
Abstract
The mTOR/S6K1 signaling pathway is the primary regulator of milk protein synthesis. While mTOR is known to be regulated at the translational level by amino acids, the mechanism by which mTOR accepts the amino acid signal is not yet clear. In this study, we describe the discovery of WISP3 as a potentially novel signaling factor that connects mTOR and amino acids. Treatment of dairy cow mammary epithelial cells with amino acids (lysine or methionine) increased both cell growth and the expression of β-casein (CSN2), WISP3, mTOR, and phospho-mTOR (p-mTOR). Notably, overexpressing WISP3 in these cells also increased both cell growth and the expression of CSN2, mTOR, and p-mTOR and decreased the expression of glycogen synthase kinase 3β (GSK3β), while repressing WISP3 had the opposite effect. The increase of the expression of CSN2, mTOR, and p-mTOR mediated by amino acid could be inhibited by repressing WISP3. The increase of the expression of CSN2, mTOR, and p-mTOR mediated by WISP3 overexpression could be inhibited by overexpressing GSK3β, and vice versa. Taken together, these results reveal that through its amino acid-mediated regulation of the mTOR pathway, WISP3 is an important regulatory factor involved in the amino acid-mediated regulation of milk protein synthesis and cell growth.
Introduction
W
The main signaling pathways of milk protein expression in mammary glands of mammals are the Jak2-Stat5 and mTOR/S6K1 signaling pathways (Yang et al., 2000a). Currently, the transcriptional process of milk protein synthesis is known to be regulated by prolactin through the JAK/STAT5 signaling pathway just as its translational process is regulated by amino acids through the mTOR/S6K1 signaling pathway (Yang et al., 2000b; Galbaugh et al., 2006; Jankiewicz et al., 2006; Wang and Proud, 2006; Prizant and Barash, 2008; Toerien et al., 2010). The activity of mTOR also appears to be modulated by branched-chain amino acids (AA) (Kimball and Jefferson, 2006; Moshel et al., 2006), and the availability of amino acids is the main limiting factor for milk protein synthesis (Reynolds et al., 1994; Appuhamy et al., 2011a), and according to previous reports, lysine and methionine were the most limiting in bovine mammary glands (Bequette et al., 1998; Hanigan et al., 2002). In addition, many previous reports have shown that lysine, as the most limiting, could modulate mammary protein synthesis (Vyas and Erdman, 2009; Appuhamy et al., 2011b; Lee et al., 2012; Wang et al., 2012; Nan et al., 2014; Paz and Kononoff, 2014). However, the precise mechanism by which mTOR complex 1 (mTORC1) activation is mediated by intracellular amino acids is poorly understood. While several studies have investigated this mechanism in a variety of formats (Bonfils et al., 2012; Durán and Hall, 2012; Han et al., 2012), it has not yet been studied in the regulation of dairy cow mammary lactation.
Cytokeratins are intermediate filaments of epithelial cells and important protein markers to identify epithelial cells (Bartek et al., 1985; Danowski et al., 2012; Sorg et al., 2012). As a member of the intermediate filament gene family, cytokeratin 18 (CK18) is used to determine epithelial cell property (Schmid et al., 1983; Hu et al., 2009; Su et al., 2013). Milk protein secretion is an important mammary-specific feature, and β-casein (CNS2) is one of the major components of milk protein (Huynh et al., 1991). The expression of CSN2 can be used to test whether the cells cultured in vitro exhibit normal mammary function (Strange et al., 1991; Ahn et al., 1995; Anand et al., 2012; Fu et al., 2014).
In this study, we investigate the function of WISP3 in dairy cow mammary epithelial cells (DCMECs) and show that WISP3 plays an important role in CSN2 synthesis and cell growth by sensing the intracellular amino acid (lysine [Lys] and methionine [Met]) and regulating amino acid-induced mTORC1 activation. Moreover, in amino acid-treated positive control cells, amino acid signaling was reliant, at least in part, on WISP to induce CSN2 synthesis and cell growth through its regulation of the mTOR pathway. As for the mechanism by which WISP3 regulates mTOR signaling in these cells, the overexpression of WISP3 inhibits glycogen synthase kinase 3β (GSK3β), an mTOR repressor, in cells expressing GSK3β.
Materials and Methods
Cell preparation and treatments
DCMECs were cultured according to a previous report by Tong et al. (2011). The cells were cultured in DMEM/F12 medium containing 10% fetal bovine serum. Establishment of the lactation model regulated by Lys was completed according to a previous report by Lu et al. (2013). The treatment of cells with Met was completed according to a previous report by Lu et al. (2012).
