
Abstract
The aim of the study was to investigate the potential mechanisms that interferencing of excision repair cross-complementing gene 1 (ERCC1) mediated by lentiviral vector in cisplatin-resistant ovarian cancer SKOV3/DDP cells. The human platinum-resistant ovarian carcinoma cell line SKOV3/DDP was transfected by pLVX-shRNA lentivirus. Interference efficiency for ERCC1 by lentiviruses transfection was detected by real-time polymerase chain reaction and western blot assay. CCK-8 assay was used for cell proliferation on cell resistance after transfection with ERCC1. Effects of cell apoptosis and cell cycles were detected by flow cytometry. The expression levels of ERCC1 were significantly suppressed in SKOV3/DDP cells after stably transfecting with shERCC1-recombinant plasmid. The results of cell viability assay demonstrated that interference with ERCC1 gene increased the sensitivity of SKOV3/DDP cells to cisplatin (p<0.01). ERCC1 gene-specific silencing promoted cell apoptosis of SKOV3/DDP cells (p<0.01) detected by flow cytometry. Cell cycle analysis results showed that the proportion of cells in G1 and S phase decreased, while the proportion of G2 phase cells increased in ERCC1-silencing cells. The differences were statistically significant (p<0.01), which demonstrated that stable interferencing with ERCC1 induced the cells arrest in G2 phase after being treated by DDP and silencing the expression of ERCC1 inhibited cell proliferation by preventing the progression of cell mitosis. ERCC1 gene silencing effectively reversed SKOV3/DDP cell resistance to cisplatin and increased sensitivity to cisplatin resistance in cisplatin-resistant ovarian cancer cells. Interference of ERCC1 promoted the apoptosis of SKOV3/DDP cells and prevented cell mitosis by inducing G0/G1 phase arrest. Thus, ERCC1 could be a potential therapeutic target for the therapy of cisplatin-resistant ovarian cancer and it would provide new ideas for epigenetic therapy of drug-resistant epithelial ovarian cancer.
Introduction
O
Platinum-based chemotherapy drugs are first-line treatment methods in patients with epithelial ovarian cancer. However, drug resistance was gradually induced in the tumor cells during chemotherapy treatment, which was a major obstacle in clinical therapy (Szakacs et al., 2006). Previous studies had focused on the genetic characteristics in ovarian cancer cells, such as gene mutations, deletions, and other genetic mechanisms. (Quaye et al., 2008; Shah et al., 2010). Recent studies showed that epigenetics played an important role in regulating gene transcription during the occurrence and development of tumors. The drug resistance of ovarian cancer cells was closely related to epigenetic mechanisms (Nephew et al., 2009; Karaca et al., 2013).
For platinum-based therapy, DNA damage and repair has an important role in the resistance of tumor cells to platinum complexes (Altaha et al., 2004). The DNA repair capacity may be closely related to the progression of cancer in patients. Some research showed that the intracellular platinum -DNA complexes can be repaired through nucleotide (nucleotide excision repair [NER]) pathway (Koc et al., 2012). Excision repair cross-complementation group 1 (ERCC1) plays an important role in the NER pathway, which is considered as a key gene in NER pathway. It is associated with resistance of platinum-based chemotherapy in different types of tumors (Ceppi et al., 2008; Ozkan et al., 2010; Fisher et al., 2013). Platinum resistance constitutes one of the most recognized clinical challenges for ovarian cancer. The expression of ERCC1 directly affects the physiological DNA repair process (Gregg et al., 2011). High expression level of ERCC1 is an important key leading to platinum drugs resistance in tumor cells. Some reports have demonstrated higher expression of ERCC1 in mRNA and protein levels in human malignancies were related to cisplatin-based chemotherapy sensitivity and directly determined the survival rates of patients (Liu et al., 2006; Ma et al., 2014; Muallem et al., 2014). However, the mechanism of ERCC1 on drug resistance in ovarian cancer is still not clearly clarified. In this study, we constructed lentiviral vectors targeting ERCC1 gene and successfully inhibited the expression of ERCC1 in ovarian cancer cells by a lentivirus-mediated RNA interference (RNAi) and explored the effects of ERCC1 on platinum-resistance in epithelial ovarian cancer.
Materials and Methods
Cell lines and plasmid
Cisplatin-resistant epithelial ovarian cancer cell line SKOV3/DDP and cisplatin-sensitive cell line DKOV3 were obtained from the Cancer Institute and Hospital, Chinese Academy of Medical Sciences (Beijing, China). The pLVX-shRNA vector was purchased from Clontech Corporation.
