
Abstract
Cytolethal distending toxin B (cdtB) is a conserved virulence factor in Salmonella enterica serovar Typhi. Here we report the presence and functionality of cdtB in some nontyphoidal Salmonella (NTS) serovars, including Salmonella Javiana (cdtB+wt S. Javiana), isolated from imported food. To understand the role of cdtB in NTS serovars, a deletion mutant (cdtB− ΔS. Javiana) was constructed. Macrophages were infected with cdtB+wt S. Javiana (wild type), cdtB− Δ S. Javiana (mutant), and cdtB-negative NTS serovar (S. Typhimurium). Cytotoxic activity and transcription level of genes involved in cell death (apoptosis, autophagy, and necrosis) were assessed in infected macrophages. The cdtB+wt S. Javiana caused cellular distension as well as high degree of vacuolization and presence of the autophagosome marker LC3 in infected macrophages as compared with cdtB− ΔS. Javiana. The mRNA expression of genes involved in the induction of autophagy in response to toxin (Esr1 and Pik3C3) and coregulators of autophagy and apoptosis (Bax and Cyld) were significantly upregulated in cdtB+ wt S. Javiana-infected macrophages. As autophagy destroys internalized pathogens in addition to the infected cell, it may reduce the spread of infection.
Introduction
N
The virulence of S. Typhi is thought to be increased by cdt's role in blocking cell division, as well as directly killing epithelial and immune cells (Eshraghi et al., 2010). In addition to having a cytotoxic effect, it has been suggested that cdt may play a role in adherence and invasion of bacteria to the host cells (Biswas et al., 2006). This toxin is widely distributed among bacterial pathogens, causing DNA damage that results in cell cycle arrest, cellular distension, and apoptosis of a broad range of mammalian cell lineages (Lara-Tejero and Galan, 2000; Lara-Tejero and Galan, 2002; Jinadasa et al., 2011). Apoptotic cell death releases apoptotic bodies that contain engulfed pathogens; these can serve as a new pathogen resource to other cells and facilitate persistent and systemic infection. The cdt is defined as a bacterial genotoxin and has been reported to be present in several food- and water-associated pathogens, including several pathotypes of Escherichia coli, and species of Campylobacter and Shigella (Jinadasa et al., 2011). Recently the presence of cdtB has been reported in some NTS serovars, including Schwarzengrund, Montevideo, Bredeney, 9,12:l,v:-, and Javiana (den Bakker et al., 2011; Mezal et al., 2013; Suez et al., 2013). The genome sequence of S. Javiana (CFSAN001992) has been closed and showed the presence of the cdtB (Allard et al., 2014). Moreover, the presence of insertion sequences and prophage-like elements upstream of cdt genes in several pathogens, as well as the fact that cdt is detected in several bacterial genera, strongly suggests that the genetic determinant of cdt might be transferred horizontally among bacterial species (Peres et al., 1997) and thus raises public health concerns.
Due to the emergence of NTS-related foodborne infections and outbreaks, regulatory agencies have paid special attention in order to identify the sources through epidemiological analysis, as well as tracking and typing methods (Johnson et al., 2011). On the global level, Salmonella infection caused by NTS ranges from 200 million to 1.3 billion cases, with an estimated 3 million deaths each year (Coburn et al., 2007). The aim of this study was to identify the cdtB gene in NTS serovars isolated from food and further address the mechanism of cell death during host–pathogen interaction. To accomplish this, several NTS strains (isolated from food) were screened for the presence of the cdtB gene. Out of three cdtB-positive NTS isolated from food, only S. Javiana showed the characteristic authophagic vacuole formation in macrophages. We created a deletion mutant of cdtB in wild-type S. Javiana (cdtB+ wt S. Javiana). A systematic study was conducted to delineate the effect of cdtB on the adherence, invasion, and persistence in macrophages during in vitro infection with cdtB+wt S. Javiana, cdtB-negative NTS serovar (S. Typhimurium), or cdtB-deletion mutant of S. Javiana (cdtB− ΔS. Javiana). Further, this study describes the involvement of specific genes involved in cytotoxicity caused by cdtB-positive S. Javiana.
Materials and Methods
Bacterial strains
Several NTS serovars isolated from foods were tested for the presence of virulence gene cdtB by polymerase chain reaction (PCR). The strains tested in this study are shown in Table 1. Presence of the cdtB gene in the bacterial strains was determined by PCR using the following primers: forward primer CCATGCTGCAGCTATTGTCA and reverse primer ATTTGCGTGGGTTCTGTAGG. SYBR green master mix and 250 nM of forward and reverse primers were used for real time-polymerase chain reaction on an ABI 7500. PCR amplification was conducted with an initial 10-min step at 95°C followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Positive isolates showed Ct values that crossed the threshold line at the mid-log phase of the amplification plot. The negative isolate did not show any fluorescence above the threshold line (Khare et al., 2004).
Three different isolates of Salmonella Typhimurium.
cdtB, cytolethal distending toxin B.
Construction of the cdtB deletion mutant
The cdtB-deleted mutants (ΔcdtB::kan) of S. Javiana isolate 1300F were constructed as described earlier (Datsenko and Wanner, 2000). Briefly, a kanamycin resistance gene on pKD4 was amplified using a primer set (cdtBdel-kanF, 5′-ACTACAAAGTTATGACCTGGAATCTTCAGGGCTCTTCAGTGT-AGGCTGGAGCTGCTTC-3′; cdtBdel-kanR, 5′-AATAAGGTGCTCGATCG-ACAATGACCCCATAATCTACATATGAATATCCTCCTTAG-3′). The PCR products were treated with DpnI (New England Biolabs, Ipswich, MA) and purified by either using QIAquick PCR purification kit or gel purified using QIAquick gel extraction kit. Overnight inocula of Salmonella isolates carrying pKD46 were diluted to 1:100 in Luria-Bertani (LB) broth containing 100 μg/mL ampicillin and incubated at 30°C with agitation for 3 h. L-arabinose was added to inocula to achieve the final concentration of 10 mM and incubated at 30°C with agitation for 1 h. Cells were harvested and washed three times with ice-cold 10% glycerol. Cell pellets were resuspended in 1:100 volume of ice-cold 10% glycerol. Approximately 400 to 800 ng of purified amplicons was electroporated into the cells using Gene Pulser® Cuvette, 0.2 cm, and Gene Pulser Xcell™. Electroporated cells were immediately mixed with LB broth and incubated at 37°C, and shaking at 200 rpm for 1 h. Cells were plated on LB agar plates containing 50 μg/mL kanamycin and incubated overnight at 37°C. Colonies were isolated and screened for the presence of kanamycin resistance gene and the absence of cdtB gene by PCR-based methods (Skyberg et al., 2006).