Immunofluorescence
DCMECs were plated onto glass coverslips at 1×105 cells per mL in culture dishes. When the cells grew to 80% confluence, the media were removed and the cells were rinsed twice with PBS and fixed in 4% (w/v) ice-cold formaldehyde at 4°C for 10 min. Cells were then rinsed thrice with tris-buffered saline with 5% skim milk and 0.1% Tween-20 (TBST) for 5 min. Following blocking in blocking buffer (TBS with 5% BSA and 0.1% Triton X-100) for 1 h at 37°C, cells were incubated with anti-CK18 primary antibody (Santa Cruz Biotechnology, Inc.) and anti-β-casein primary antibody (Abbiotec) at a 1:200 dilution for 1.5 h at 37°C, respectively. After washing thrice in TBST, specimens were incubated in the dark with FITC-conjugated secondary antibodies at a 1:200 dilution for 1 h at 37°C, and then washed thrice with TBST. One μg per mL of propidium iodide (PI) was used for nuclear staining at 37°C for 10 min. Cells were then rinsed thrice with TBST and affixed on the slides (antifluorescent quencher liquid was dropped on the slides), and then observed by laser scanning confocal microscopy (LEICA) (Chan et al., 2010).
Construction of WISP3 eukaryotic expression plasmid and transfection
Total RNA from cultured DCMECs was extracted using Trizol reagent (Invitrogen), and the cDNA was synthesized using M-MLV reverse transcriptase (Promega). The CDS area of the WISP3 gene (NCBI Reference Sequence: XM_003586465.2) was amplified using polymerase chain reaction (PCR) and specific primers (Forward: 5′-GGA
Cells were transfected with pGW using the Lipofectamine™ 2000 transfection reagent (LF2000; Invitrogen) according to the manufacturer's instructions. Cells were incubated at 37°C in 5% CO2 for 24 h, after which they were collected for further experiments. pGCMV-IRES-EGFP (empty vector control, EV) and untransfected (blank, B) cells were incubated with the same conditions as the control (Zhang et al., 2014). For the cotransfection with WISP3-specific small interfering RNA (siRNA) and WISP3 overexpression vector, the dose of WISP3-specific siRNA was 1.0 μg per well of six wells and the dose of WISP3 overexpression vector was 3.5 μg per well of six wells according to the manufacturer's instructions of Lipofectamine 2000.
Small interfering RNA transfection
WISP3-specific siRNA, GSK3β-specific siRNA, and negative control siRNA were synthesized by GenePharma Co., Ltd. DCMECs were transfected with either WISP3-specific siRNA (WI) or negative control siRNA (NC) using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. Transfected cells were cultivated for 24 h, and then the cells were collected for further experiments (Wang et al., 2014).
Quantitative real-time PCR
Total RNA extraction from DCMECs and the quantitative real-time PCR (qRT-PCR) analysis method were used according to a method previously reported by Huang et al. (2012). Subsequently, the total RNA was reverse transcribed into cDNA using thermoscript reverse transcriptase (TaKaRa) according to the manufacturer's instructions. qRT-PCR reactions were performed using the Sensimix™ SYBR and Fluorescein real-time PCR kit, and the analysis was performed using an ABI PRISM 7300 RT-PCR System (Applied Biosystems) with a total volume of 20 μL per well in 96-well microwell plates. All target mRNAs were normalized to the level of β-actin mRNA in each sample. The gene transcript primers are shown in Table 1.
Data analysis was performed with the comparative Ct method using SDS V1.2 software.
Western blotting analysis
Western blotting (WB) analysis was performed using standard techniques reported by Luo et al. (2013). After 24 h of transfection, each group of cells was harvested with lysis buffer (20 mM Tris–HCl, pH 7.5; 150 mM NaCl; 2.5 mM sodium pyrophosphate; 1 mM sodium β-glycerophosphate; 5 mM NaF; 1 mM Na3VO4; 1 mM phenylmethylsulfonyl fluoride; 1% Triton X-100; and 1 tablet of protease inhibitor mixture per 40 mL of lysis buffer). Cellular debris was removed by centrifugation (12,000 g for 10 min at 4°C). Cell lysate (containing about 30 μg of protein) was separated on a 10% SDS-PAGE gel and transferred onto nitrocellulose membranes (Bio-Rad) using 20 V of constant voltage. Following blocking in TBST for 1.5 h at 37°C, membranes were probed with primary antibodies specific for the following antibodies: WISP-3, STAT5a, phospho-Stat5a (p-STAT5a), β-actin (Santa Cruz), mTOR, phospho-mTOR (p-mTOR) (Cell Signaling Technology), and β-casein (Abbiotec) for 1.5 h at 37°C, washed several times with 5% milk/TBS, and incubated with HRP-conjugated secondary antibodies (1:1000) (ZSGB-BIO). The chemiluminescence detection of HRP-conjugated secondary antibodies was performed using Super ECL Plus (ApplyGEN). Antibodies purchased from Santa Cruz and Abbiotec were used at 1:500 dilutions, and those purchased from Cell Signaling Technology were used at 1:1000 dilutions. Analysis of the WB results was subsequently performed using the Quantity One software.
Cell viability assay
Cell viability was determined using a CASY model TT Analyzer System (Schärfe System GmbH) according to the manufacturer's instructions. The experimental operation used was according to a method previously reported by Li et al. (2012).
Statistical analysis
The results are reported as mean±SE. Data statistics and individual differences among groups were analyzed using the t-test by Sigma Plot 9.0 software, and differences were considered statistically significant at p<0.05 or p<0.01. Gray-scale scanning of WB results was analyzed by BandScan 4.3 software. All data were obtained from at least three independent experiments.
