
Abstract
Adenosine monophosphate (AMP)-activated protein kinase (AMPK) plays a central role in energy homeostasis and regulation of inflammatory responses. The present study is aimed to investigate the anti-inflammatory effects of ENERGI-F704, a nucleobase analogue isolated from bamboo leaves, on expression of proinflammatory mediators in murine macrophage RAW264.7 in response to lipopolysaccharide (LPS). ENERGI-F704 enhanced phosphorylation of AMPK(T172) but insignificantly affected the viability of RAW264.7 cells. Further investigation showed that ENERGI-F704 decreased mRNA expression of interleukin (IL)-6, IL-8, tumor necrosis factor-α (TNF-α), cyclooxygenase-2 (COX2), and inducible nitric oxide synthase (iNOS) induced by LPS, as well as suppressed the production of prostaglandin E2 (PGE2) and nitric oxide (NO). Additionally, the inhibitory effects of ENERGI-F704 on the LPS-induced proinflammatory mediators were diminished by pretreatment of AMPK inhibitor Compound C. ENERGI-F704 also inhibited LPS-triggered activation of nuclear factor kappa B (NF-κB), phosphatidylinositol 3-kinase (PI3K), and p38 mitogen-activated protein kinase (p38), whereas extracellular signal-regulated kinase (Erk)1/2 and c-Jun N-terminal kinase (JNK) were insignificantly influenced. Our findings indicate that ENERGI-F704 may exert anti-inflammatory activity on RAW264.7 cells in response to LPS through the activation of AMPK and suppression of PI3K/P38/NF-κB signaling and the consequent decreased expression of proinflammatory mediators, suggesting that ENERGI-F704 is beneficial to the amelioration of inflammatory disorders.
Introduction
A
Inflammation originates from the attempt at self-protection against harmful stimuli through the activation of serial immune responses. Among the immune systems participating in host defense, macrophages are the primary cells triggered by lipopolysaccharide (LPS). LPS binds to toll-like receptor (TLR)-4 and induces secretion of a variety of proinflammatory cytokines, such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α (Raabe et al., 1998; Hsu and Wen, 2002). In addition, LPS also enhances expression of cyclooxygenase-2 (COX2) and iNOS, which produce potent inflammatory mediator prostaglandins and nitric oxide (NO) (Erwig and Rees, 1999).
Activation of the nuclear factor kappa B (NF-κB) plays a crucial role in inflammatory diseases (Tak and Firestein, 2001; Imanifooladi et al., 2010). NF-κB is a heterodimer composed of p50 and p65, which functions as a transcriptional activator to induce gene expression of proinflammatory mediators. In its inactive state, NF-κB is a cytoplasmic heterodimer that consists of three subunits: p50, p65, and inhibitor of κB (IκB). Upon stimuli such as LPS, IκBα is phosphorylated and then degraded through the ubiquitin–proteasome pathway, exposing nuclear localization signals on the p50-p65 heterodimer (Guha and Mackman, 2001).
In the present study, we investigated anti-inflammatory effects of an AMPK activator ENERGI-F704 identified from Bambusa oldhamii Munro on murine macrophage RAW264.7 in response to LPS. Cell viability was analyzed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assay. mRNA expression and protein production of proinflammatory mediators was determined by reverse transcription-polymerase chain reaction (RT-PCR), real-time (RT) quantitative PCR (qPCR), and quantitative sandwich enzyme-linked immunosorbent assay (ELISA). Kinase and NF-κB signaling cascades were investigated by immunoblotting analysis.
Materials and Methods
Chemicals and reagents
Aprotinin, leupeptin, LPS, MTT, Igepal CA-630, phenylmethylsulfonyl fluoride (PMSF), NaF, NaCl, Na2HPO4, Tris-HCl, Triton X-100, Tween-20, and Compound C (Dorsomorphin) were purchased from Sigma-Aldrich (St. Louis, MO). ENERGI-F704 was provided by Energenesis Biomedical Co. Ltd. (New Taipei City, Taiwan).
Cell culture and treatment
Murine macrophage cell line, RAW264.7, purchased from ATCC was cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Gaithersburg, MD) supplemented with 0.1% sodium bicarbonate, 2 mM glutamine, penicillin G (100 U/mL), streptomycin (100 μg/mL), and 10% fetal bovine serum and maintained at 37°C in a humidified incubator containing 5% CO2. Cells were plated at a density of 3×105 cells in 60-mm Petri dish. After incubation with ENERGI-F704 at indicated concentrations for 3 h, the RAW264.7 cells were then treated with 0.1 μg/mL LPS for 6 h (immunoblotting, RT-PCR, and RT-qPCR analysis) or 24 h (cell viability assay, protein secretion, and NO production). For AMPK inhibition, pretreatment with 5 μM AMPK inhibitor Compound C for 1 h was performed before incubation with ENERGI-F704.