Agent and instrument
Revert Aid™ First Strand cDNA Synthesis Kit was used for reverse transcription and was obtained from Thermo Fisher Scientific Corporation. Total RNA extraction reagent TRIzol was obtained from Invitrogen. Fluorescence quantitative polymerase chain reaction (PCR) kit was FastStart Universal SYBR Green Master and was obtained from Roche Applied Science Corporation. The fluorescence quantitative PCR instrument ABI 7500 was obtained from ABI Applied Biosystems, Inc. Multifunctional microplate reader was obtained from BioTEK. Restriction endonucleases and T4 DNA ligase was obtained from TAKARA. Gel extraction kit was purchased from Qiagen Corporation. Electrochemiluminescence color kit and bicinchoninic acid (BCA) protein assay kit were purchased from Biological Technology Development Co., Ltd. Nanjing KGI; DDP was purchased from Qilu Pharmaceutical Co., Ltd.
Cell culture and transfection
SKOV3 cells were routinely cultured in RPMl640 medium containing 10% fetal bovine serum and placed in the incubator at 37°C with the concentration of 5% CO2. SKOV3/DDP cell culture conditions were consistent with that of SKOV3 cells, but substituting for every two generation. DDP was added to the culture medium at a final concentration of 5 μM to maintain DDP resistance.
The pLVX-shRNA was an empty vector, called pLVX-ERCC1shRNAC, and the recombinant vectors were called pLVX-ERCC1shRNA1, pLVX-ERCC1shRNA2, and pLVX-ERCC1shRNA3, respectively. The cells were transfected with lenti-X™ lentiviral expression systems kit for lentiviral packaging according to the protocols. Generally, ∼3×105 SKOV3/DDP cells were plated in 25 cm2 cell culture flasks. After 18 h, polybrene was added at the final concentration of 8 μg/mL and 100 μL of virus supernatant was added. The cells were cultured for 24 h, followed by removal of RPMI-1640 medium containing lentiviruses, and normal RPMI-1640 complete medium was added for 24 h. Then, 2 μg/mL of puromycin was used for screening every 2 days. The total time for screening was 7 days. After the screening course, the cell lines were gotten containing the plasmids of shRNAC, pLVX-ERCC1shRNA1, pLVX-ERCC1shRNA2, and pLVX-ERCC1shRNA3, respectively.
Reverse transcription-polymerase chain reaction is used to detect the mRNA levels of ERCC1
Total RNA was extracted using TRIzol reagent, and the integrity was determined by 1.5% agarose gel electrophoresis. Nanodrop UV spectrophotometer was used to measure RNA concentration and purity. The total RNA sample was used as a template and the complementary strand was synthesized by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) cDNA kit. Real-time PCR reactions were performed on the ABI 7500 instrument. Primers were synthesized by Shanghai Invitrogen Biotech Co. Ltd. The sequences of the primers were as follows: ERCC1 up: GGGGCAAAATCCAACAGCATC and ERCC1 down: AGTCGGGAATTACGTCGCCAA; glyceraldehyde 3-phophate dehydrogenase (GAPDH) up: AGCCACATCGCTCAGACAC and GAPDH down: CTCGCTCCTGGAAGATGGT. The PCR reaction conditions were 95°C, 10 min, followed by 40 cycles of 95°C, l5 s, and 59°C, 40 s. Relative quantitative analysis is performed using 2−Δ CT method and GAPDH is used as reference gene for correction. The experiment was repeated three times.
Expression of ERCC1 is detected by western blotting analysis
Whole cell extracts were prepared and the total proteins were extracted and quantified by BCA method. The total proteins were added loading buffer solution containing sodium dodecyl sulfate (SDS) and boiled 5 min for denaturation. The proteins were separated by polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride (PVDF) membrane. The membrane was blocked at 37°C for 40 min with 5% skim milk powder. The primary antibodies included rabbit anti-human ERCC1 polyclonal antibody and rabbit anti-human GAPDH polyclonal antibody (all diluted 1: 1000) used overnight at 4°C.
Then, the membrane was incubated with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (Santa Cruz) for 1 h at room temperature. PVDF membrane was exposed at darkroom using enhanced chemiluminescence developing X-ray film with an ECL detection kit (Amersham). The experiment was repeated three times.