Results
Lys and Met promoted both cell growth and the expression of CSN2 and WISP3
Primary DCMECs were cultured and purified according to a previous report by Wang et al. (2014). The expression of CK18 and CSN2 in the purified DCMECs was tested by immunofluorescence (IF), using mouse IgG as a control. The results are shown in Figure 1A. The purified DCMECs showed positive staining for CK18 and CSN2, suggesting that these cells were indeed responsible for the secretion of CSN2.
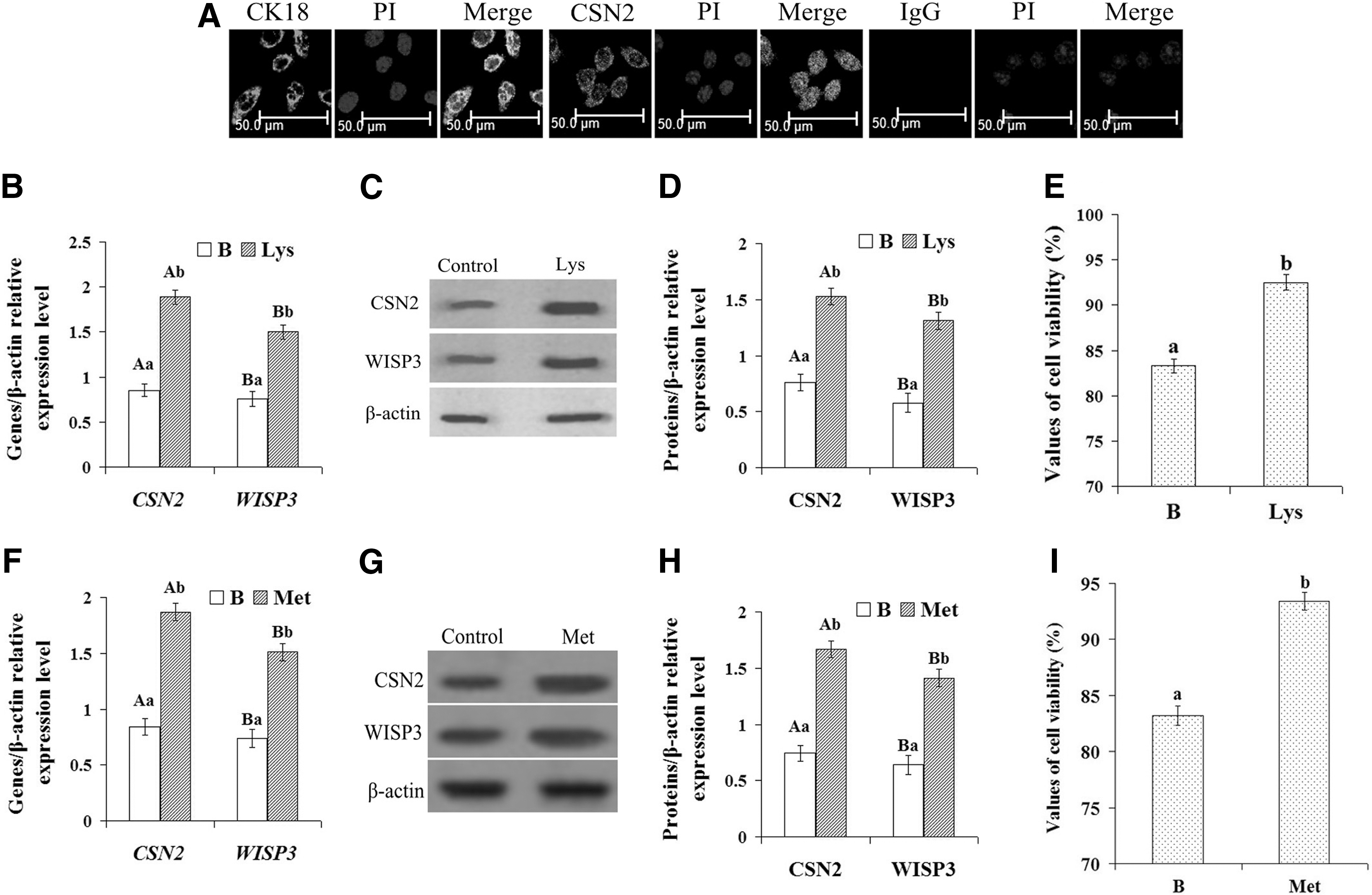
Culture and identification of DCMECs and expression of CSN2 and WISP3 in DCMECs and viability of DCMECs treated with amino acid.