DNase activity
DNase test agar was used to determine the DNase activity of cdtB+wt S. Javiana and cdtB− ΔS. Javiana. Staphylococcus aureus and Klebsiella pneumoniae were used as the positive and negative controls for DNase activity test (Jeffries et al., 1957). Plates were streaked with the test inoculum, and incubated at 37°C for 24 h. After 24 h, plates were flooded with 1 N HCl. DNase-positive organisms showed a clearance around the streak.
Cell culture
Mouse macrophage cells (J774 A.1, TIB-67; American Type Tissue Culture, Manassas, VA) were cultured in the tissue culture flask, which contained Dulbecco's modified Eagle medium (DMEM; Life Technologies, Grand Island, NY) supplemented with 10% fetal bovine serum, L-glutamine, sodium bicarbonate, and antibiotic solution at 37°C with 5% CO2. Upon confluence, cells were seeded in 24-well tissue culture plates at a density of 4×105 cells per well and incubated for 48 h at 37°C in the presence of 5% CO2.
Adherence, invasion, and persistence of bacterial strains in the macrophages
S. Typhimurium, S. Schwarzengrund, S. Aberdeen, cdtB+wt S. Javiana, and cdtB− ΔS. Javiana were cultured overnight in LB broth at 37°C. On the day of infection, bacteria were subcultured, harvested upon reaching an OD 600=0.600–0.800, and were resuspended in culture media for infection (multiplicity of infection=1:200) with a slight modification in the method described earlier (Raffatellu et al., 2005; Gokulan et al., 2013; Han et al., 2013). In brief, bacteria were added to the macrophage monolayer (three sets of macrophages in duplicate for adherence, invasion, and persistence). The plates were centrifuged at 500 rpm for 5 min. All plates were incubated for 1 h for the macrophage invasion to occur; then, cell monolayer was washed three times with prewarmed phosphate-buffered saline (PBS). Total cell-associated (Set 1=invading plus surface-adherent) bacteria were quantified by lysing first set of duplicate wells using 1% Triton X100; cell lysate was collected, diluted in PBS, and plated onto the LB agar plate for overnight growth. For the invasion assay, a second set of duplicate wells (bacteria removed and washed as described earlier) were treated with gentamicin (50 μg/mL) for 1 h and incubated at 37°C with 5% CO2. After 1 h, cells were washed, lysed, and plated on LB agar plate as described earlier (Set 2). For the persistence assay, a third set of cells were incubated with gentamicin (50 μg/mL) for 24 h at 37°C with 5% CO2. After 24 h, cells were washed and lysed as described earlier (Set 3). The calculation of colony-forming units (CFUs) adhered to macrophages was done as follows: CFU count from the Set 2 was subtracted from CFU counts obtained from Set 1.
The percent of bacteria adhered to the macrophages was calculated by number of CFUs adhered to macrophages divided by total CFU counts obtained after the macrophages were lysed immediately without gentamicin incubation step and this end product was multiplied by 100. Bacterial invasion and persistence were calculated by counting the CFU count of experiment Sets 2 and 3, respectively. All assays were performed in duplicate and repeated at least three times.
Cytotoxicity and apoptosis assays
The cytotoxicity and apoptosis assays were performed using ApoTox-Glo Assay from Promega (Madison, WI). Cells (20,000) were seeded in 96-well plates. After an overnight incubation, cells were infected as described earlier with S. Typhimurium, cdtB+wt S. Javiana, and cdtB− ΔS. Javiana. Cytotoxicity and apoptotic activity were assessed in separate wells at 5 and 60 min after infection as suggested by the supplier. In brief, the cytotoxicity reagent containing bis-AAF-R110 substrate was added to the cells. After 30-min incubation at 37°C, fluorescence was recorded at 485 nm excitation/520 nm emission for cytotoxicity by using a microplate reader for fluorescence (Synergy 2, Winooski, VT). Caspase-Glo 3/7 Reagent was further added to the cells and briefly mixed by orbital shaking (1300–1500 rpm for ∼30 s). After 30 min of incubation at room temperature, luminescence (caspase activation, a hallmark of apoptosis) was recorded. Results for apoptosis were expressed as percent changes in luminescence in comparison to control uninfected cells. All assays were performed in duplicate, and repeated at least three times. Statistical significance was calculated using “unpaired t-test” by GraphPad Software.
Microscopic evaluation of infected macrophages
Another set of cells from the above experiments (1 and 24 h postinfection in 24-well format) were washed with PBS and then fixed with 4% paraformaldehyde. The cells were examined using the Nikon Eclipse TS100 microscope, and micrographs were analyzed using image analysis software NIS Elements BR™. Emphasis was given to changes in the cellular and cytoplasmic distension of macrophages infected with cdtB-positive and -negative NTS serovar.
Visualization of autophagy with acridine orange
Macrophages were seeded onto 24-well cell culture plates at a density of 4×105 cells per well and incubated for 24 h. After incubation, cells were either not infected (served as control) or infected as previously described with cdtB+wt S. Javiana, cdtB− ΔS. Javiana, and S. Typhimurium. Staining for autophagy was performed at 1 and 24 h postinfection in separate sets of wells. After a 1-h incubation period, media were removed and cells were washed once with plain media. Media for all wells were then replaced with cell culture media containing gentamicin (50 μg/mL). Macrophages were stained for detection of autophagy according to the protocol developed by Arthur et al. (2007). Acridine orange dye was added to the media for a total concentration of 1 μg/mL and cells were incubated for 15 min. Staining was visualized and pictures were taken using an inverted fluorescence microscope (Advanced Microscopy Group, Bothell, WA).