Cell viability detected by CCK-8
Cell viability was determined by CCK-8. Generally, 2×104 cells/well were plated into 96-well plates. After 24 h, the cells were treated with DDP for 24 h at concentrations of 3, 6, 12, and 20 μg/mL with five replicates. Before the test, 10 μL of CCK-8 agent was added into the wells, and cultured for 2 h at 37°C. The OD values were read on a microplate reader at a test wavelength of 450 nm and cell viability was calculated. Cell viability=(OD450 in treatment group −OD450 in blank wells)/(OD450 in untreatment group −OD450 in black wells)×100%. Data were expressed as mean±standard deviation.
Cell apoptosis is detected by annexin V-PI dual staining analysis
The cell apoptosis was tested by annexin V-PI staining and the method was according to the kit protocols (Santacruz). Generally, the cells were plated into six-well plates. After 24 h, the cells were treated with DDP at the concentration of 6 μg/mL for 24 h. Then, the cells were digested with 0.25% trypsin for 1–2 min and washed twice with precooled phosphate-buffered saline (PBS). The cells were resuspended in binding buffer (HEPES–NaOH 10 mM pH 7.4, 25 mM CaCl2, and 144 mM NaCl). The staining dye of EGFP-annexin V (0.1 μg/μL) and PI (0.05 μg/μL) was added and incubated in dark for 15 min at room temperature. Finally, cells were then subjected to fluorescence activated cell sorting (FACS) analysis for detection within 1 h. PI was detected in FL1 channel, and EGFP was detected in FL2 channel. Each sample collected 10,000 cells for detection.
Cell cycle analysis
The cells in the logarithmic growth phase were plated into six-well plates, and treated with 6 μg/mL DDP for 24 h. The cells were digested and adjusted at the density of 1×106/mL. The cells were centrifuged at 2000 rpm for 5 min, and the supernatant was discarded. The cells were washed twice with precooled PBS. Then, the cells were fixed and resuspended by 5 mL of 70% ethanol at 4°C overnight. They were washed twice and Rnase A was added with final concentration of 200 μg/mL at 37°C for 30 min. Then, the staining dye of PI (0.05 μg/μL) was added with the final concentration of 20 μg/mL and incubated in dark for 30 min on ice. Finally, cells were subjected to FACS analysis and 10,000 cells were counted per sample.
Statistical analysis
SPSS statistical package (13.0) was used to analyze the data. A two sample t-test was used to compare mean for two groups of cases. Mean value between multiple groups were compared using analysis of variance (ANOVA). LSD-t test was used for pairwise comparisons. The results of cell viability was analyzed by two-way ANOVA. All data were shown as mean±standard error of the mean (x±s). p-Values<0.05 were defined as statistical difference.
Results
The most effective shRNA vector is screened for interfering ERCC1 gene
To detect the effectiveness of shRNA vectors for interfering ERCC1 gene, three pairs of interfering vectors were designed for specific interferencing with endogenous ERCC1 gene. The expression of ERCC1 was detected by RT-PCR and GAPDH was used as positive control in this study. As shown in Figure 1A, the relative expression level was 0.25, 0.84, 0.41, and 1.00, respectively, when the cells were infected with the vectors of PLVX-ERCC1-shRNA1, PLVX-ERCC1-shRNA2, PLVX-ERCC1-shRNA3, and PLVX-ERCC1-shRNAC. The result was consistent with that from western blotting analysis (Fig. 1B). This suggested that PLVX-ERCC1-shRNA1 had the best interferencing effect and it was the best effective vector for interferencing ERCC1 gene.