To establish a model of DCMEC lactation regulated by Lys, the purified cells were treated with 1.2 mM Lys for 24 h according to a previous report by Lu et al. (2013), while normal cultured cells were used as a control. The expression of CSN2 and WISP3 in the Lys-treated and control groups was tested by qRT-PCR and WB. The results are shown in Figure 1B–D. Compared with the control, the expression of CSN2 and WISP3 genes was significantly increased (Fig. 1B, p<0.01), as was the synthesis of CSN2 and WISP3 proteins (Fig. 1C, D, p<0.01). After treatment with 1.2 mM Lys for 24 h, cell viability was determined using a CASY model TT cell counter. The results showed that in comparison with the control group, cell viability was significantly increased (Fig. 1E, p<0.01). Taken together, these results showed that WISP3 and CSN2 were both upregulated following Lys stimulation, suggesting that WISP3 may be a regulatory factor involved in the Lys-mediated regulation of CSN2 synthesis and cell growth.
To detect whether other amino acids also had the similar affect on cells, cells were treated with 0.6 mM Met for 24 h according to a previous report by Lu et al. (2012), and the expression of CSN2 and WISP3 and the cell viability were tested. The results are shown in Figure 1F–H. The results were similar to treatment with Lys; when treated with Met, the expression of CSN2 and WISP3 and the cell viability were significantly increased (Fig. 1F–H, p<0.01). To detect whether the effect of Lys on the cells is dose-dependent, cells were treated with different concentrations of Lys. The results showed that the effect of Lysine on the cells is dose-dependent and 1.2 mM was the optimal concentration (Supplementary Fig. S1; Supplementary materials are available online at
WISP3 overexpression promoted both cell growth and the expression of milk protein synthesis-associated proteins