Cell viability assay
Cell viability was determined by mitochondrial-dependent reduction of MTT to formazan. Ten microliters of MTT solution (5 mg/mL in DMEM) was added to the culture medium and incubated for 4 h at 37°C. After removal of medium, 2-propanol was added into Petri dish and then solubilize formazan was quantitated by optical density at 570 nm obtained from a microplate reader (Benchmark; Bio-Rad Laboratories, Hercules, CA). The optical density of formazan generated by untreated cells was regarded as 100%.
RNA extraction, RT-PCR, and RT-qPCR
After treatments, total RNA was isolated from lysed cells using the RNeasy kit according to the manufacturer's instructions (Qiagen, Valencia, CA). The purified RNA was used as a template to generate the first-strand cDNA synthesis using RevertAid™ First Strand cDNA Synthesis Kit (Fermentas Life Sciences, St. Leon-Rot, Germany). The primer sequences used for RT-PCR and qPCR were listed in Table 1. qPCR was performed using the ABI PRISM 7700 sequence detection system (Applied Biosystems, Foster City, CA). For mRNA quantitation, FastStart Universal SYBR Green Master kit (Roche Applied Science, Mannheim, Germany) was used for Taqman PCR. The threshold cycle numbers were calculated using the ΔΔCT relative value method and normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Three independent experiments were performed for RT-PCR and RT-qPCR analysis. The correct size of the PCR products was checked by agarose gel electrophoresis.
COX2, cyclooxygenase-2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IL-6, interleukin 6; IL-8, interleukin 8; iNOS, inducible nitric oxide synthase; RT-qPCR, reverse transcription real-time quantitative polymerase chain reaction; RT-PCR, reverse transcription-polymerase chain reaction; TNF-α, tumor necrosis factor-α.
Quantitation of IL-6, IL-8, TNF-α, and PGE2 using ELISA
For quantitative analysis of IL-6, IL-8, TNF-α, and prostaglandin E2 (PGE2) production, cells were seeded in six-well plates at an initial density of 5×105 cells/mL. After indicated treatments, the resulting culture supernatants were collected and the concentrations of IL-6, IL-8, TNF-α, and PGE2 were determined using Mouse Quantikine® ELISA Kits (IL-6, IL-8, TNF-α) and PGE2 Parameter Assay Kit (R&D Systems, Abingdon, United Kingdom) according to the manufacturer's instructions.
NO assay
After treatments, the culture supernatants were gently mixed with the equal volume of modified Griess reagent (Sigma-Aldrich) and then absorbance at 540 nm was measured by the microplate reader (Benchmark; Bio-Rad Laboratories). Quantitation of NO was performed by using sodium nitrite (NaNO2) as a standard (Green et al., 1982).
Immunoblotting analysis
Cells (3×106 cells) were harvested, washed twice with ice-cold PBS, and incubated with lysis buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% v/v Igepal CA-630, 1 mM PMSF, 1 mM NaF, and 10 μg/mL aprotinin and leupeptin). The cell lysates were centrifuged at 14,000 g for 10 min at 4°C to remove debris, and then, the resulting supernatants were collected. Crude protein concentrations were quantitated using BCA™ protein assay kit (Pierce, Rockford, IL) and bovine serum albumin as standard. Proteins were electrophoresed on 12.5% SDS–polyacrylamide gel and transferred onto a nitrocellulose membrane (Millipore, Bedford, MA). The blotted membrane was blocked with 5% w/v skimmed milk in phosphate-buffered saline for 1 h and then incubated with 1/1000 dilution of the primary antibodies for 2 h. The primary antibodies included antibody against phosphorylated-threonine172 AMPK [p-AMPK(T172)], AMPKα1, phosphorylated phosphatidylinositol 3-kinase (p-PI3K), total PI3K (t-PI3K), phosphorylated extracellular signal-regulated kinase 1/2 (p-Erk1/2), total Erk1/2 (t-Erk1/2), phosphorylated c-Jun N-terminal kinase (p-JNK), total JNK (t-JNK), phosphorylated p38 mitogen-activated protein kinase (p-p38), total p38 (t-p38; Cell Signaling Technologies, Danvers, MA) and GAPDH (Abcam, Cambridge, MA). Bound antibodies were detected using 1/3000 dilution of peroxidase-conjugated secondary antibodies (Sigma-Aldrich), and signals were developed using ECL chemiluminescence reagent (Millipore) as the substrate system. Quantitative analysis was performed using chemiluminescent densitometry.