Immunofluorescence detection of LC3 protein
J774A.1 mouse macrophage-like cells were plated onto cell-culture-treated Thermanox coverslips (Nunc, Rochester, NY) and incubated for 24 h. Infection of cells with Salmonella strains (cdtB+ wt S. Javiana, cdtB− ΔS. Javiana, and S. Typhimurium) and gentamicin (50 μg/mL) treatment were completed as described earlier in this section. Antibody staining of intracellular LC3 was performed using a modified method of Nicola et al. (2012). Briefly, infected cells and controls were fixed and permeabilized with methanol at −20°C for 10 min. Cells were washed, blocked with 3% milk powder, and incubated with a rabbit polyclonal anti-LC3 antibody (1:200; AbD Serotec, Raleigh, NC). After washing, cells were incubated with a goat anti-rabbit fluorescein-conjugated secondary antibody (1:1000; Molecular Probes, Eugene, OR). Coverslips were then mounted on glass slides and imaged with the EVOS Cell Imaging System (Life Technologies, Carlsbad, CA).
Determination of gene expression in macrophages
RNA extraction and reverse transcription
RNA extraction from the control and infected cells was performed using the TRI reagent following the recommended protocol from the manufacturer (Molecular Research Center, Cincinnati, OH) and as described earlier (Khare et al., 2012). The final RNA pellet was resuspended in DNase- and RNase-free water (Ambion, Austin, TX). Contaminated genomic DNA was removed by RNase-free DNase I treatment according to the manufacturer's instructions (DNA-free; Ambion), and samples were stored at −80°C until used. The concentration of RNA was quantified by measuring absorbance at λ260 nm using a NanoDrop® ND-1000 (NanoDrop, Wilmington, DE). RNA was reverse transcribed to make cDNA using the Two-Step Reverse Transcription-TaqMan kit (Applied Biosystems, Foster City, CA).
Host response during invasion and persistence of cdtB+wt S. Javiana, S. Typhimurium, and cdtB−ΔS. Javiana with a focus on the genes involved in the cell death
Gene expression of 84 cell-death-related genes was examined using mouse Cell Death RT2 Profiler PCR array (SABioscience, Valencia, CA). In brief, a total volume of 25 μL of PCR mixture, which included 12.5 μL of RT2 Real-Time SYBR Green/ROX PCR master mix and 12.5 μL of DNase- and RNase-free water and template, was pipetted in each well of the PCR array. The PCR amplification was conducted with an initial 10-min step at 95°C followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Data were imported into an Excel spread sheet and analyzed using the comparative cycle threshold method with normalization of the raw data to housekeeping gene (glyceraldehyde 3-phosphate dehydrogenase). For calculation of quantitative real-time PCR, data were analyzed by using the ddCt method (Applied Biosystems Prism 7700 Users bulletin Number 2). Data from control uninfected macrophages were compared with data from macrophages infected with cdtB+wt S. Javiana, S. Typhimurium, and cdtB− ΔS. Javiana.
Data analysis and statistics
Adhesion, invasion, and persistence of bacteria in the macrophages were considered to be positive when CFUs were detected by bacteriological culture on the LB agar plates. Cellular burden (invasion and persistence) was defined as the number of CFUs per well. The statistical significance of differences was calculated using two-tailed Student's t-test.
For the calculation of quantitative real-time PCR, data were analyzed by using the ddCt method (Applied Biosystems Prism 7700 Users bulletin Number 2). Statistical significance was determined by Student's t-test and expression differences were considered significant when p<0.05.
Results
Presence of cdtB gene in few NTS serovars as detected by PCR
Few NTS serovars, for example, Schwarzengrund, Aberdeen, and Javiana, isolated from the food were positive for the cdtB gene, whereas other NTS serovars were negative for the cdtB gene (Table 1).
Selection of wild-type S. Javiana to construct a cdtB mutant
S. Javiana was the only cdtB-positive serovar among the most frequently encountered NTS serovars (Jackson et al., 2013). For this reason, S. Javiana was chosen to assess the role of cdtB in NTS. To achieve this, a deletion mutant was constructed in wild-type S. Javiana to assess the role of cdtB gene in the S. Javiana pathogenesis. To assess whether the deletion of cdtB altered the stability of pltA and pltB genes, PCR was performed for the presence of these genes in cdtB+wt S. Javiana and cdtB−
ΔS. Javiana (please see the Supplementary Fig. S1A for the primer location; Supplementary Data are available online at
Adherence, invasion, and persistence of cdtB-positive NTS in the macrophages
The macrophage interaction with cdtB-positive NTS (S. Schwarzengrund, S. Aberdeen, and S. Javiana) was compared with cdtB− ΔS. Javiana using invasion and persistence assays. A well-studied NTS serovar S. Typhimurium (lacks cdtB gene) was used as an experimental control. This study was initiated to conduct experimentation in nonhuman cell lines because S. Javiana is an enteric (zoonotic) pathogen and causes infections via consumption of meat or fresh-cut produce (Jackson et al., 2013) or amphibian contact (Srikantiah et al., 2004; Clarkson et al., 2010). For infection in humans, S. Javiana must first be able to infect and persist in a nonhuman host to facilitate foodborne/contact transmission. As expected, S. Typhimurium was more invasive (p<0.001) as compared with cdtB-positive serovars (Fig. 1A). The cdtB− ΔS. Javiana was slightly more invasive than the wild-type serovar; however, this difference in the invasion was not statistically significant. Persistence of S. Typhimurium was also significantly higher as compared with cdtB-positive NTS serovars. Next, we checked whether there was a difference in the adherence of bacteria to macrophages due to the presence of cdtB gene. The percentage of cdtB− ΔS. Javiana adhered to macrophages was significantly higher as compared with cdtB+wt S. Javiana (Fig. 1B). Similar results were also found when macrophages were infected with S. Typhimurium, in which about 50% of adhered, invasive bacterial population was due to the adhered bacterial population on the macrophages.