Purified cells were divided into three groups: the untransfected group (B), the transfected empty plasmid group (EV), and the WISP3-overexpressing group (WO). The expression of mTOR, Stat5a, CSN2, and WISP3 mRNA transcripts was tested using qRT-PCR. The results showed that the expression of mTOR, CSN2, and WISP3 was significantly increased in the WO group compared with that in the B and EV groups (Fig. 2A, p<0.01); however, Stat5a expression was unchanged (Fig. 2A, p>0.05). The expression of mTOR, p-mTOR, Stat5a, p-Stat5a, CSN2, and WISP3 proteins was tested by WB. The WB results were consistent with the qRT-PCR results, showing that the expression of mTOR, p-mTOR, CSN2, and WISP3 was also significantly increased in the WO group compared with that in the B and EV groups (Fig. 2B, p<0.01); however, Stat5a and p-Stat5a were unchanged (Fig. 2B, p>0.05). Cell viability was determined using a CASY model TT cell counter. The results showed that cell viability was significantly increased in the WO group compared with that in the B and EV groups (Fig. 2D, p<0.01). These data indicate that WISP3 may upregulate CSN2 synthesis and cell viability in DCMECs, and this function may be related to the mTOR signaling pathway, but not the Jak2/Stat5a signaling pathway.
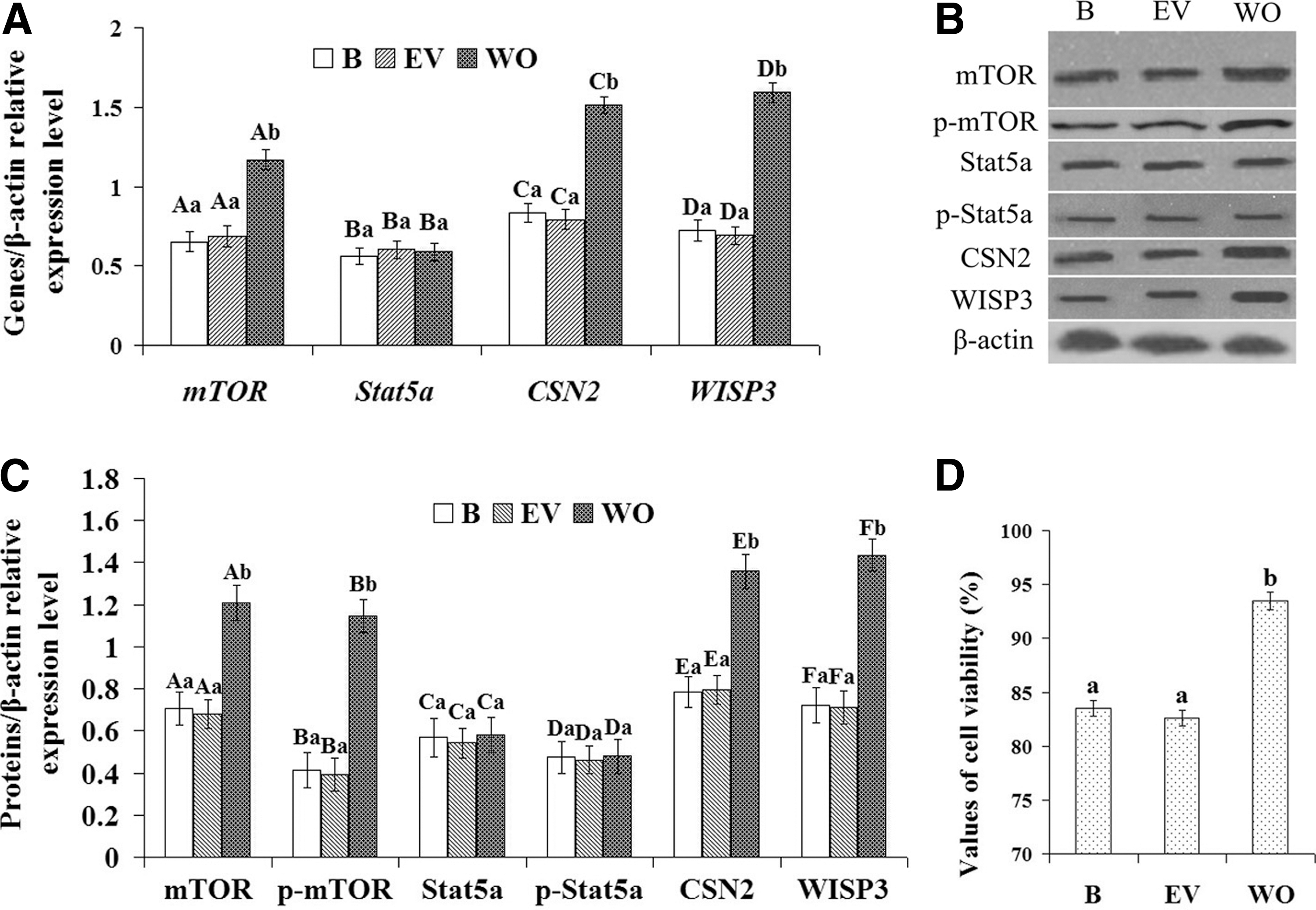
Effect of WISP3 overexpression on CSN2 synthesis in DCMECs and cell viability of DCMECs.
WISP3 knockdown suppressed both cell growth and the expression of milk protein synthesis-associated proteins
Using siRNA targeted to WISP3, we examined the effect of WISP3 inhibition on both cell growth and the expression of mTOR, p-mTOR, Stat5a, p-Stat5a, and CSN2 in DCMECs in three cell groups: the untransfected group (B), the transfected negative control RNA group (NC), and the WISP3 siRNA group (WI). After knockdown of WISP3, the expression of mTOR, p-mTOR, CSN2, and WISP3 was detected using qRT-PCR and WB at 24 h, and cell viability was examined using a CASY model TT cell counter. qRT-PCR results showed that the mRNA expression of mTOR, CSN2, and WISP3 was downregulated in the WI group compared with the B and NC groups (Fig. 3A, p<0.01); however, the expression of Stat5a was unchanged (Fig. 3A, p>0.05). WB results showed that the expression of mTOR, p-mTOR, CSN2, and WISP3 was also significantly decreased (Fig. 3B, p<0.01); however, the expression of Stat5a and p-Stat5a was unchanged (Fig. 3B, p>0.05), which was consistent with the qRT-PCR results. Cell viability results showed that compared with the B and EV groups, cell viability was significantly decreased (Fig. 3D, p<0.01) in the WI group. These data indicated that WISP3 silencing may suppress CSN2 synthesis and cell viability in DCMECs, and this function may relate to the mTOR signaling pathway, but not the Jak2/Stat5a signaling pathway.