Subcellular fractionation
Cells were washed with normal saline and incubated with a lysis buffer (10 mM HEPES, pH7.6; containing 15 mM KCl, 2 mM MgCl2, 0.1 mM EDTA, 1 mM dithiothreitol, 0.05% v/v Igepal CA-630 and 1 mM PMSF, 1 mM Na3VO4, 50 mM NaF, 10 μg/mL leupeptin, and 10 μg/mL aprotinin) for 10 min. After centrifugation at 2500 g for 10 min at 4°C, the supernatant was transferred into a new Eppendorf, further centrifuged at 20,000 g for 15 min at 4°C, and then the resulting supernatant was collected as cytosolic fraction. The pellets containing nuclei were washed with normal saline, incubated with a nuclear buffer (25 mM HEPES, pH7.6, 0.1% v/v Igepal CA-630, 1 M KCl, 0.1 mM EDTA, 1 mM PMSF, 1 mM Na3VO4, 2 mM NaF, 10 μg/mL leupeptin, and 10 μg/mL aprotinin), and then centrifuged at 10,000 g for 15 min at 4°C. The resulting supernatants were collected as nuclear fraction.
Statistical analysis
Data were presented as means±SD of the three to five independent experiments. Statistical comparisons are made by a one-way analysis of variance, followed by the Duncan multiple comparison test. Differences are considered to be significant when the p-values are <0.05.
Results
Effects of ENERGI-F704 on AMPK phosphorylation and viability of murine macrophage RAW264.7 cells
To examine whether ENERGI-F704 induced AMPK activation in RAW264.7 cells without or with exposure to LPS (0.1 μg/mL), the level of AMPK with phosphorylation at threonine172 was determined by immunoblotting. The results showed that both ENERGI-F704 alone or ENERGI-F704 combing with LPS dose dependently enhanced the phosphorylation of AMPK [p-AMPK(T172)] in RAW264.7 cells compared to the control (Fig. 1A). In parallel, LPS alone slightly increased the level of p-AMPK(T172).
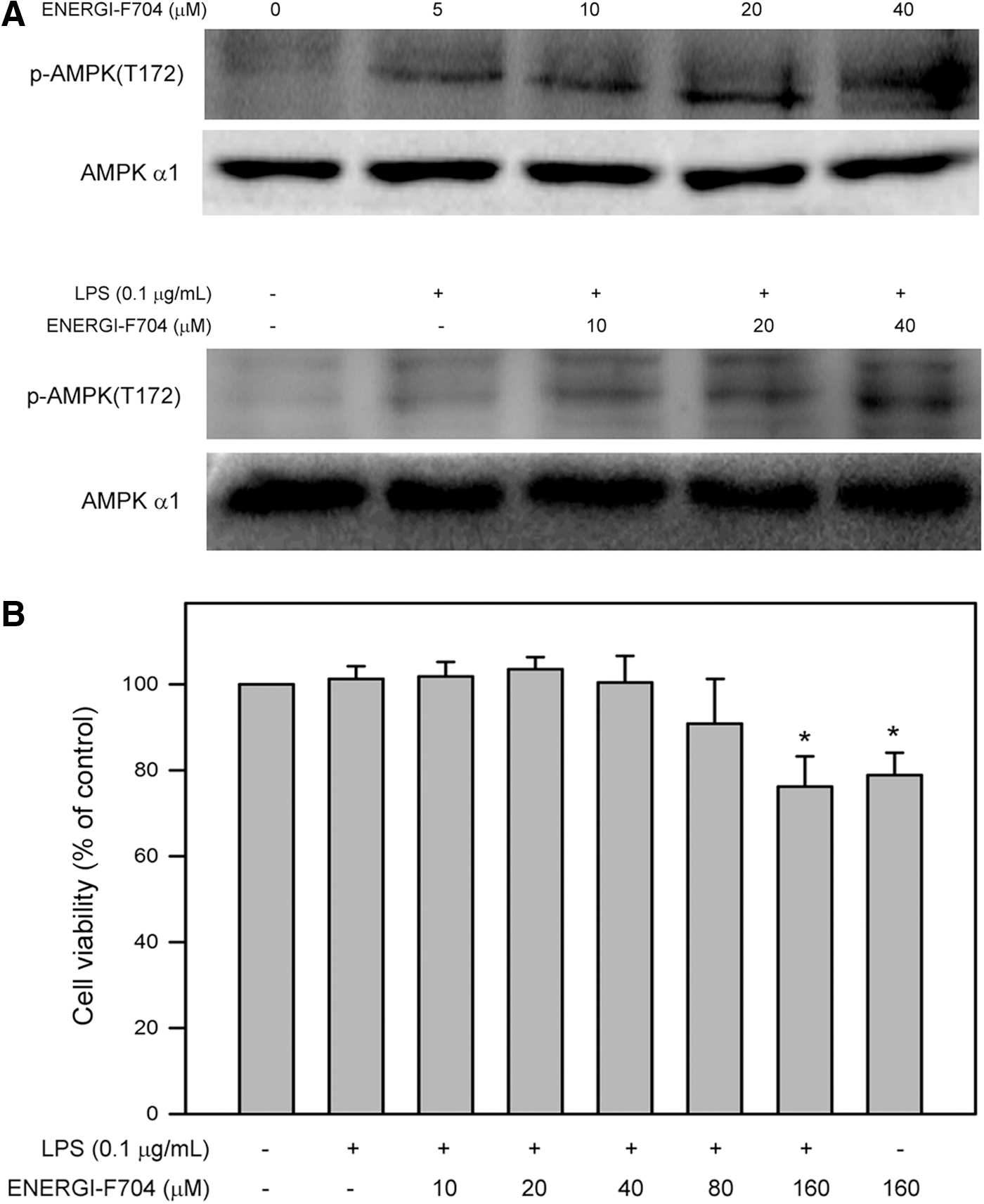
ENERGI-F704 induced AMPK phosphorylation but insignificantly affected the viability of RAW264.7 cells.