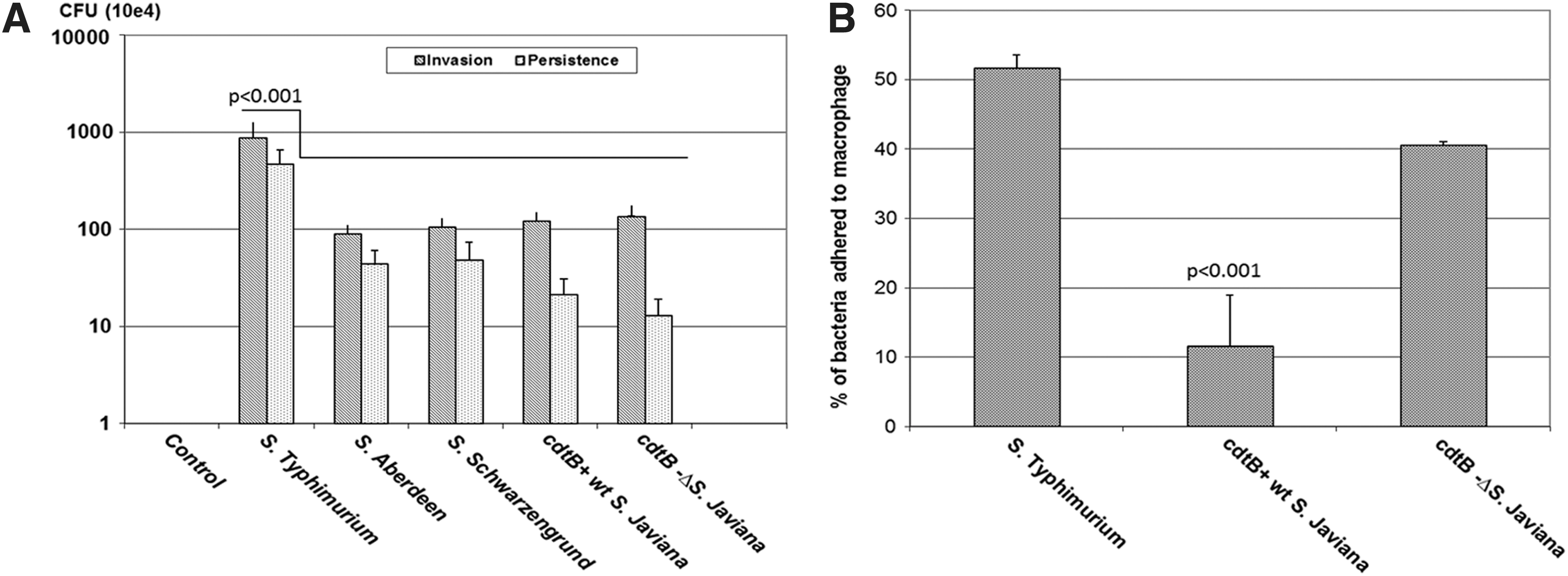
Invasion (1 h postinfection) and persistence (24 h postinfection) of NTS serovars during macrophage infection
Cytotoxicity and caspase activation during interaction of cdtB-positive S. Javiana with macrophages
The fluorescence and luminescence assays clearly showed a distinct pattern of cytotoxicity and apoptosis, respectively, during S. Typhimurium and S. Javiana infection. The S. Typhimurium showed significantly increased level of cytotoxicity (p<0.0002) in infected cells as compared with control uninfected cells (Fig. 2A). Both cdtB+wt S. Javiana and cdtB− ΔS. Javiana showed significant changes in cytotoxicity as compared with control uninfected cells (p<0.02 and p<0.0002, respectively) as soon as the pathogen entered the macrophages (5 min postinfection). However, differences in cytotoxicity exerted by cdtB+wt S. Javiana and cdtB− ΔS. Javiana were not statistically significant (p<0.09) at 5 min postinfection. On the other hand, at 60 min postinfection, cytotoxicity exerted by cdtB+wt S. Javiana was significantly higher as compared with cdtB− ΔS. Javiana (statistical significance p<0.0002).

Cytotoxicity and apoptosis of macrophages caused by NTS strains
The luminescence intensity is directly proportional to the amount of caspase 3/7 activity. All infected macrophages (60 min postinfection with S. Typhimurium, cdtB+wt S. Javiana, and cdtB− ΔS. Javiana) showed significant increase in caspase activity as compared with control uninfected cells (Fig. 2B). The caspase activity was significantly higher (p<0.0001) in the cells infected with S. Typhimurium as compared with cdtB+ wt S. Javiana and cdtB− ΔS. Javiana.
Morphological changes during interaction of cdtB+wt S. Javiana, S. Typhimurium, and cdtB− ΔS. Javiana
The uninfected macrophages showed a well-defined round structure, with a clear cell wall (Fig. 3A, B). Infection of macrophages with NTS serovars resulted in different kinds of morphological changes; S. Typhimurium caused the lysis of cells within 1 h of infection, which increased significantly after 24 h of infection (Fig. 3C, D, respectively). In comparison, cdtB+wt S. Javiana infection of macrophages showed changes in the morphology of the macrophages, which was evident by the elongation and distended shape of macrophages, within an hour of infection (Fig. 3E). After 24 h, cdtB+wt S. Javiana-infected macrophages showed severe vacuolization (Fig. 3F). The cdtB− ΔS. Javiana showed almost the same characteristics as S. Typhimurium; however, the lysis of cells was not as prominent (Fig. 3G, H).