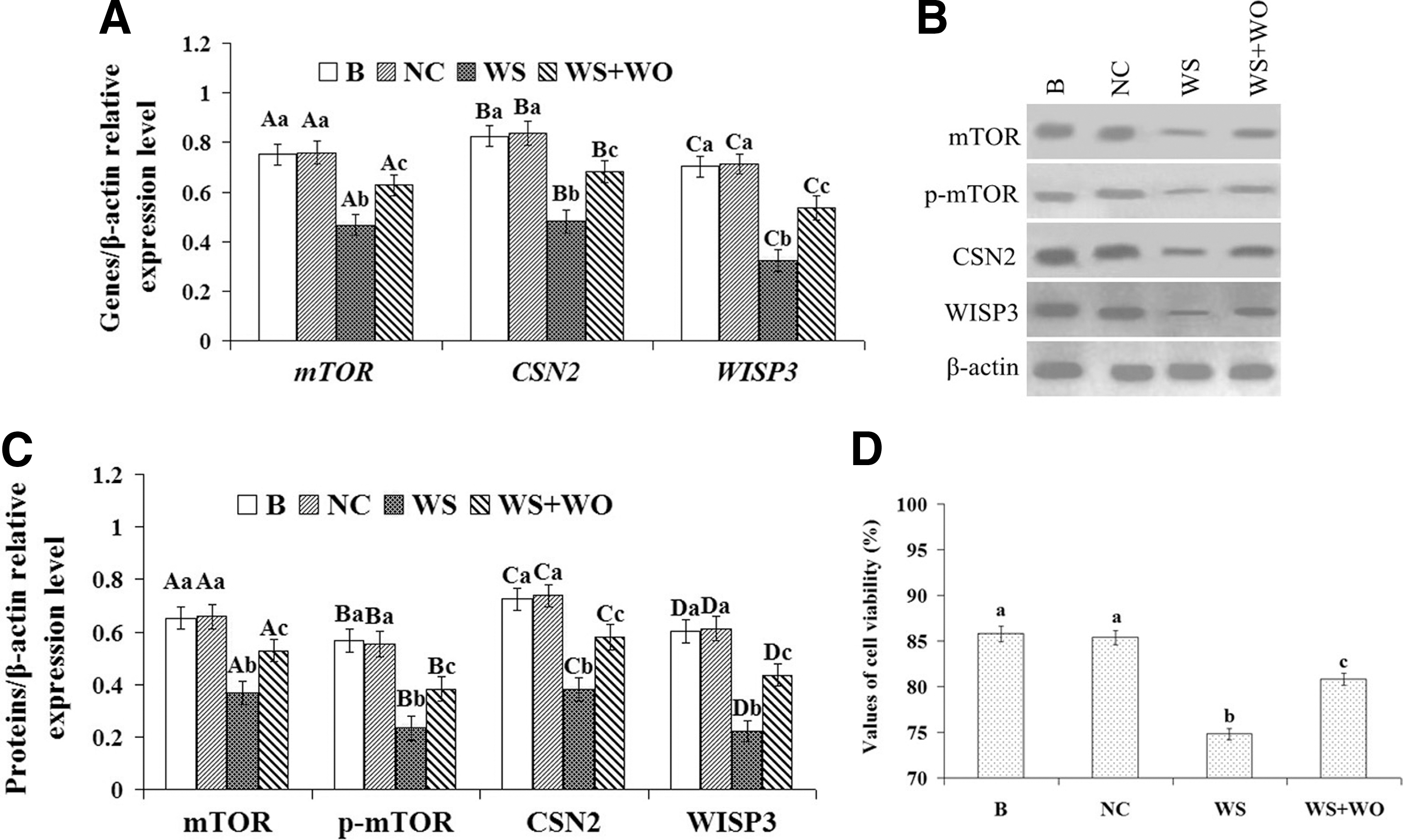
Effect of WISP3 silencing on CSN2 synthesis in DCMECs and cell viability of DCMECs.
In addition, as a complementation experiment, the cells were cotransfected with WISP3-specific siRNA and pGW, and the expression of WISP3, mTOR, p-mTOR, and CSN2 in DCMECs and the cell viability of DCMECs were tested. The results showed that after being treated with cotransfection (WS/WO), the expression of WISP3, mTOR, p-mTOR, and CSN2 in DCMECs and the cell viability of DCMECs were significantly increased (Fig. 3E–H, p<0.05) compared with the cells treated with WISP3-specific siRNA (WS), but significantly decreased (Fig. 3E–H, p<0.05) compared with the cells that were not treated (B). These data indicated that WISP3 may regulate CSN2 synthesis and cell viability through the mTOR signaling pathway.
WISP3 is a regulatory factor for CSN2 synthesis and cell growth regulated by Lys
To investigate the importance of Lys-regulated WISP3 for CSN2 synthesis and cell growth, related gene expression, protein expression, and cell viability experiments were completed using four cell groups: the normal cultured group (B), the Lys-treated normal culture group (B+Lys), the Lys-treated transfected negative control RNA group (NC+Lys), and the Lys-treated transfected WISP3 siRNA group (WS+Lys). The results showed that the expression of CSN2, mTOR, p-mTOR, and WISP3 and cell viability were significantly increased (Fig. 4A–D, p<0.01) in Lys-treated groups compared with group B. In group WS+Lys, the expression of CSN2, mTOR, p-mTOR, and WISP3 and cell viability were significantly increased (Fig. 4A–D, p<0.01) compared with group B, but significantly decreased (Fig. 4A–D, p<0.01) compared with group B+Lys, suggesting that in the process of CSN2 synthesis and cell growth regulated by Lys, the Lys signal was mediated, at least in part, by WISP.
