
Abstract
Curcumin, a traditional Chinese derivative from the rhizomes of Curcuma longa, is beneficial to health by modulating lipid metabolism and suppressing atherogenesis. A key part of atherosclerosis is the failure of macrophages to restore their cellular cholesterol homeostasis and the formation of foam cells. In this study, results showed that curcumin dramatically increased the expression of ATP-binding cassette transporter 1 (ABCA1), promoted cholesterol efflux from THP-1 macrophage-derived foam cells, and reduced cellular cholesterol levels. Curcumin activated AMP-activated protein kinase (AMPK) and SIRT1, and then activated LXRα in THP-1 macrophage-derived foam cells. Inhibiting AMPK/SIRT1 activity by its specific inhibitor or by small interfering RNA could inhibit LXRα activation and abolish curcumin-induced ABCA1 expression and cholesterol efflux. Thus, curcumin enhanced cholesterol efflux by upregulating ABCA1 expression through activating AMPK-SIRT1-LXRα signaling in THP-1 macrophage-derived foam cells. This study describes a possible mechanism for understanding the antiatherogenic effects of curcumin on attenuating the progression of atherosclerosis.
Introduction
A
Curcumin, a major bioactive component of turmeric spice, impedes tumorigenesis and diabetes through its antioxidant and anti-inflammatory properties. Curcumin also exhibits beneficial effects in modulating several molecules, such as lipids and cholesterol, which are involved in the development of cardiovascular disease (Gupta et al., 2013; Liu et al., 2013). Moreover, limited animal studies have demonstrated that curcumin reduces atherogenesis. Olszanecki et al. (2005) reported that low-dose curcumin (0.3 mg/mouse/day) reduces the progression of atherosclerosis in apoE/low-density lipoprotein receptor (LDLR)-double knockout mice that were fed a high-fat diet. In a mouse macrophage study, Zhou et al. (2014) found that curcumin prevents a decrease in thrombospondin-4 induced by oxidized low-density lipoprotein (ox-LDL) to inhibit cell interaction. Furthermore, Dong et al. (2011) found that curcumin can affect the cholesterol efflux from adipocytes by regulating the PPAR-LXR-ABCA1 pathway. On the basis of this study, Zhao et al. (2012) demonstrated that curcumin reduces cholesterol accumulation in foam cell development by downregulating SR-A and upregulating ATP-binding cassette transporter 1 (ABCA1) using proteasome- and liver X receptor alpha (LXRα)-dependent pathways, respectively. Although its mechanism in the prevention of atherosclerosis was increasingly detected, curcumin's antiatherogenic mechanism is still unclear, especially the molecular mechanism by which it regulates cholesterol efflux by upregulating ABCA1 expression in macrophage foam cells. The PPARγ-LXRα axis is a major pathway to induce cholesterol efflux in macrophages, and curcumin demonstrates promoted cholesterol efflux by upregulating ABCA1 using the LXRα-dependent pathway (Zhao et al., 2012). New molecular mechanisms of curcumin in inhibiting the formation of foam cells have been discovered.
Numerous mechanisms may mediate curcumin's antiatherogenic effect, including the alteration of lipids and cholesterol in the liver and circulation, as well as the expression of inflammatory cytokines (Zhai et al., 2015). Furthermore, this component may be activated by AMP-activated protein kinase (AMPK) signaling (Jiménez-Flores et al., 2014). AMPK, a member of a metabolite-sensing protein kinase family, is implicated in the development of atherosclerosis. AMPK activation exerts anti-inflammatory effects in the endothelium by reducing TNF-α-stimulated monocyte adhesion to human aortic endothelial cells (Xu and Si, 2010). AICAR is an AMPK activator that reduces reactive oxygen species and inhibits macrophage proliferation, which decreases the level of circulating ox-LDL (Wong et al., 2009). In addition, several studies indicate that AMPK activation induces the expression of ATP-binding cassette transporters to reduce macrophage-derived foam cell formation (Li et al., 2010). These reports support that AMPK possesses an antiatherosclerotic factor. SIRT1 is a downstream target of AMPK that can activate LXRα, increase ABCA1 expression, promote cholesterol efflux, and reduce foam cell formation (Li et al., 2007).