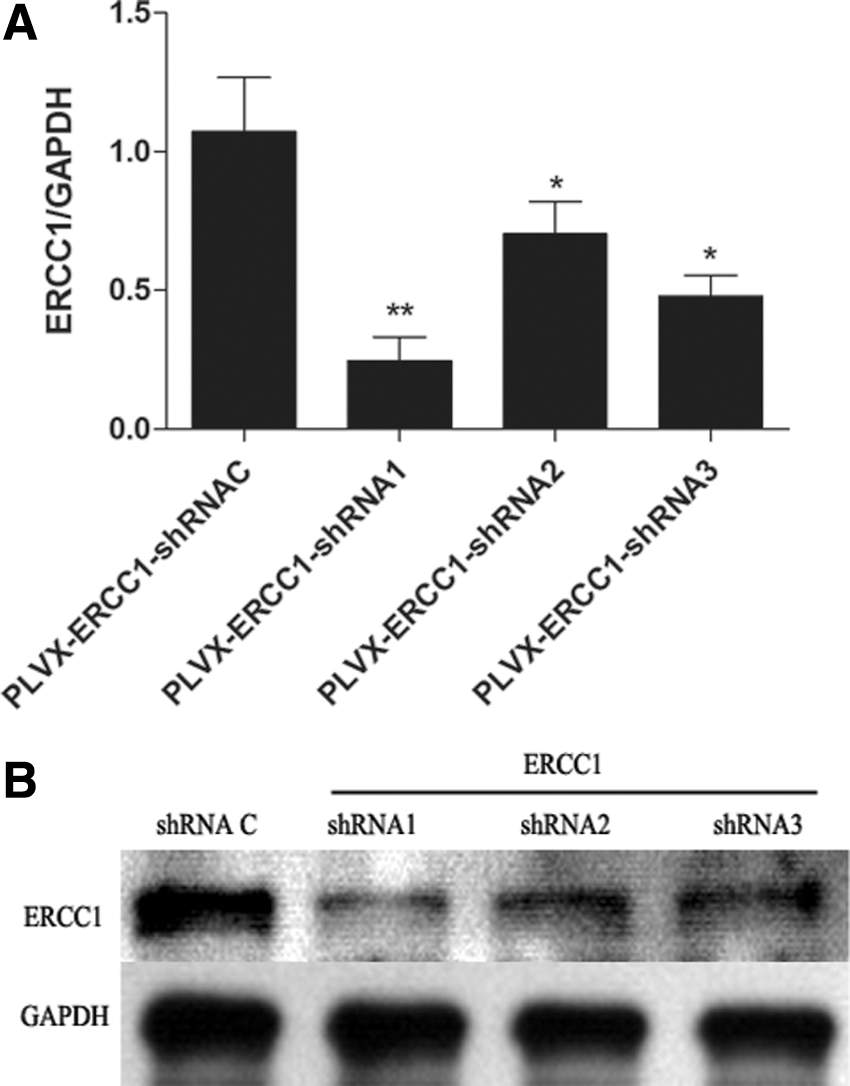
The most effective shRNA vector is screened.
Identification of cell lines with silencing ERCC1 gene
The cells infected with PLVX-ERCC1-shRNA1 vector and control vector were screened by puromycin, and a pair of stably transfected cell lines were obtained. As seen in Figure 2, the western blotting analysis demonstrated that the expression level of ERCC1 in SKOV3/DDP-shRNA1 cells was obviously lower than that in SKOV3/DDP cells infected by shRNA control vectors. All of the data suggested that the stable cell line with ERCC1 silencing was successfully constructed.
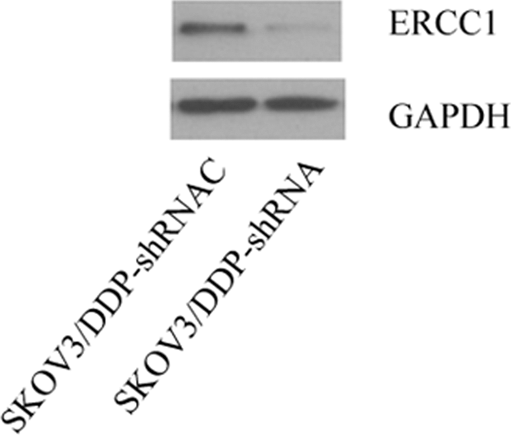
The expression levels of ERCC1 in SKOV3/DDP cells transfected with ERCC1-specific shRNA and shRNA control vector. Stable cell lines with ERCC1 silencing is screened by western blotting analysis. The cells were plated in 24-well plates. Eight hours later, the cells were infected with PLVX-ERCC1-shRNA1 vector and control vector. Then, the transfected cells were screened by puromycin and a pair of stably transfected cell lines were obtained.
The effects of ERCC1-specific shRNA on DDP resistance in SKOV3/DDP cells
The stable cell line was treated with different doses of DDP for 24 h. As shown in Figure 3, the results of cell viability assay demonstrated that cell viability in stable cells transfected with specific ERCC1 shRNA was significantly lower than the control cells, and they were statistically different (p<0.05). These results suggested that ERCC1 gene-specific silencing could enhance the sensitivity of SKOV3/DDP cells to DDP.