Cytotoxic effects of ENERGI-F704 on viability of RAW264.7 cells were next demonstrated. Viability of RAW264.7 cells cultured with LPS alone (0.1 μg/mL) or LPS combining with ENERGI-F704 at a serial concentration (10, 20, 40, 80, and 160 μM) for 24 h was determined using the MTT assay. As shown in Figure 1B, our findings revealed that LPS alone and LPS combining with ENERGI-F704 (10, 20, 40, and 80 μM) insignificantly affected the viability of RAW264.7 cells (p>0.05 compared to control). However, high concentration of ENERGI-F704 (160 μM) alone or combing with LPS reduced the cell viability to 78.9%±2.4% and 76.3%±4.1% of control, respectively (p<0.05 compared to control). Accordingly, ENERGI-F704 at concentrations of 20 and 40 μM were used for the subsequent investigation of anti-inflammatory activity.
ENERGI-F704 inhibited both mRNA expression and secreted protein level of IL-6, IL-8, and TNF-α in RAW264.7 cells in response to LPS
Effects of ENERGI-F704 on mRNA expression and protein production of proinflammatory cytokines in RAW2647 cells exposed to LPS were next investigated by using RT-PCR, RT-qPCR, and sandwich ELISA. RT-PCR analysis revealed that LPS alone significantly enhanced mRNA expression of IL-6, IL-8, and TNF-α, and the increased mRNA expressions were suppressed by pretreatment of ENERGI-F704 in a dose-dependent manner (Fig. 2A). Involvement of AMPK activation in ENERGI-F704-lowered mRNA expression of the proinflammatory mediators is accessed by using Compound C, a cell-permeable pyrazolopyrimidine compound that inhibits the AMPK kinase activity (IC50=234.6 nM) and is commonly used in combination with AMPK activators, such as Metformin for studying AMPK-dependent cellular events in vitro (Zhou et al., 2001). Our results revealed that the suppressive effects of ENERGI-F704 on the mRNA expression were diminished by cotreatment with the AMPK inhibitor Compound C (Fig. 2A). Subsequent quantitative analysis using RT-qPCR showed the similar influences as RT-PCR. LPS alone significantly increased mRNA levels of IL-6, IL-8, and TNF-α up to 13.5±2.5-fold, 22.6±0.7-fold, and 8.1±0.6-fold of control (p<0.01), respectively. Compared to LPS alone, pretreatment of 40 μM ENERGI-F704 reduced LPS-induced mRNA levels of IL-6, IL-8, and TNF-α to 3.8±0.5-fold, 7.5±1.7-fold, and 3.4±0.2-fold of control, respectively (p<0.05) (Fig. 2B). Besides, the ENERGI-F704-suppressed mRNA levels of IL-6, IL-8, and TNF-α was restored to 10.5±0.6-fold, 17.5±1.4-fold, and 6.9±0.2-fold of negative control, respectively, by cotreatment with Compound C (p<0.05 compare to LPS combining with 40 μM ENERGI-F704) (Fig. 2B).

ENERGI-F704 suppressed mRNA expression and protein production of proinflammatory cytokines, IL-6, IL-8, and TNF-α, in LPS-stimulated RAW264.7 cells. Cells were preincubated with ENERGI-F704 (20 or 40 μM) for 3 h, treated with 0.1 μg/mL LPS for 24 h, and then, the treated cells and resulting supernatants were individually collected for analysis of mRNA expression
In addition to mRNA expression, protein production of the three proinflammatory mediators was also determined. As shown in Figure 2C, LPS alone increased protein production of IL-6, IL-8, and TNF-α to 121.3±11.2, 168.3±13.2, and 507.2±27.3 pg/mL, respectively (p<0.05 compared to control). Treatment with 40 μM ENERGI-F704 combining with LPS lowered protein production of IL-6, IL-8, and TNF-α to 55.6±8.8, 68.5±10.6, and 352.2±20.5 pg/mL, respectively (p<0.05 compared to LPS alone). Similar to the mRNA expression, the ENERGI-F704-suppressed protein production of IL-6, IL-8, and TNF-α was elevated to 82.7±7.2, 111.2±8.2, and 386.9±17.2 pg/mL, respectively, by cotreatment with Compound C (p<0.05 compared to LPS combining with 40 μM ENERGI-F704, Fig. 2C).