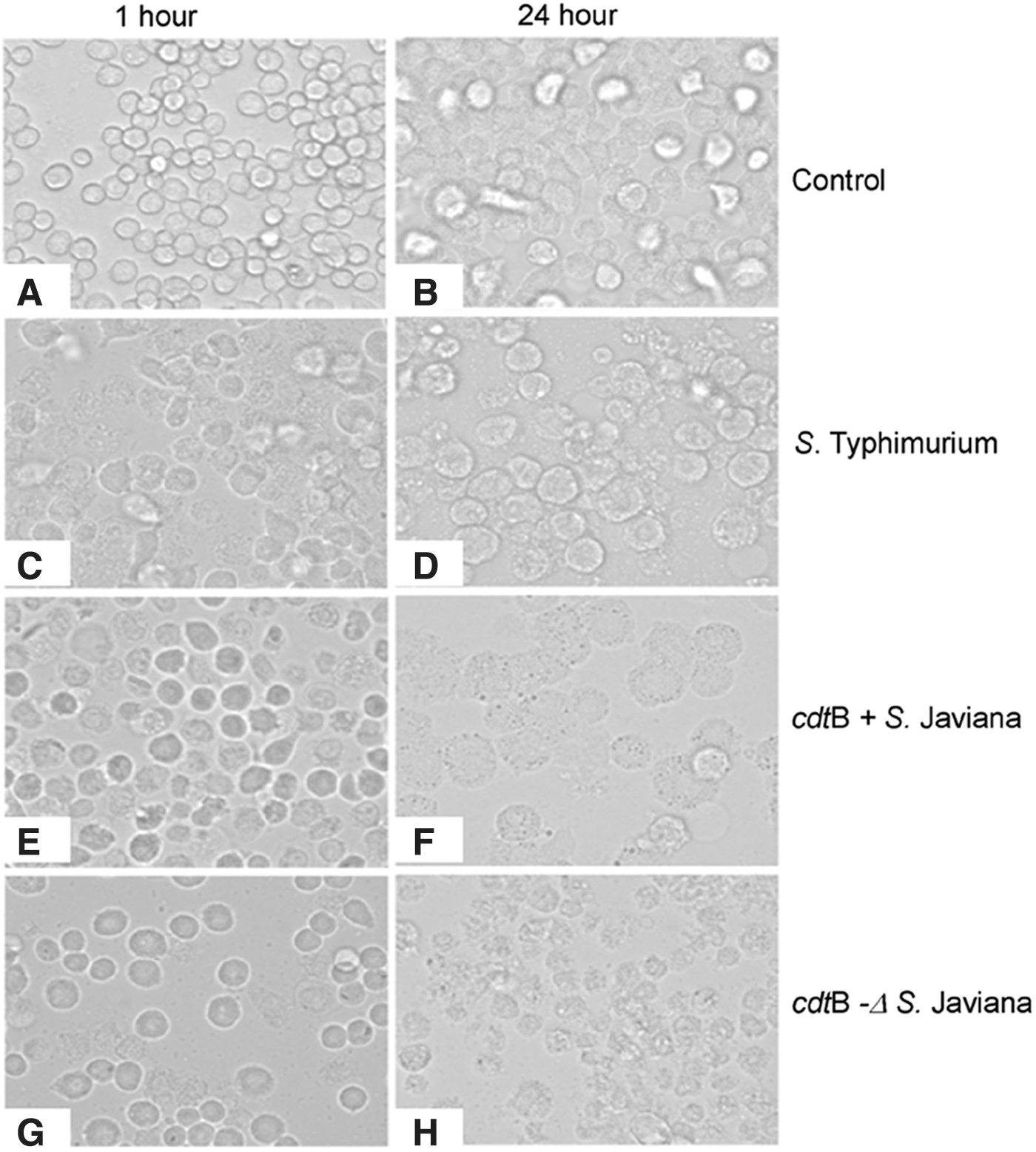
Changes in the morphology of macrophages due to infection with NTS strains at 1 h (left panel
Detection of autophagy with acridine orange staining
Acridine orange staining was used to visualize autophagic compartment, as an indicator of autophagic activity in macrophages during infection with S. Typhimurium, cdtB+wt S. Javiana, and cdtB− ΔS. Javiana. At 60 min postinfection, differences between infected groups were not significant (data not shown). The control cells did not show any staining with acridine orange at 24 h (Fig. 4A). However, 24 h postinfection, cdtB− ΔS. Javiana showed a little staining of acridine orange (Fig. 4B), whereas cdtB+wt S. Javiana displayed increased cell distension and vacuolization, the hallmark of autophagy (Fig. 4C). Visualization of S. Typhimurium at 24 h revealed that most of the cells had been lysed, with only a few live cells remaining (Fig. 4D). These results provide evidence that the mode of cell death is different in S. Typhimurium and S. Javiana. Given autophagy's known role in the destruction of intracellular pathogens within the cell, this observation supports the assertion that the wild-type cdtB+ wt S. Javiana is contained in the macrophages. Additionally, these results provide further evidence that autophagy may be one of the predominant cell death mechanisms for macrophages infected with cdtB+wt S. Javiana.