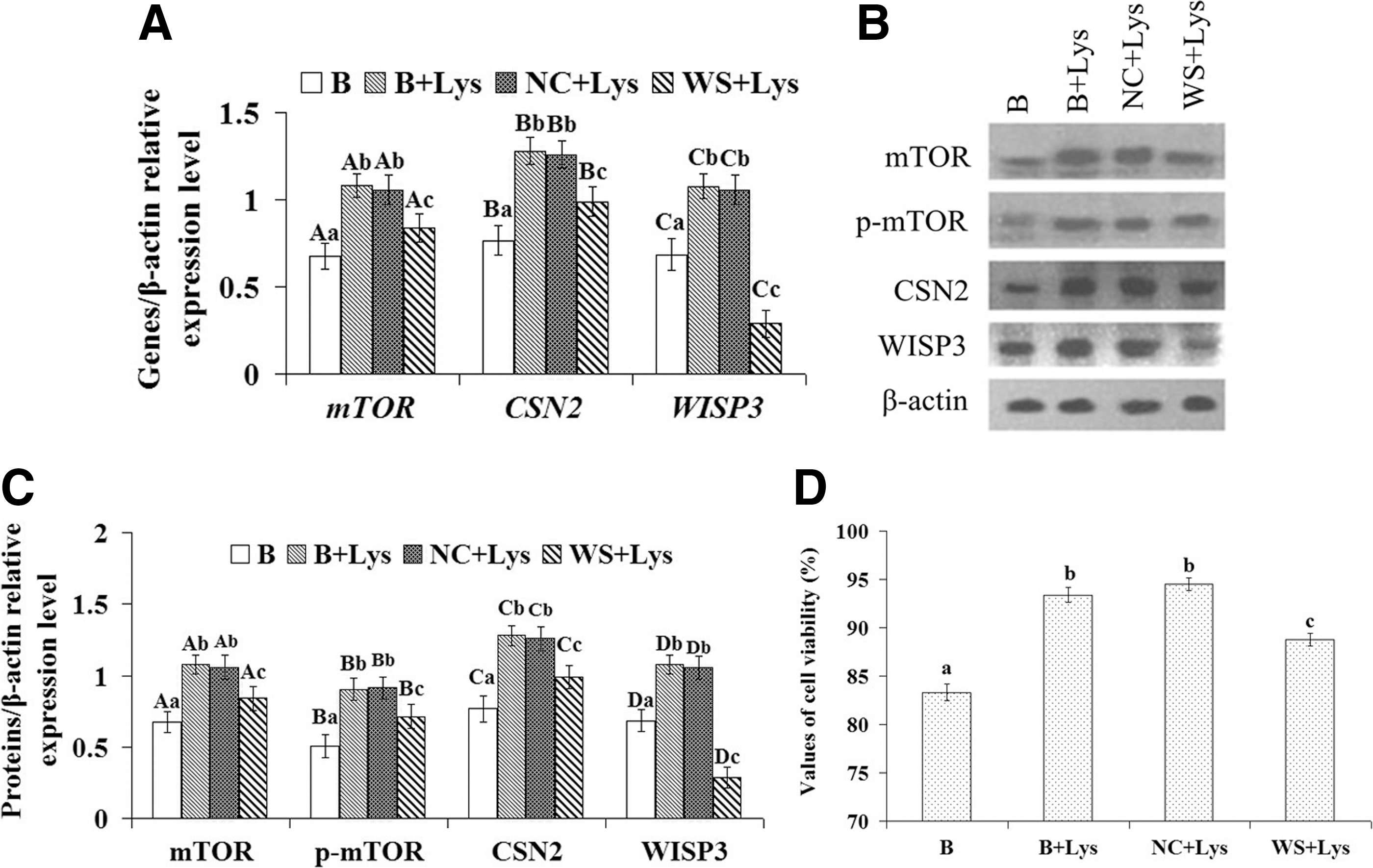
Effect of Lys stimulation and/or WISP3 silencing on the expression of CSN2, mTOR, p-mTOR, and WISP3 and cell viability in four groups of DCMECs: the normal cultured group (B), the Lys-treated normal culture group (B+Lys), the Lys-treated transfected negative control RNA group (NC+Lys), and the Lys-treated transfected WISP3 siRNA group (WS+Lys).
WISP3 promoted mTOR expression by inhibiting GSK3β expression
In the WNT signaling pathway, GSK3β is the downstream protein of Wnt. In a previous report by Porstmann et al. (2009), GSK3β is also shown to be an inhibitor of the mTOR/S6K1 signaling pathway. To investigate whether the WISP3-mediated increase in mTOR activation was related to GSK3β, related gene expression, protein expression, and cell viability experiments were completed using four cell groups: the untransfected group (B), the transfected empty plasmid group (EV), the WISP3-overexpressing group (WO), and the WISP3/GSK3β-overexpressing group. The results showed that the expression of mTOR and p-mTOR and cell viability were significantly increased (Fig. 5A–D, p<0.01) in cell group WO, while the expression of GSK3β was significantly decreased (Fig. 5A–C, p<0.01). However, when GSK3β and WISP3 were overexpressed together, mTOR and p-mTOR expression and cell viability were significantly decreased (Fig. 5A–D, p<0.01) compared with group WO, and the protein expression of mTOR and p-mTOR and cell viability could not be upregulated by WISP3 overexpression. These results suggested that WISP3 may increase mTOR activation by inhibiting GSK3β.

Effect of WISP3 and WISP3/GSK3β overexpression and silencing on the expression of CSN2, mTOR, p-mTOR, and WISP3 in DCMECs and cell viability of DCMECs.
In addition, related gene expression, protein expression, and cell viability were tested after the cells were treated with WISP3-specific siRNA (WS) and WISP3-specific siRNA/GSK3β-specific siRNA (WS+GS). The results showed that the expression of mTOR and p-mTOR and cell viability were significantly decreased (Fig. 5E–H, p<0.01) in the cell group WS, while the expression of GSK3β was significantly increased (Fig. 5E–H, p<0.01). However, when GSK3β and WISP3 were inhibited together, mTOR and p-mTOR expression and cell viability were significantly increased (Fig. 5, p<0.01) compared with group WS, and the protein expression of mTOR and p-mTOR and cell viability could be upregulated by GSK3β inhibition. These results suggested that WISP3 may increase mTOR activation by inhibiting GSK3β.