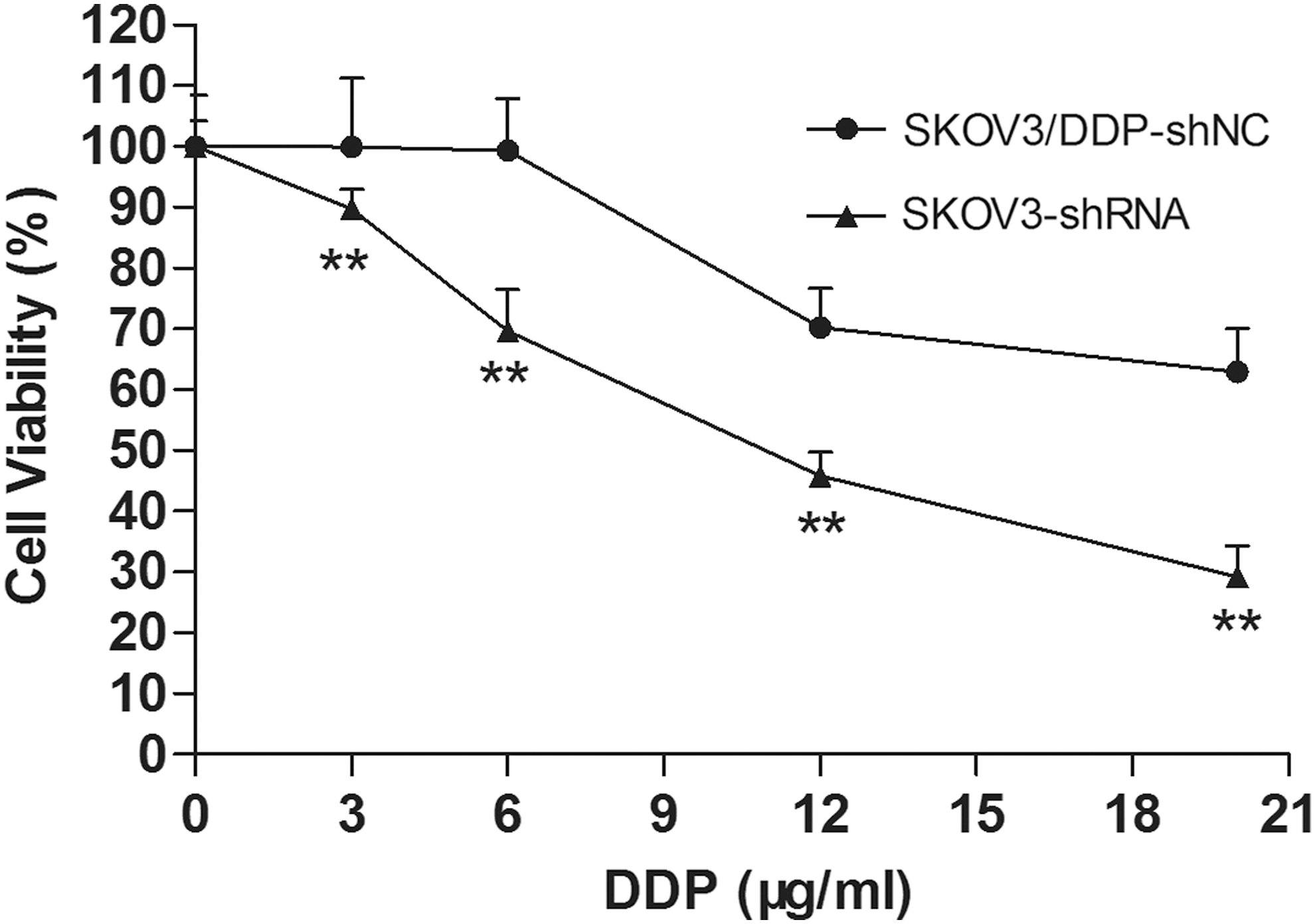
The effects of ERCC1-specific shRNA on DDP resistance in SKOV3/DDP cells. The stable cell lines were treated with different doses of DDP for 24 h, and cell viability assay was performed. **p<0.01.
Interferencing ERCC1 with shRNA promotes apoptosis of SKOV3/DDP
To investigate the mechanism that how the drug DDP affects the cell viability of SKOV3/DDP with ERCC1 interference, we tested the apoptosis rates in DDP-treated stable cell lines transfected with ERCC1 shRNA and negative controls. As shown in Figure 4, the results demonstrated that the stable cell lines with ERCC1-specific interference had significantly lower tolerance than that of control cells. The apoptosis rate in ERCC1 stable interferencing cell lines was 17.8%, which was much higher than that of 10.6% in control cell lines, and there was statistical difference between them (p<0.05).