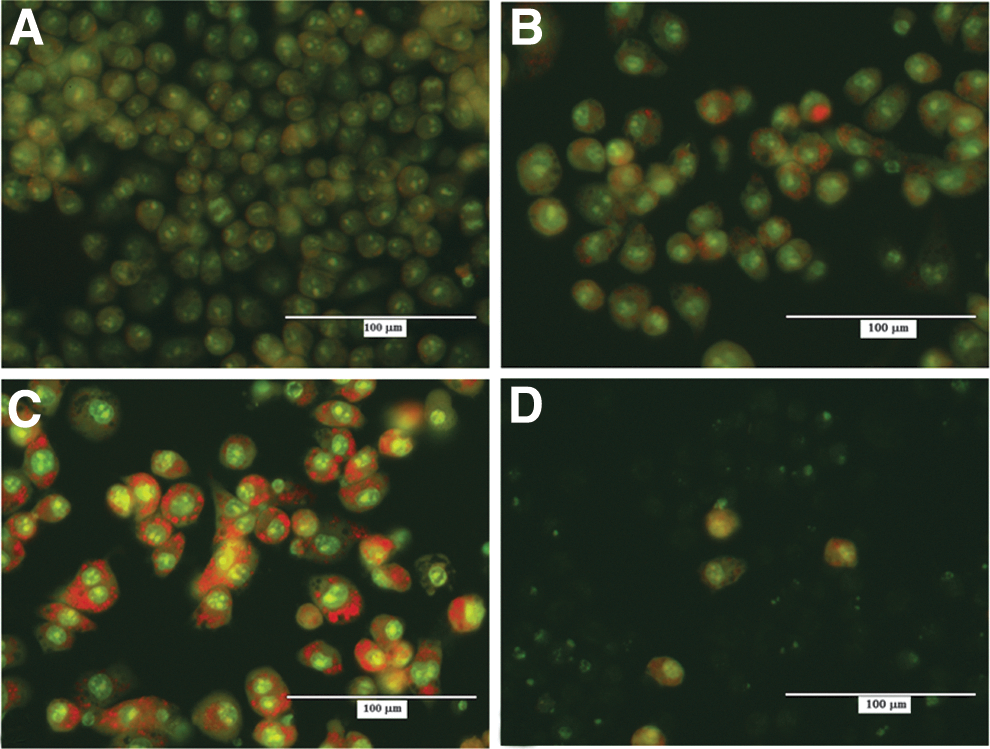
Acridine orange staining to determine autophagy in infected macrophages. Acridine orange staining was used to visualize autophagy in macrophages after 24 h of infection with cdtB−
ΔS. Javiana
Immunofluorescence detection of LC3 protein
The immunofluorescence visualization of LC3 (microtubule-associated protein 1 light chain 3 alpha) clearly shows that the autophagosome marker is present in the macrophages infected with cdtB+wt S. Javiana (Fig. 5C). Further, as observed earlier, these cells also showed significant cell distension and vacuolization. In contrast, presence of the LC3 protein was substantially reduced in macrophages that were infected with cdtB− ΔS. Javiana or S. Typhimurium (Fig. 5B, D).
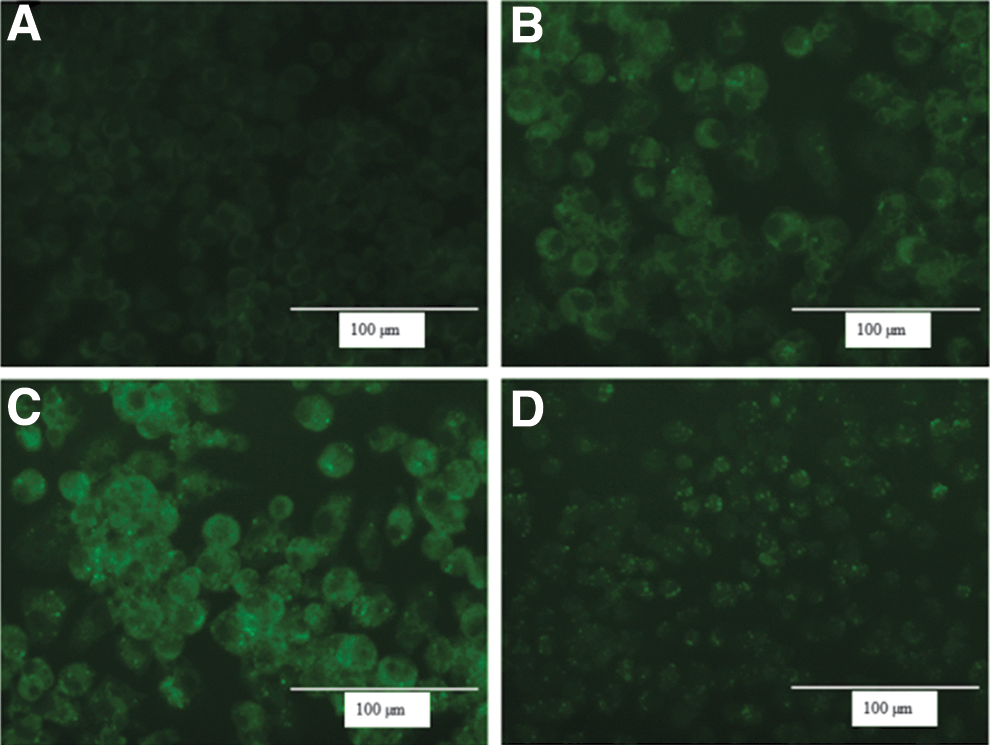
Immunofluorescence visualization of LC3 protein in macrophages in
Host response during invasion and persistence with a focus on the genes involved in the cell death
The cell death pathway array was probed to identify the cell death mechanism involved in necrosis, apoptosis, and autophagy during interaction of cdtB with macrophages after 24 h of infection. Here, our focus was to emphasize the uniquely expressed genes during infection of macrophages with S. Typhimurium, cdtB+wt S. Javiana, and cdtB− ΔS. Javiana. Macrophages infected with cdtB+wt S. Javiana did not show any increase in antiapoptotic or proapoptotic genes as compared with S. Typhimurium (Fig. 6A, B); however, deletion of cdtB gene resulted in higher expression of Caspase 2. Macrophages infected with cdtB+wt S. Javiana showed a slight increased expression of genes involved in necrosis (coiled-coil domain-containing protein 103 [CCDC103] and Voltage gated potassium channel interacting protein 1 [Kcnip1]) and apoptosis (Bcl-2–associated X protein [Bax] and cylindromatosis [Cyld]) (Fig. 6C, D). There was a significant increase in the expression of genes involved in autophagy during macrophage infection with cdtB+wt S. Javiana as compared with S. Typhimurium and cdtB− ΔS. Javiana (Fig. 6E, F). The examples of significantly upregulated genes are Estrogen receptors 1 (ESR1) and Phosphatidylinositol 3-kinase catalytic subunit type 3 (PIK3C3). Interestingly, Microtubule-associated proteins 1A/1B light chain 3A (Map1lc3a) gene (a mammalian homolog of ATG and a key marker for autophagy) was highly upregulated in macrophages infected with S. Javiana (both cdtB positive and mutant strains) as compared with macrophages infected with S. Typhimurium. Conversely, during interaction of S. Typhimurium with macrophages, higher expression of tumor necrosis factor receptor superfamily, member 11b (antiapoptotic gene), caspase 6 gene (proapoptotic gene), and thioredoxin-like 4B (txnl4b; necrosis gene) was found, as compared with cdtB− ΔS. Javiana and cdtB+ ΔS. Javiana. The cdtB− ΔS. Javiana-infected macrophages showed an overlapping pattern for gene expression associated with cell death (apoptosis, necrosis, and autophagy); some overlapped with S. Typhimurium and others overlapped with cdtB+wt S. Javiana. However, some genes were uniquely upregulated in cdtB− ΔS. Javiana-infected macrophages and they were Caspase 2 (antiapoptotic gene), 9430015G10RiK (necrosis), and Spata2 (apoptosis).

