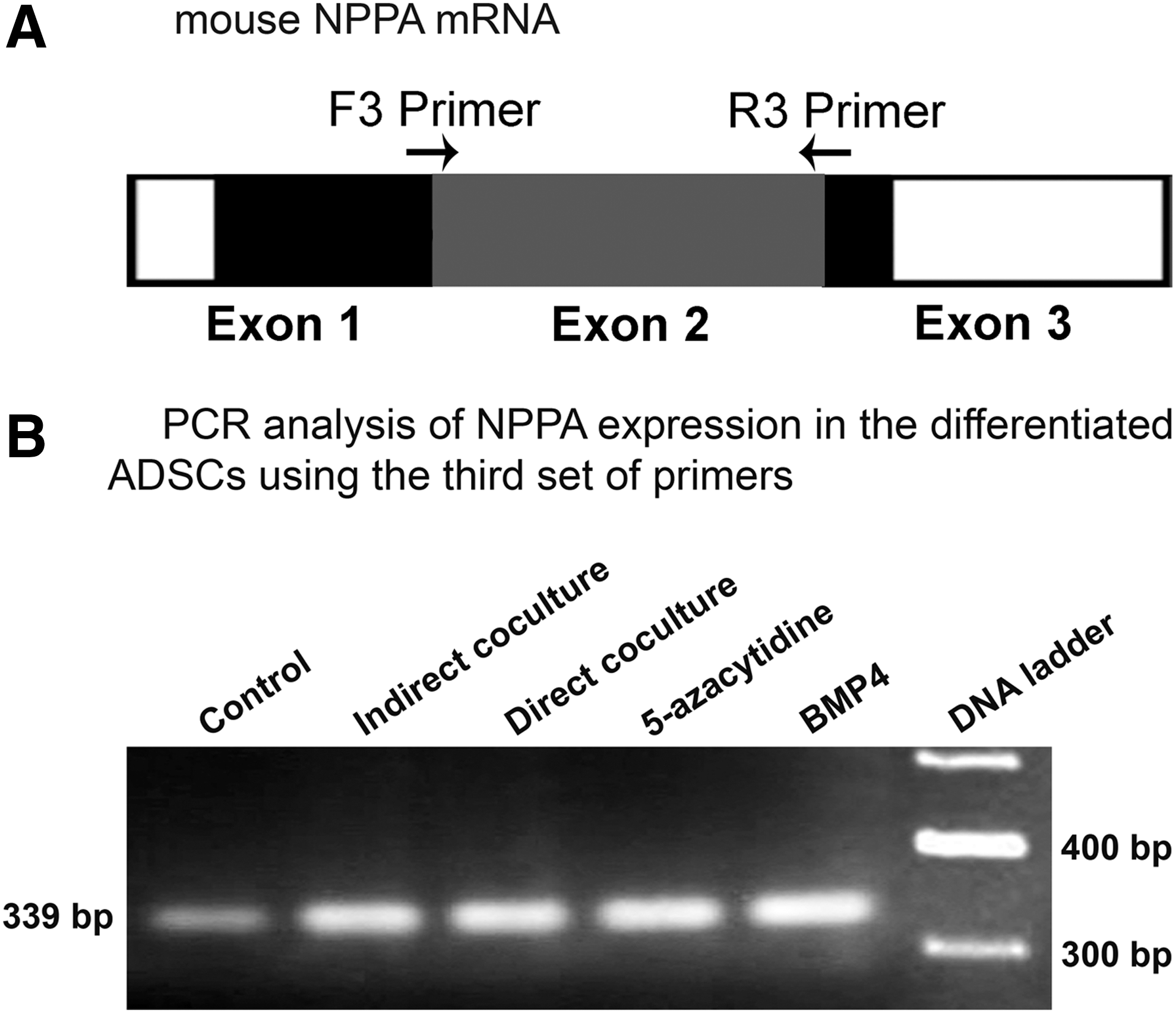
To ensure that amplification of the intron-retained splices of NPPA gene was not the consequence of contamination with genomic DNA, a third set of primers (F3 and R3) was designed to span exon–exon boundaries (Fig. 5A). These primers specifically attach to the nonintronic segments of cDNA. PCR using this set of primers resulted in the amplification of a 339-bp product (Fig. 5B). Additional PCR experiments were also performed with combinations of the fourth (Supplementary Table S1) and third sets of primers. As shown in Figure 6A, PCR using F3 and R4 primers led to amplification of three products. The first transcript with the size of 355 bp (

The expression of NPPA variants in the freshly isolated stromal vascular fraction
In the current study, freshly isolated stromal vascular fraction (SVF) expressed an isoform of NPPA with retained introns 1 and 2 (Fig. 4D).