ENERGI-F704 inhibited mRNA expression of COX2 and iNOS in RAW264.7 cells and production of PGE2 and NO in response to LPS
Both COX2 and iNOS are known to play important roles in the development and progression of inflammation; therefore, the effects of ENERGI-F704 on mRNA expression of COX2 and iNOS in LPS-stimulated RAW264.7 cells were analyzed. RT-PCR analysis showed that LPS alone significantly elevated mRNA expression of COX2 and iNOS, and the increase of these mRNA expressions was inhibited by pretreatment of ENERGI-F704 in a dose-dependent manner (Fig. 3A). In addition, the inhibitory effects of ENERGI-F704 on COX2 and iNOS were diminished by cotreatment with Compound C. Further quantitative analysis using RT-qPCR revealed that LPS alone significantly increased mRNA levels of COX2 and iNOS to 33.8±2.2-fold and 7.1±0.2-fold of negative control, respectively (p<0.05) (Fig. 3B). Comparing to LPS alone, pretreatment of ENERGI-704 (40 μM) inhibited LPS-induced mRNA levels of COX2 and iNOS to 8.8±0.9-fold and 2.9±0.2-fold of negative control, respectively (p<0.05) (Fig. 3B). Moreover, the ENERGI-F704-suppressed mRNA levels of IL-6, IL-8, and TNF-α was restored to 26.8±1.5-fold and 4.6±0.3-fold of negative control, respectively, by cotreatment with Compound C (p<0.05 compared to ENERGI-F704 combining with LPS) (Fig. 3B). These findings showed that ENERGI-F704 significantly suppressed mRNA expression of COX2 and iNOS in response to LPS, and AMPK activation was involved in the suppression of COX2 and iNOS mRNA expression.
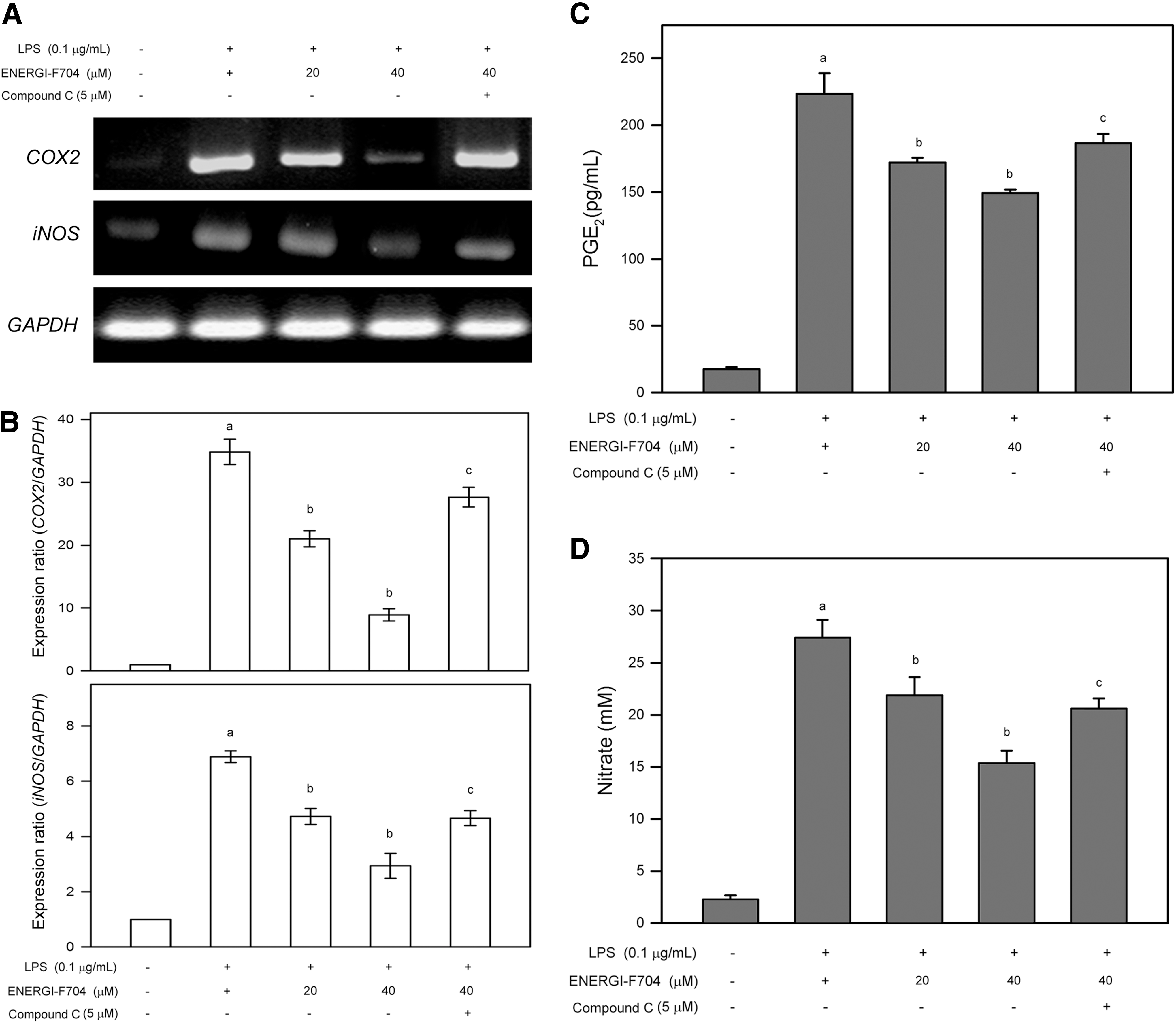
ENERGI-704 inhibited mRNA expression of COX2 and iNOS in LPS-stimulated RAW264.7 cells and reduced the production of PGE2 and NO. Cells were preincubated with ENERGI-F704 (20 or 40 μM) for 3 h, treated with 0.1 μg/mL LPS for 24 h, and then lysed for mRNA extraction. mRNA expression level was analyzed by RT-PCR