Expression of genes involved in cell death during interaction of macrophages with NTS strains. Figure shows the fold change in the expression of genes involved in antiapoptotic
Discussion
The ability of Salmonella to persist within the host depends on a balance between immune responses that lead to the clearance of the pathogen and avoidance of damage to host tissues (Ruby et al., 2012). Usually the estimated numbers of cases of NTS are much higher than S. Typhi. However, the proportions of hospitalized cases for S. Typhi are much higher than for the NTS (75% vs. 1.1%, respectively, in the United States). This may be due to the more typically persistent and systemic infection caused by this pathogen. This study was carried out to investigate the presence of the cdtB gene in the NTS serovars. The aim of this study was twofold; first, the outcome of screening for cdtB in NTS would reveal whether this unique virulence gene was present in the NTS, and, second, to delineate the mechanism of cdtB+wt S. Javiana interaction with macrophages. Salmonella pathogenesis requires the coordinated expression of virulence factors that allow the bacterium to invade the host cells and evade the host's immune system. Once inside the macrophages, the bacteria replicate and ultimately burst the host cells. The released bacteria start a new phase of bacterial internalization and intracellular replication.
Three NTS serovars (out of 17 serovars) showed the presence of the cdtB gene, and, out of three cdtB-positive NTS, only S. Javiana showed the characteristic autophagic vacuole formation in the macrophages. According to the Foodborne Diseases Active Surveillance Network, S. Javiana is reported to be a highly prevalent NTS strain for number and incidences of infections caused by NTS (2012). The cdtB+wt S. Javiana also contains pltA and pltB genes. These genes (pltA and pltB) form a complex with cdtB and induce toxicity to the target cells (Song et al., 2013). Thus, these results raise the question whether the pathogenesis of the cdtB+wt S. Javiana is similar to that of S. Typhi and has the capability for extended persistence in the host. To understand the role of cdtB during early pathogenesis of cdtB+wt S. Javiana, we created a cdtB mutant of S. Javiana (cdtB− ΔS. Javiana) and extended the study to understand whether there is a difference in the interaction of cdtB+wt S. Javiana, S. Typhimurium, and cdtB− ΔS. Javiana with the macrophages. Our results showed that (1) bacteria that do not possess cdtB adhered more to macrophages, and (2) the presence of the cdtB gene probably does not facilitate invasion of bacteria in the macrophages. Kalischuk et al. (2007) also showed that invasion of cdtB-positive Campylobacter jejuni was independent of cdt. On the other hand, Haghjoo and Galan (2004) showed that cdtB-positive bacteria have an increased capability to internalize within the host cells. Based on our results, we can speculate that due to the lack of cdtB, the cdtB− ΔS. Javiana might have a slower rate of internalization/invasion; however, they adhered to the cell surface more efficiently as compared with cdtB+wt S. Javiana. Ideally, the increased adhesion should result in increased invasion or bacterial recovery after 24 h. This is only possible in the absence of any antibiotic. However, it is important to note for these studies (adhesion, invasion, and persistence experiments) that after the 1 h of invasion and adhesion, the extracellular bacteria were killed by gentamycin treatment. Further, for the persistence assay, after removal of nonadherent bacteria (after 1 h of infection) the cells were cultured in the gentamycin-containing media. In this situation all the bacteria that adhered to macrophages will be killed and thus would not contribute for the increased bacterial recovery after 24 h.
The cytotoxic mechanisms of cell death involve several processes, including necrosis, apoptosis, and autophagy. During the pathogen interaction with a host cell, necrosis and apoptotic cell death are considered cytotoxic; however, the autophagic process of cell death is considered cytoprotective (Rikiishi, 2012). We extended our study to distinguish the cell death mechanisms in macrophages during infection with S. Typhimurium, cdtB−ΔS. Javiana, and cdtB+wt S. Javiana. Macrophages infected with S. Typhimurium showed a significantly increased level of cytotoxicity in the biochemical assay, as well as cytoplasmic swelling and rupture of the plasma membrane during morphological assessment, all of which are hallmarks of necrotic cell death. Further, a significant increase in caspase 3/7 activity may reflect the induction of cell death by apoptosis. This observation is in agreement with the earlier studies that showed that the S. Typhimurium invades macrophages and the effector molecule SipB secreted by SPI-1 activates the secretion of proinflammatory cytokines and induction of apoptosis (van der Velden et al., 2000).