The expression of NPPA splice variants during cardiac differentiation of ES cells
The expression of NPPA splice variants was studied in the ES-cell-derived EBs of the control and BMP4 treatment groups. Two weeks after plating, both groups showed positive immunostaining for cardiac troponin-I (Fig. 7A, B). Nevertheless, spontaneously contracting cardiomyocytes of the control group formed thicker and more branching bundles. When the EBs were assessed for the expression of NPPA splices, EBs of the control group showed the expression of correctly spliced variant of NPPA mRNA, while the BMP4-treated EBs expressed correctly spliced and intron-1-retained isoforms of NPPA mRNA (Fig. 7C).
Cardiac differentiation of mouse embryonic stem (ES) cells.
The expression of NPPA variants in the mouse embryonic heart
To assess the expression of NPPA splice variants during heart development, the cardiac area of 8.5-day mouse embryos was isolated surgically. Total RNA was isolated and RT-PCR analysis was performed with F2 and R2 primers. This revealed the expression of the same four NPPA mRNA splices as observed in 3-week-differentiated ADSCs (Fig. 7D).
Discussion
Cardiac differentiation of the ADSCs
Cardiac differentiation of the ADSCs was induced by 5-azacytidine or BMP4, which are known cardiogenic factors (Schultheiss et al., 1997; Rangappa et al., 2003b; Zhang et al., 2005; Taha and Valojerdi, 2008), or by direct or indirect coculture with mouse cardiomyocytes. The role of 5-azacytidine in cardiac differentiation of MSCs has been controversial. Several investigators have reported that 5-azacytidine alone is not sufficient for differentiation of mesenchymal stem cells (Martin-Rendon et al., 2008; Lee et al., 2009; Wan Safwani et al., 2012). In our study, both 5-azacytidine and BMP4 treatment induced the expression of cardiac-specific genes and proteins, but they were not sufficient for differentiation of the ADSCs into functional contracting cells. Some additional growth and differentiation factors may be required to induce functional maturation of the ADSC-derived cardiomyocytes. Moreover, direct and indirect coculture systems induced the expression of some cardiac markers, although neither the chemical niche created by viable cardiomyocytes nor the physical contact with nonviable cells was sufficient for generation of mature cardiac cells. As reported previously (Choi et al., 2010; Metzele et al., 2011), direct contact with the viable cardiomyocytes may be necessary for generation of functional beating cells.
The expression of alternatively spliced variants of NPPA gene during cardiac differentiation of the ADSCs
Our study has demonstrated the expression of four alternatively spliced transcripts of NPPA after differentiation of ADSCs into immature cardiomyocyte-like cells. Variants with retained intron 1 and retained introns 1 and 2 are similar to those previously reported for NPPA in human (Annilo et al., 2009), while the 846-bp transcript that includes intron 1 and a 142-bp fragment of intron 2 is reported for the first time. Since in our culture conditions, the ADSCs could not generate functional cardiomyocytes, it may be suggested that the NPPA splice variants are expressed in the early stages of cardiac differentiation.