In parallel to the analysis of COX2 and iNOS mRNA expression, the effects of ENERGI-F704 on production of PGE2 and NO in response to LPS were also demonstrated. As shown in Figure 3C and D, LPS alone increased PGE2 and NO production to 228.3±16.2 pg/mL and 27.5±1.5 mM, respectively (p<0.05 compared to negative control). Comparing to LPS alone, pretreatment of ENERGI-F704 (40 μM) inhibited LPS-induced production of PGE2 and NO to 154.5±2.1 pg/mL and 15.5±1.1 mM, respectively (p<0.05) (Fig. 3C, D). In addition, the ENERGI-F704-suppressed production of PGE2 and NO was restored to 191.6±8.2 pg/mL and 20.7±0.8 mM, respectively, by cotreatment of Compound C (p<0.05 compared to LPS combining with 40 μM ENERGI-F704) (Fig. 3C, D).
ENERGI-F704 suppressed phosphorylation of PI3K, p38 MAPK, and IκBα and reduced nuclear NF-κB in LPS-stimulated RAW264.7 cells
Activation of PI3K and MAPKs and the consequent NF-κB signaling is reported to involve in LPS-induced production of proinflammatory cytokines (Deng et al., 2013; Huo et al., 2013). Therefore, the effects of ENERGI-F704 on phosphorylation of PI3K, Erk1/2, JNK, p38, and IκBα and nuclear fraction of NF-κB in LPS-stimulated RAW264.7 cells were investigated. As shown in Figure 4A, phosphorylation of PI3K, Erk1/2, JNK, and p38 was significantly enhanced in RAW264.7 cells in response to LPS (p<0.05 compared to negative control). Pretreatment with ENERGI-F704 inhibited LPS-induced phosphorylation of PI3K and p38 in a dose-dependent manner (p<0.05 compared to LPS alone) but insignificantly affected phosphorylation of Erk1/2 and JNK (Fig. 4A, lower panel). In addition, cotreatment with Compound C restored the ENERGI-F704-inhibited phosphorylation of PI3K and p38 and slightly affected the phosphorylation of Erk1/2 and JNK (Fig. 4A, lower panel).