This interaction of macrophage and pathogen results in the escape of S. Typhimurium, which is taken up by new macrophages and renews the replication cycle (van der Velden et al., 2000). In contrast, the cdtB+wt S. Javiana showed the lowest cytotoxicity at the start of the infection (5 min postinfection). This initial period may reflect the initiation of cell cycle arrest with no parallel increase in cytotoxicity. However, engulfed bacteria were associated with an increase in the cytotoxicity at 60 min postinfection. The morphological analysis of cdtB+wt S. Javiana-infected macrophages indicated signs of autophagic vacuole formation. Moreover, the delay in the cytotoxicity caused by cdtB+wt S. Javiana may be due to the initiation of the autophagy process. The results of staining with acridine orange provided further evidence that autophagy is one of the primary cell death mechanisms of macrophages infected with cdtB+wt S. Javiana. At 24 h postinfection, a majority of macrophages infected with cdtB+wt S. Javiana exhibited the bright red vacuole staining and LC3 protein activation, both characteristic of autophagy. The autophagic pathway is a pathogen-driven process, and starts with the internalization of bacteria in autophagic vacuoles that may serve as a replicative niche for the pathogen by delaying its fusion with lysosome (Colombo, 2007). The process of autophagy is completed with the formation of large double-membrane vesicles, called autophagosomes, which encapsulate the cytoplasm and organelles and then fuse with lysosomes, thus degrading the contents of vesicles. In the host cells, autophagy prevents the accumulation of damaged organelles and invading microorganisms at the site of infection. Higher occurrence of the autophagosome marker LC3 in the macrophages infected with cdtB+wt S. Javiana demonstrates the role of cdtB in macrophage autophagy. This phenomenon could be extrapolated in the in vivo situation as restricting the intracellular survival and spread of cdtB+wt S. Javiana.
In addition to staining with acridine orange alone, cells were also stained with ethidium bromide to visualize necrotic cells in addition to those undergoing autophagy (data not included in the article). The images stained with ethidium bromide indicate that necrosis, while present in cdtB+wt S. Javiana-infected cells, is not a major cause of cell death. The cellular morphology clearly indicated differences in the host cells infected with cdtB+wt S. Javiana, cdtB− ΔS. Javiana, and S. Typhimurium. Image analysis suggested that ∼12–13% of the total cells showed signs of necrosis after infection with S. Javiana. To further delineate the cell death mechanism of cdtB-positive and -negative NTS, transcriptional analysis of genes involved in necrosis, apoptosis, and autophagy was conducted in detail. As expected, there was a cumulative effect of cell death due to necrosis, apoptosis, and autophagy in all the pathogens tested in this study. However, there was predominance of one type of cell death (evident due to the expression of genes involved in apoptosis, necrosis, or autophagy) to the other types, during infection with different bacteria. In fact, the significant overexpression of ESR1, PIK3C3, and Map1lc3a genes in cdtB+wt S. Javiana-infected macrophages, which are involved in the autophagic process, clearly supports that cdtB-mediated cytotoxicity is due, at least in part, to autophagy (Chung and Van Hul, 2012; Jaber et al., 2012; Loria and Graf, 2012). Autophagy has also been defined as a protective intracellular pathway against secreted bacterial toxin (Gutierrez et al., 2007).
Overall, these results led us to postulate that the activation of the autophagic process in infected macrophages could be influenced by the presence of cdtB. The cytotoxicity, morphological analysis, and gene expression data of macrophages infected with cdtB+wt S. Javiana support the assertion that autophagy may be a predominant mechanism of host cell death, though there was some contribution of necrosis and apoptosis as well. The cdtB-induced upregulation of Kcnip1 suggests that cdtB may partially relate to the Taiwan cobra cardiotoxin3 (CTX3), which also interacts with Kcnip1 (Lin et al., 2004). The cdtB+wt S. Javiana-infected macrophages also showed significant increase in the expression of Bax and Cyld genes that are involved in the crosstalk between apoptosis and autophagy. Upregulation of Bax is associated with promotion of apoptosis by competing with Bcl-2 proper (Souvannavong et al., 2007; Juhasova et al., 2011). The cyld gene is a deubiquitinating enzyme and has been identified as a key negative regulator for NF-kappaB-dependent induction of proinflammatory mediators in phagocytic cells (Tsagaratou et al., 2011; Wang et al., 2012). Significantly higher expression of cyld gene in cdtB+wt S. Javiana-infected macrophages may be a host-protective mechanism in which bacteria-induced negative-feedback regulation plays a major role in preventing detrimental inflammatory responses. Further, this gene has been shown as a regulator of necroptosis (Bonapace et al., 2010).
An interesting difference between cdtB− ΔS. Javiana and cdtB+ S. Javiana infections was the higher expression of Atg3 and Spata2. Atg3 expression was higher when macrophages were infected in the absence of cdtB gene (cdtB− ΔS. Javiana and S. Typhimurium). Earlier, it was shown that Atg3 is required for S. Typhimurium-mediated autophagy (Huett et al., 2009). The role of Spata2 is not very clear in the cell death; it is a coregulator of apoptosis and autophagy and the molecular mechanism of action of this gene is still under investigation (Maran et al., 2009).
Conclusions
This study shows clear differences in the outcome of macrophage infection with cdtB+wt S. Javiana and cdtB− ΔS. Javiana at cell morphology, mechanism of cell death, and gene transcription level. In conclusion, several cell death pathways (apoptosis, necrosis, and autophagy) were identified in macrophages infected with cdtB+wt S. Javiana. However, our work provides evidence that autophagy was a major contributor toward macrophage death by cdtB+wt S. Javiana. As the autophagic process is known to kill the intracellular pathogen and digest the byproducts, we propose that infection with S. Javiana may be less likely to lead to systemic and persistent infection. Further in vivo experiments are needed to confirm this hypothesis.
Footnotes
Acknowledgments
Ms. Katherine Williams, Ms. Diamond Shelman, and Mr. Tatsuya Akiyama were supported by Oak Ridge Institute for Science and Education. Dr. Kuppan Gokulan was supported by FDA Commissioner Fellowship Program. The authors would like to thank Drs. Carl Cerniglia, Steven Foley, Doug Wagner, and Jyotsnabala Kanungo for their critical review of the article. The authors would like to acknowledge help provided by Dr. Dongryeoul Bae for identification of pltA and pltB genes in cdtB+wt S. Javiana and cdtB− ΔS. Javiana.
Disclosure Statement
The views expressed in this article do not necessarily reflect those of the U.S. Food and Drug Administration. Authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.