The expression of alternatively spliced variants of NPPA gene during cardiac differentiation of the ES cells
We previously studied the role of BMP4 in cardiac differentiation of mouse ES cells and showed that BMP4 treatment decreases population of beating EBs, downregulates the expression of cardiac-specific genes, and retards the ultrastructural and functional maturation of the ES-cell-derived cardiomyocytes in serum-containing media (Taha et al., 2007). In the current study, we studied the expression of alternatively spliced transcripts of NPPA in the EBs of the control and BMP4 treatment groups. As revealed by RT-PCR analysis, contracting EBs of the control group showed the expression of correctly spliced variant of NPPA mRNA, while the BMP4-treated EBs expressed both the correctly spliced and intron-1-retained transcripts of NPPA. These findings provided more evidence regarding the stage-dependent expression of NPPA mRNA splices during cardiac development.
The expression of NPPA variants in the freshly isolated SVF
We showed that freshly isolated SVF cells express an isoform of NPPA with retained introns 1 and 2. So far, only the expression of one correctly spliced variant of NPPA gene has been reported in mouse tissues. Our findings confirm the expression of one intron-retained splice of NPPA gene in the adult mouse adipose tissue for the first time.
The expression of NPPA variants in the mouse embryonic heart
The cardiac area of 8.5-day mouse embryos was isolated and examined for the expression of alternatively spliced NPPA transcripts. PCR analysis revealed the expression of the same four NPPA mRNA splices as observed in 3-week-differentiated ADSCs. Based on previous studies, full maturation of mammalian cardiomyocytes only develops postnatally (Viragh and Challice, 1969). This might be a reason for the expression of four NPPA mRNA variants in the embryonic heart and the expression of the correctly spliced variant in the cardiomyocytes of 1-week-old mice.
Based on previous reports, NPPA expression is tightly regulated during embryonic development of heart (Zeller et al., 1987; Houweling et al., 2005). Annilo et al. (2009) found a strong expression of correctly spliced NPPA mRNA in human heart and a moderate expression in a number of human tissues, such as prostate, pancreas, and small intestine. In addition, they observed the expression of NPPA isoforms with retained intron 1 or retained introns 1 and 2 in several human tissues. However, according to these investigators, the expression profile of NPPA in mouse and human was different. In mouse, NPPA was strongly expressed in some tissues, such as brain, lung, and liver, where human NPPA was expressed weakly. They did not detect any alternatively spliced or intron-retained transcript of NPPA in mouse tissues, and this is in contrast to our result concerning the expression of intron-retained transcripts of NPPA during embryonic development and in the adult mouse adipose tissue.
Conclusions
In the present study, cardiac differentiation of the ADSCs was induced by 5-azacytidine, BMP4, or coculture with the mouse cardiomyocytes. Differentiated ADSCs expressed some cardiac markers but did not generate functional beating cardiomyocytes. After induction by different cardiogenic factors, ADSCs expressed four alternatively spliced variants of NPPA gene. When cardiac differentiation was continued for a longer duration, the expression of different splices of NPPA gene was diminished, while the expression of correctly spliced NPPA was enhanced. The 8.5-day mouse embryonic heart expressed both the correctly spliced and three alternatively spliced variants of NPPA, while mature cardiomyocytes of 1-week-old mice and ES-cell-derived beating cardiomyocytes expressed the correctly spliced transcript of NPPA. The expression of alternatively spliced variants of NPPA during cardiac development in mouse has not been reported previously. We also showed for the first time that freshly isolated SVF cells express a splice variant of NPPA gene with retained introns 1 and 2. These findings indicate that alternatively spliced variants of NPPA gene are expressed during the early stages of mouse cardiogenesis as well as in the mouse adipose tissue. However, further studies would be required to clarify the expression of these variants in the other mouse tissues and the functional significance of these transcripts.
Footnotes
Acknowledgments
This study was supported by research grants from the National Elites Foundation and the National Institute of Genetic Engineering and Biotechnology (343).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.