In this study, we tested the hypothesis that curcumin regulates the expression of ABCA1 through stimulation of AMPK-SIRT1-LXRα signaling. We conducted experiments to investigate the relationship between curcumin and ABCA1 expression in THP-1 macrophage-derived foam cells. We also identified the role of AMPK-SIRT1-LXRα signaling on increasing ABCA1 expression by curcumin. Results revealed that curcumin enhanced cholesterol efflux by upregulating ABCA1 expression through activating AMPK-SIRT1-LXRα signaling in THP-1 macrophage-derived foam cells, which suggests the protective function of curcumin against atherosclerosis.
Materials and Methods
Materials
Curcumin was purchased from Sigma (St. Louis, MO). TRIzol reagent (Invitrogen, Carlsbad, CA), BCA protein assay reagent (Pierce Chemical, Rockford), ReverAid™ first-strand cDNA synthesis kit (Fermentas, Burlington, Canada), LXRα agonist T0901317 rabbit monoclonal anti-ABCA1, anti-β-actin and rabbit polyclonal anti-LXRα, anti-AMPK, antiphospho-AMPK (anti-p-AMPK), anti-SIRT1, and antiphospho-SIRT1 (anti-p-SIRT1) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). SIRT1 inhibitor, EX 527, was purchased from Selleckchem (Houston, TX), and AMPK inhibitor, compound C, was purchased from Boster (Wuhan, China). Horseradish peroxidase-conjugated goat anti-rabbit polyclonal antibody was purchased from CWBIO (Beijing, China). All other reagents were obtained in the best grade available from commercial sources.
Cell culture
Human THP-1 cells were cultured in RPMI1640 medium (Hyclone, Logan, UT) with 10% fetal bovine serum (Invitrogen), 20 μg/mL penicillin, and 20 μg/mL streptomycin and maintained at 37°C in a humidified atmosphere of 5% CO2, and then treated with 100 nM phorbol 12-myristate 13-acetate (Sigma) for 24 h to induce differentiation into macrophages, and then the medium was replaced with a serum-free medium containing ox-LDL (50 mg/mL) for 48 h to fully differentiate THP-1 cells to foam cells.
Cell viability assay
THP1-derived macrophage foam cells (8×103/mL) were seeded into each of the 96-well culture plates overnight and kept in a humidified atmosphere of 5% CO2 and 95% air at 37°C. After 12 h of incubation, the medium was exchanged for serum RPMI1640 medium and added to curcumin at different concentrations (0, 5, 10, 20, 40, 80 μM) or treated with curcumin for different times (0, 6, 12, 24, and 48 h). Meanwhile, cells without any treatment were used as a control. Following this, the culture medium was removed, and 5 mg/mL methyl thiazolyl tetrazolium (MTT; CWBIO) was added to each well. The plates were then incubated for 4 h at 37°C. The culture medium was replaced with 150 μL of dimethyl sulfoxide (CWBIO) per well, and the absorbance at the 490 nm wavelength was measured using a 2104 EnVision Multilabel Reader (PerkinElmer, Inc., Waltham, MA). Cell viability (%)=[(curcumin OD-bland group OD)]/[(control group OD-bland group OD)]×100%.
Oil red staining
Cell lipid accumulation was measured through staining of neutral fats and cholesterol esters (CEs) with Oil Red O. THP1-derived macrophage foam cells were seeded into six-well plates with slides at a density of 4×105 cells/cm2, and then treated with curcumin in different concentrations (0, 5, 10, and 20 μM) or pretreated with 10 μM curcumin for different times (0, 6, 12, and 24 h), after which the cells were rinsed with phosphate-buffered saline (PBS) thrice, and then fixed with 10% formalin for 5 min at room temperature and incubated in fresh formalin for at least 1 h. The cells were then rinsed again with 60% isopropanol, and then incubated with fresh filtered Oil Red O solution (60% saturated Oil Red O/40% deionized water) for 15 min. For analysis, slides were then washed in isopropanol for 10 min, rinsed in tap water, counterstained with hematoxylin, and mounted in glycerol solution, and then images of cells were captured using a light microscope.