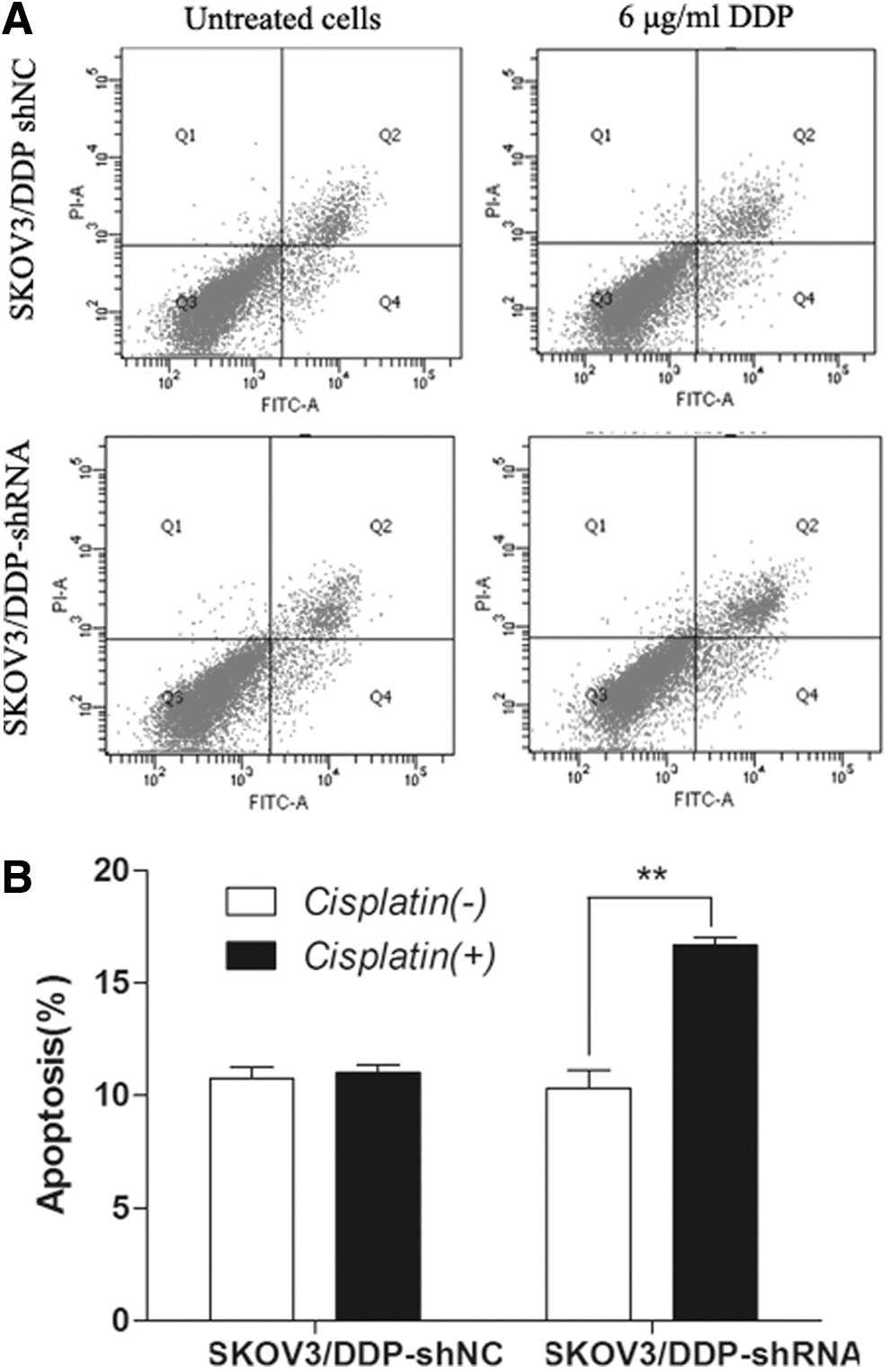
Interfering ERCC1 with shRNA promotes apoptosis of SKOV3/DDP.