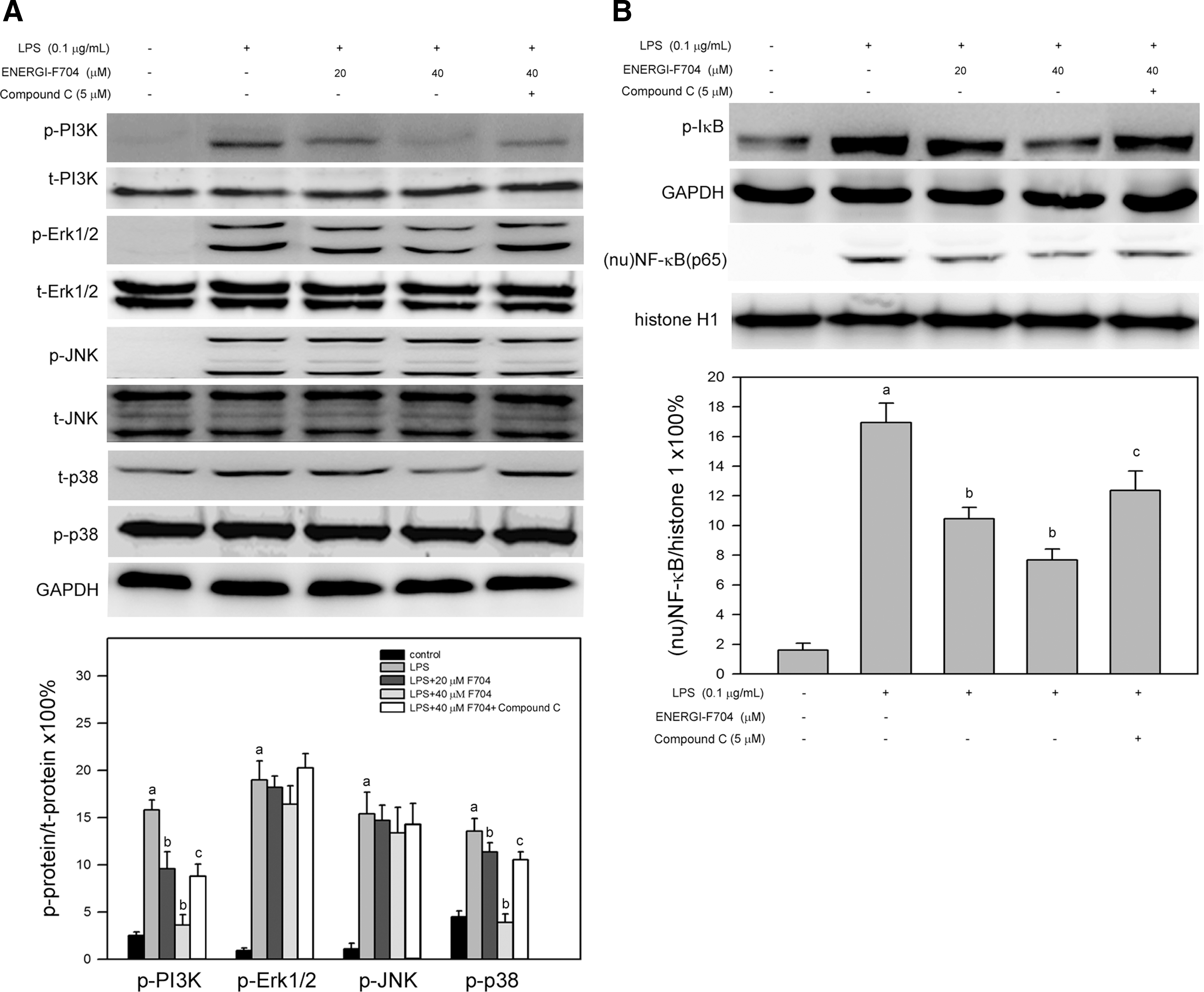
ENERGI-F704 suppressed the phosphorylation of PI3K and p38 and reduced nuclear level of NF-κB in LPS-stimulated RAW264.7 cells. Cells were preincubated with ENERGI-704 (20 or 40 μM) for 3 h, treated with 0.1 μg/mL LPS for 24 h, and then lysed for crude protein extraction and subcellular fractionation. Target proteins were detected using immunoblotting.
In parallel to activation of PI3K and MAPKs, the effects of ENERGI-F704 on NF-κB signaling were also investigated. As shown in Figure 4B, LPS alone significantly increased phosphorylation of IκBα (p-IκBα) and nuclear fraction of NF-κB [(nu)NF-κB(p65)]. Pretreatment of ENERGI-704 reduced both the LPS-induced p-IκBα and (nu)NF-κB(p65). In addition, cotreatment of Compound C restored the ENERGI-F704-reduced p-IκBα and (nu)NF-κB(p65) (Fig. 4B, lower panel, p<0.05 compared to ENERGI-F704 combining with LPS). Taken together, these results showed that ENERGI-F704 suppressed the activation of PI3K, p38, and NF-κB signaling.
Discussion
Inflammatory response induced by tissue injury or infection is highly associated with energy-intensive process. In the presence of LPS, macrophages can immediately switch from a resting state to a highly active state, displaying a great increase in the production of host defense factors, enhanced phagocytosis, and antigen presentation. During the process, ATP generation undergoes a switch from predominant oxidative phosphorylation to glycolysis (Krawczyk et al., 2010). Therefore, it is proposed that alteration of energy metabolism may modulate the inflammatory signaling, interpreting the dual effects of AMPK in macrophage-oriented inflammation (O'Neill and Hardie, 2013). Recent studies have shown that AMPK activator, such as AICAR or resveratrol, exhibits anti-inflammatory activities through AMPK-mediated B-cell lymphoma-6 protein (Bcl-6), poly[ADP-ribose] polymerase 1 (PARP-1), and NF-κB signaling (Yi et al., 2011; Gongol et al., 2013). Similarly, our results showed that an AMPK activator ENERGI-F704 isolated from bamboo activated AMPK by inducing phosphorylation of threonine 172 in RAW264.7 cells, and the ENERGI-F704-induced AMPK activation was involved in suppressing mRNA expression and protein production of IL-6, IL-8, TNF-α, COX2, and iNOS in response to LPS. Moreover, ENERGI-F704 also lowered PGE2 and NO production by RAW264.7 cells exposed to LPS. These results indicate that ENERGI-F704 possesses inhibitory activity on inflammatory responses triggered by murine macrophage with the stimulation of LPS.