Preparation of ox-LDL
The human LDL was purchased from Sigma, and it was diluted by PBS. LDL was oxidized with 5 μM CuSO4 at 37°C for 24 h, and then transferred into ethylenediaminetetraacetic acid (EDTA; 200 μmol/L) in PBS for 24 h at 4°C. Subsequently, oxidation was stopped by extensive dialysis against PBS with 0.01% EDTA and sterilized by filtration. LDL oxidation was confirmed by thiobarbituric acid reaction substances with malondialdehyde as the standard (Wei et al., 2013).
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted from macrophages using TRIzol reagent following the manufacturer's instructions (Invitrogen). First-strand cDNA synthesis was done with 1 μg total RNA using a TaqMan Reverse Transcription Reagent Kit (Applied Biosystems, Foster city, CA). Real-time quantitative polymerase chain reaction (PCR) was performed with 2 μg cDNA using SYBR Green detection chemistry on a Roche LightCycler real-time PCR system (Roche Diagnostics, Mannheim, Germany). The primers used to amplify specific gene products were as follows: glyceraldehyde-3-phosphate dehydrogenase (GAPDH) sense 5′-GTG GGG CGC CCC AGG CAC CA-3′, antisense 5′-CTC CTT AAT GTC ACG CAC GAT TC-3′; human ABCA1 sense 5′-GGAGCAGGCAATCATCAG-3′, antisense 5′-ACACGGACAGGAAGACAA-3′; and LXRα sense 5′-TCTGCGGTGGAGCTGTGGAA-3′, antisense 5′-TGACGCTGGGCGGAAGAAT-3′ (Zeng et al., 2013). All data were evaluated with the Roche LightCycler Run 5.32 software (Roche Diagnostics). Melt curve analyses of all real-time PCR products were performed as shown to ensure production of a single DNA duplex. The relative expression level was determined by normalizing the expression of each gene to that of the β-actin gene by the 2−ΔΔCt method.
HPLC assays
High-performance liquid chromatography (HPLC) analysis was conducted as described previously (Lin et al., 2014). In short, cells were washed thrice with PBS. The appropriate volume (usually 1 mL) of 0.5% NaCl was added to∼50–200 mg cellular proteins per mL. The cells were sonicated using an ultrasonic processor for 2 min. The protein concentration in the cell solution was measured using a BCA kit. A 0.1 mL aliquot of the cell solution (containing 5–20 μg protein) was used to measure free cholesterol, and a different aliquot was used to measure total cholesterol. Free cholesterol was dissolved in isopropanol (1 mg cholesterol/mL) and stored at −20°C as stock solution. A cholesterol standard calibration solution ranging from 0 to 40 mg cholesterol/mL was obtained by diluting the cholesterol stock solution in the same cell lysed buffer. Then, 0.1 mL of each sample (cholesterol standard calibration solutions or cell solutions) was supplemented with 10 mL reaction mixture, including 500 mM MgCl2, 500 mM Tris-HCl (pH 7.4), 10 mM dithiothreitol, and 5% NaCl. A total of 0.4 U of cholesterol oxidase in 10 mL 0.5% NaCl was added to each tube for free cholesterol determination, while 0.4 U of cholesterol oxidase and 0.4 U of cholesterol esterase were added to each tube for total cholesterol measurement. The total reaction solution in each tube was incubated at 37°C for 30 min, and 100 mL of methanol: ethanol mixture (1:1) was then added to stop the reaction. Each solution was kept cold for 30 min to allow protein precipitation, and then centrifuged at 1500 rpm for 10 min at 15°C. Then, 10 mL of supernatant was applied onto a chromatograph system (PerkinElmer, Inc.), which consisted of a PerkinElmer series 200 vacuum degasser, pump, PerkinElmer series 600 LINK, PerkinElmer series 200 UV/vis detector, and a Discovery C-18 HLPC column (Supelco, Inc., Bellefonte, PA). The column was eluted using an isopropanol:n-heptane:acetonitrile mixture (35:13:52) at a flow rate of 1 mL/min for 8 min. Absorbance at 216 nm was monitored. Data were analyzed using TotalChrom software (PerkinElmer, Inc.).