The effects of ERCC1 silencing on cell cycle in SKOV3/DDP cells
Next, cell cycle analysis of the cells was analyzed by FACS assay. As shown in Figure 5, the distribution of the cells showed that compared with control cells, the rates of the stable cells with ERCC1 interferencing in G1 and S phase were decreased, while the rate of G2 phase was increased. There was statistical difference (p<0.05). All the results suggested that G2 phase arresting of the cells with ERCC1 interferencing was induced after treatment with DDP. Thus, ERCC1 silencing could inhibit cell proliferation by preventing cell mitosis.
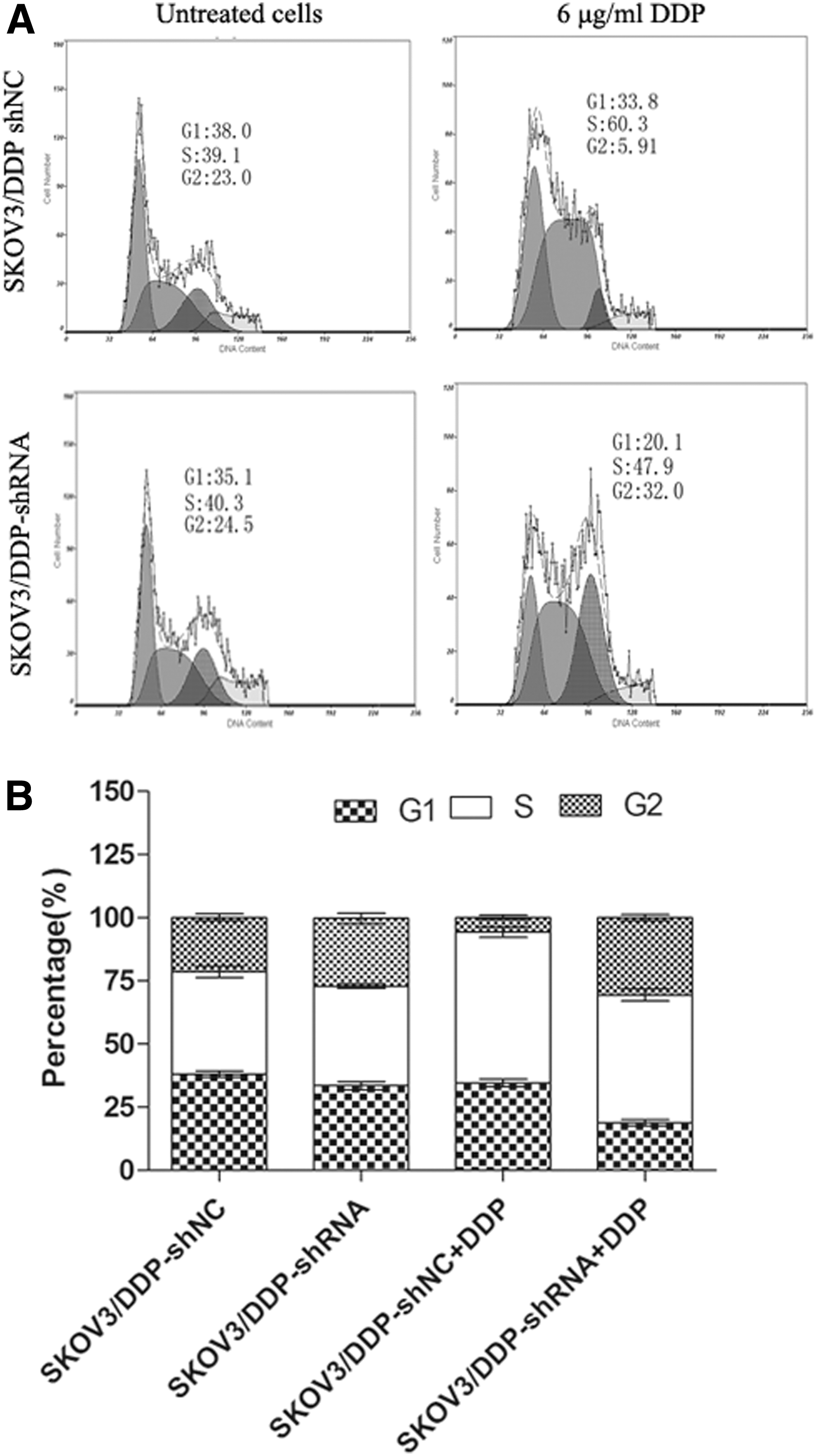
The effects of ERCC1 silencing on cell cycle in SKOV3/DDP cells.