Discussion
To study dairy cow mammary lactation signaling pathways, use of a lactating DCMEC model is the key. CK18 and CSN2 are important marker proteins of lactating DCMECs (Jedrzejczak and Iwona Szatkowska, 2014). CK18 is an epithelial cell marker (Pantschenko et al., 2000; Zhao et al., 2010), and CSN2 is the main composition of the milk proteins; CSN2 synthesis in cells is a sign of their ability to lactate (Hu et al., 2009). In this study, primary DCMECs were cultured and purified, then the expression of CK18 and CSN2 was tested using IF. The results showed positive expression of CK18 and CSN2 in purified cells, suggesting that the purified DCMECs were capable of lactation.
Lys is one of the eight essential amino acids required by the body and it is a necessary building block for all proteins. In addition to this well-known function, in recent years, studies have found that Lys is also an important regulator of various signaling pathways, cell growth, and protein synthesis (Appuhamy et al., 2011a; Lu et al., 2013). In this study, after treatment with 1.2 mM Lys for 24 h, the expression of CSN2 in DCMECs and cell viability were assessed. The results showed that the expression of CSN2 in DCMECs and cell viability were significantly increased (p<0.01) after Lys treatment, suggesting that an enhanced lactating cell model regulated by Lys was established. The expression of WISP3 in this model was then detected. The results showed that CSN2 and WISP3 were both upregulated by Lys, suggesting that WISP3 may be involved in the Lys-mediated regulation of CSN2 synthesis and cell growth.
WISP3 is a member of the CCN protein family and it can regulate a variety of cellular activities, such as cell growth and protein synthesis, through its involvement in many cell signaling pathways. In this study, by overexpressing and inhibiting WIPS3, we found that WISP3 can positively regulate CSN2 synthesis and cell viability in DCMECs. The main signaling pathways of milk protein expression are the Jak2/Stat5 (He et al., 2012; Jiang et al., 2012) and mTOR signaling pathways (Gingras et al., 1999; Wullschleger et al., 2006; Yang et al., 2008; Ma and Blenis, 2009). Notably, the mTOR/S6K1 signaling pathway also plays an important role in cell growth. In this study, the overexpression of WISP3 increased not only cell viability and the expression of CSN2 but also the expression of mTOR and p-mTOR (p<0.01); however, the expression of Stat5a and p-Stat5a was unchanged (p<0.01). These results suggested that WISP3 could positively regulate CSN2 synthesis and cell viability in DCMECs, and in this process, CSN2 synthesis may be involved in mTOR signaling, but not Jak2/Stat5 signaling. Additionally, after adding Lys to the DCMECs, we found that Lys positively regulated CSN2 synthesis and cell viability, and in this process, the Lys signal was mediated, at least in part, by WISP.
GSK3β is a serine/threonine kinase (Aparicio et al., 2010; Jacobs et al., 2012), which plays an important physiological role in the regulation of a wide range of cellular functions, including differentiation, growth, apoptosis, and cell response to stimuli (Dong et al., 2005; Stein et al., 2011; Chen et al., 2012). The GSK3β was inhibited in the PI3K/Akt pathway, Wnt/GSK3 signaling, and ERK pathway (Zhang et al., 2006; Fuentealba et al., 2007; Kiyonari et al., 2010; Wu and Pan, 2010). The CCN proteins can regulate crucial biological processes, including cell differentiation, proliferation, adhesion, migration, and angiogenesis, and some important signaling pathways, including Wnt signaling, PI3K/Akt pathway, and ERK pathway (Chen et al., 2004; Gao et al., 2004; Leask and Abraham, 2006; Shi-wen et al., 2006; Rey and Ellies, 2010). However, the mechanism for WISP3-mediated downregulation of GSK3β expression was unknown.
In this study, we found that when WISP3 was overexpressed in DCMECs, the expression of mTOR, p-mTOR, and CSN2 was significantly increased and the expression of GSK3β was significantly decreased. However, when GSK3β and WISP3 were overexpressed together, the expression of mTOR, p-mTOR, and CSN2 was significantly decreased compared with the cells overexpressing only WISP3, and the expression of the three proteins could not be upregulated by WISP3 overexpression. These results suggested that WISP3 increases mTOR activation by inhibiting GSK3β. Because the gene transcription and post-transcriptional modification processes between the endogenous and exogenous GSK3β are different, the regulation of GSK3β by Wnt signaling, PI3K/Akt pathway, or ERK pathway may be in transcription and post-transcriptional modifications of the GSK3β gene expression, so the overexpression of WISP3 could inhibit expression of endogenous GSK3β, but not exogenous GSK3β, were discussed in the revised study, but the mechanism is still unknown.
Conclusion
In summary, in DCMECs, WISP3 can respond to amino acid signaling and can upregulate CSN2 gene expression and cell growth through the mTOR signaling pathway, which WISP3 can activate by inhibiting the expression of GSK3β. Ultimately, amino acid can upregulate milk protein synthesis through a signaling process that is transmitted, at least in part, through WISP3.
Footnotes
Acknowledgment
This work was supported by the National Key Basic Research Program of China (Project No. 31472169).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.