Western blot analysis
The cells were harvested and lysed. Cell lysate was separated in sodium dodecyl sulfate–polyacrylamide electrophoresis gels (8%) and transferred to a polyvinylidene difluoride membrane. The membranes were blocked with 5% milk, and then incubated with ABCA1 (1:300 dilution), AMPK (1:300 dilution), p-AMPK, SIRT1 (1:400 dilution), p-SIRT1 (1:400 dilution), LXRα (1:400 dilution), and β-actin (1:500 dilution) at 4°C overnight, then with horseradish peroxidase-conjugated secondary antibodies (goat anti-rabbit). Visualization was done using the enhanced chemiluminescence detection system from Amersham Biosciences (Amersham, United Kingdom).
Assay of apoAI-mediated cholesterol efflux
Assay of apoAI-mediated cholesterol efflux was conducted as described previously (Liu et al., 2013; Lin et al., 2014). Briefly, Cells were cultured as described above, and then labeled with 0.2 μCi/mL of [3H] cholesterol for 24 h. After 24 h, the cells were washed with fresh media and curcumin was added. The cells were washed thrice with PBS and incubated overnight in RPMI 1640 medium containing 0.1% (wt/vol) bovine serum albumin (BSA) with 10. Equilibrated [3H] cholesterol-labeled cells were washed with PBS and incubated in 2 mL of the efflux medium containing RPMI 1640 medium and 0.1% BSA with 25 μg/mL human plasma apoA-1. A 150 mL sample of the efflux medium was obtained at designated times and passed through a 0.45-μm filter to remove any floating cells. The cell monolayer was washed with PBS and extracted with 0.15 M NaOH to determine cellular total [3H] radioactivity. The medium and cell-associated [3H] cholesterol were then measured by liquid scintillation counting. Percent efflux was calculated using the following equation: [total media counts/(total cellular counts+total media counts)]×100%.
Transfection of small interfering RNA
Small interfering RNA (siRNA) targeting LXRα was purchased from Santa Cruz Biotechnology, and nonsilencing control siRNA was purchased from Biology Engineering Corporation (Shanghai, China). THP-1 macrophage-derived foam cells (2×106 cells/well) were transfected using Lipofectamine 2000 (Invitrogen) as previously described (Liu et al., 2013).
Statistical analysis
All results are expressed as the mean±standard deviation (SD) from three independent experiments, and data were analyzed by one-way ANOVA and Student's t-test using SPSS 18.0 software, and p<0.05 was considered statistically significant.
Results
Effect of curcumin on the viability of THP-1 macrophage-derived foam cells
We initially examined the cytotoxicity of curcumin in THP-1 macrophage-derived foam cells. MTT assay was used to measure cell viability. THP-1 macrophage-derived foam cells were treated with different curcumin concentrations (0, 5, 10, 20, 40, and 80 μM) for 24 h. As shown in Fig. 1A, curcumin cytotoxicity was negligible at 0–20 μM concentrations in THP-1 macrophage-derived foam cells. Cells treated with 20 μM curcumin were observed at different times (0, 3, 6, 12, 24, and 48 h). As shown in Figure 1B, the cytotoxicity of curcumin was negligible at 0–24 h. These results indicate that treatment with 0–20 μM curcumin for 0–24 h was not toxic to THP-1 macrophage-derived foam cells.