Discussion
According to recent estimates and statistics from the American Cancer Society, there were about 490,000 cases of deaths in cancer patients every year, more than 90% of which were caused by tumor resistance (Wang and Chen, 2010; Yin et al., 2013). Therefore, drug resistance is a realistic and pressing problem in today's cancer treatment. Ovarian cancer is the fifth most common cancer among women, and the mortality rate ranks first in gynecological malignancies. Chemotherapy resistance is an important obstacle in improving survival rates in patients with ovarian cancer (Yin et al., 2013). For instance, 20–30% of epithelial ovarian cancer patients had primary platinum-resistant tumors, and secondary resistance eventually developed in more than 80% of patients. Majority of patients will relapse and eventually die of drug-resistant disease.
Compared with low levels of ERCC1, high ERCC1 levels were associated with shorter survival and poor prognosis in patients with nonsmall-cell carcinoma, which was not related to clinical therapy (Milovic-Kovacevic et al., 2011; Murphy and Stordal, 2011; Rubatt et al., 2012). High expression level of ERCC1 is one of the predictive indicators for poor effects of platinum-based chemotherapy (Li et al., 2014). It was reported that ovarian cancers with negative ERCC1 expression have a better response to platinum-based chemotherapy than those with positive ERCC1 expression (Yan et al., 2012), and ERCC1 protein expression status is significantly associated with response to platinum-based chemotherapy in ovarian cancers (Li et al., 2013). To study the potential role of ERCC1 on drug resistance in ovarian cancer cells, we used real-time PCR and immunoblot assay to detect ERCC1 expression in cisplatin-resistant and cisplatin-sensitive ovarian cancer cells. Then, RNAi technology was used to test cell resistance variation after silencing ERCC1 gene in ovarian cancer cells SKOV3/DDP. The results showed that ERCC1 mRNA levels were significantly increased in DDP-resistant cell line SKOV3/DDP than that of DDP-sensitive cells SKOV3, suggesting that overexpression of ERCC1 gene was associated with drug resistance to DDP. To further explore the relationship between ERCC1 gene expression and drug resistance in epithelial ovarian cancer chemotherapy, recombinant lentiviral vector targeting ERCC1 gene silencing was constructed in this study. Next, it was used for transfection of DDP-resistant epithelial ovarian cancer cell line SKOV3/DDP, and reversal of drug resistance on DDP was studied by silencing the expression of ERCC1 gene. The results from CCK-8 assay showed that stable transfection of shRNA specific to ERCC1 significantly increased sensitivity to DDP in SKOV3/DDP cells and reversed cisplatin resistance in SKOV3/DDP cells to some extent, suggesting that ERCC1 played an important role on DDP resistance in ovarian cancer cells.
To detect the effects of ERCC1 silencing on drug resistance and apoptosis in ovarian cancer cells, we used SKPV3/DDP as cell model and interferred the expression of ERCC1 by a lentiviral RNAi system targeting ERCC1. The results revealed that the cell viability of SKOV3/DDP cells transfected with ERCC1 shRNA was significantly lower compared with cells transfected with negative control shRNA, which demonstrated that specific interferencing of the expression of ERCC1 could enhance the sensitivity of SKOV3/DDP cells to DDP. Moreover, the apoptosis rates were significantly increased in ovarian cells transfected with ERCC1-shRNA, which showed that ERCC1 silencing had a close relationship with cell apoptosis. ERCC1 interference was involved in the cisplatin-resistance in ovarian cancer cells in which cell apoptosis might be the major reason.
To further explore the possible role of ERCC1 on drug resistance in ovarian cancer cells, we detected the cell cycles in ERCC1 interferencing SKVO3/DDP cells. Consistently, interference of ERCC1 blocked proliferation of DDP-resistant cancer cells by inhibiting G1 and S phase and inducing G2 cell-cycle arrest. All of the data demonstrated that ERCC1 silencing inhibited cell proliferation by preventing cell mitosis and ERCC1 may play a role in ovarian cancer cell proliferation by regulating G2 phase during cell cycles.
In conclusion, high expression of ERCC1 is closely related to the proliferation of epithelial ovarian cancer cells resistant to cisplatin. RNAi technology targeting ERCC1 gene can effectively restore the sensitivity to chemotherapy in SKOV3/DDP cells by promoting cell apoptosis. The results in this study confirmed the idea that ERCC1 silencing could provide a useful therapeutic protocol for the treatment of cisplatin-resistant ovarian cancer. It will offer new targets for clinical therapy of ovarian cancer.
Footnotes
Disclosure Statement
No competing financial interests exist.