In addition to macrophage, neutrophil infiltrating into injury sites is also known as a remarkable inflammatory event in skin wounding (Li et al., 2005). Similarly, our preliminary results also revealed that the external use of ENERGI-F704 exerted anti-inflammatory effects on surgical incision wounding of mice, including reduced neutrophil infiltration (Supplementary Fig. S1 and Table S1; Supplementary Data are available online at
Macrophage is one of the major effector cells during inflammatory process (Murakami et al., 2005). The main function of macrophage is to modulate immune responses through the production of various cytokines, reactive oxygen and nitrogen species, growth factors, and chemokines in response to pathogens or potent immunogens, such as bacterial LPS (Fujiwara and Kobayashi, 2005). Although the bioactive factors produced by macrophages have valuable outcomes in inflammation, these factors were also exhibited unfavorable and damaging effects (Heumann and Roger, 2002). Therefore, the regulation of these products provides a potential target to restrain inflammatory diseases. IL-6 has various biological properties in a number of chronic endothelial dysfunctions, such as modulation of hematopoiesis, proliferation and differentiation of lymphocytes, and induction of acute-phase reactions (Desai et al., 2002). IL-8 is known as a key intermediate regulator in acute inflammatory responses (Noda et al., 2008). TNF-α was a pivotal mediator produced by activated macrophages and was shown to affect various biological processes, including the regulation and the production of other cytokines (Hume et al., 2001; Aggarwal et al., 2002). Our findings revealed that ENERGI-F704 significantly inhibited mRNA expression of IL-6, IL-8, and TNF-α in LPS-stimulated RAW264.7 cells, suggesting that ENERGI-F704 may attenuate the LPS-induced activation of RAW264.7 cells and the consequent inflammatory mediators produced by the murine macrophage.
COX2 and iNOS are important enzymes that catalyze the synthesis of inflammatory mediators in promotion of inflammation. COX is the common target protein for analgesic and anti-inflammatory therapies that have been used for hundreds of years. The COX enzyme consists of at least two isoforms: COX1 and COX2. In mammals, the expression of COX1 is constitutive in normal tissues, such as production of prostaglandin precursors for thromboxane in platelets and the regulation of blood flow. In contrast to COX1, COX2 protein is only slightly expressed in most resting tissues. However, upon inflammatory stimulation, the expression levels of COX2 protein and mRNA are increased (Lee et al., 1992). NOSs are a family of enzymes that catalyze the production of NO from L-arginine. Three isoforms of NOS have been characterized, including neuronal NOS (nNOS), endothelial NOS (eNOS), and inducible NOS (iNOS). nNOS and eNOS mainly exists in neuronal tissues and vascular endothelial cells, respectively, whereas iNOS can be detected in a variety of cell types, including macrophages, microglial cells, astrocytes, and keratinocytes (Andrew and Mayer, 1999). In addition, iNOS is overexpressed in the cells with infectious and proinflammatory stimuli. Our results showed that ENERGI-F704 dose dependently suppressed mRNA expression of COX2 and iNOS, as well as PGE2 and NO production by RAW264.7 cells in response to LPS, indicating that ENERGI-F704 may alleviate adverse reactions attributing to excess production of PGE2 and NO.
LPS is known to activate several signaling kinases, including PI3K, Erk1/2, MAP kinase kinase (MEK) (Geppert et al., 1994), JNK (Hambleton et al., 1996), P38, and other MAP kinase family members (Han et al., 1994), and the consequent NF-κB signaling contributing to expression of proinflammatory mediators. Our findings show that ENERGI-F704 inhibits the activation of PI3K and P38 but not Erk1/2 and JNK, as well as reduced phosphorylation of IkBα and nuclear level of NF-κB, suggesting that the inhibitory effects of ENERGI-F704 on expression of proinflammatory cytokines, COX2, and iNOS may attribute to the modulation of PI3K and P38 and the consequent NF-κB signaling. In conclusion, these findings indicate that ENERGI-F704 possesses potent anti-inflammatory activity on murine macrophage RAW264.7 cells with stimulation of LPS, attributing to the inhibition of PI3K and P38 activation and NF-κB signaling and the consequent suppressed expression of proinflammatory cytokines, COX2, and iNOS.
Footnotes
Acknowledgments
We thank the support of real-time quantitative PCR machine by the Instrument of Center, Chung Shan Medical University, which is partially supported by the Ministry of Education and the Chung Shan Medical University.
Disclosure Statement
The authors have declared that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.