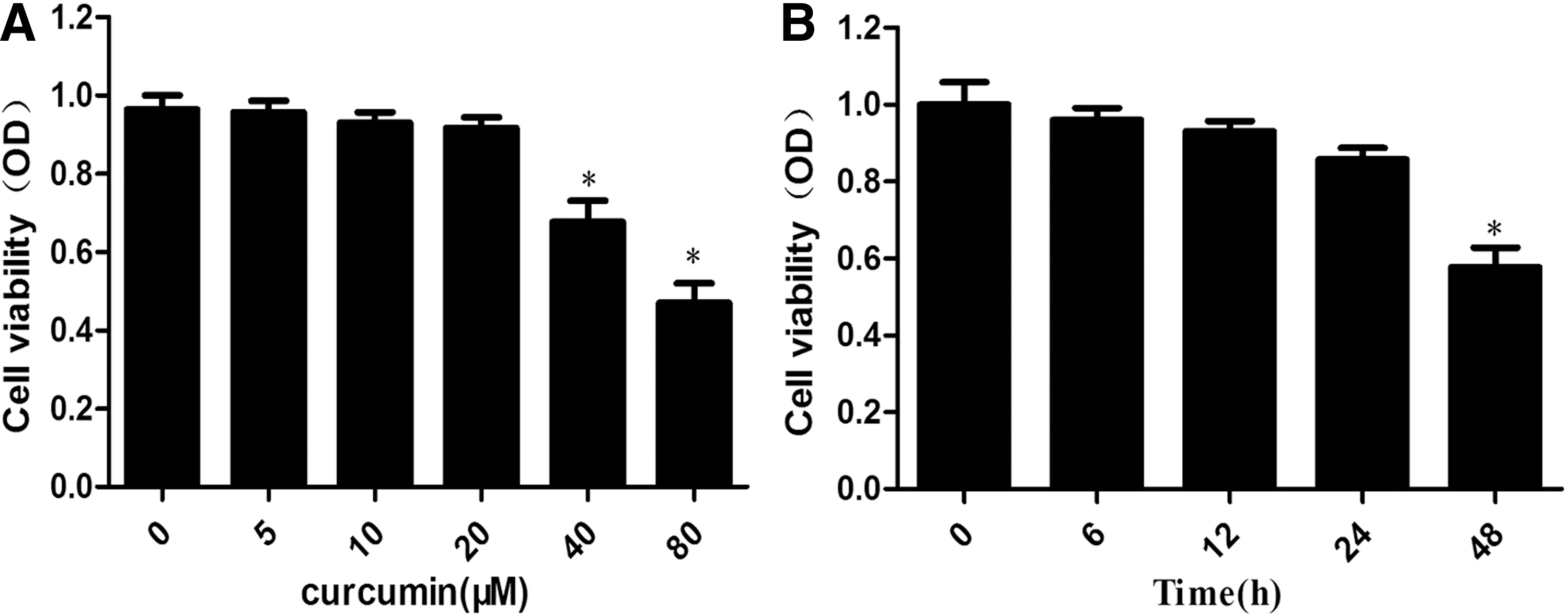
Cytotoxicity of curcumin on macrophages.
Curcumin promotes cholesterol efflux and inhibits the formation of foam cells in THP-1 macrophage-derived foam cells
The formation of macrophage foam cells is essential in atherosclerotic lesion progression. We initially examined the effect of curcumin on cellular cholesterol levels in THP-1 macrophage-derived foam cells. Table 1 shows that curcumin significantly reduced total cholesterol, free cholesterol, and CE at 10 and 20 μM concentrations. We then performed a time course experiment using 10 μM curcumin and found that curcumin started to significantly reduce cellular cholesterol and CE levels at 12 h (Table 2). Then, we measured the intracellular lipid droplets in THP-1 macrophage-derived foam cells, which were incubated with different curcumin concentrations or 10 μM at various times. Figure 2A, B show that curcumin decreased the amount of intracellular lipid droplets in a time-dependent and dose-dependent manner. These findings demonstrate that curcumin may reduce cellular cholesterol accumulation to inhibit foam cell formation.
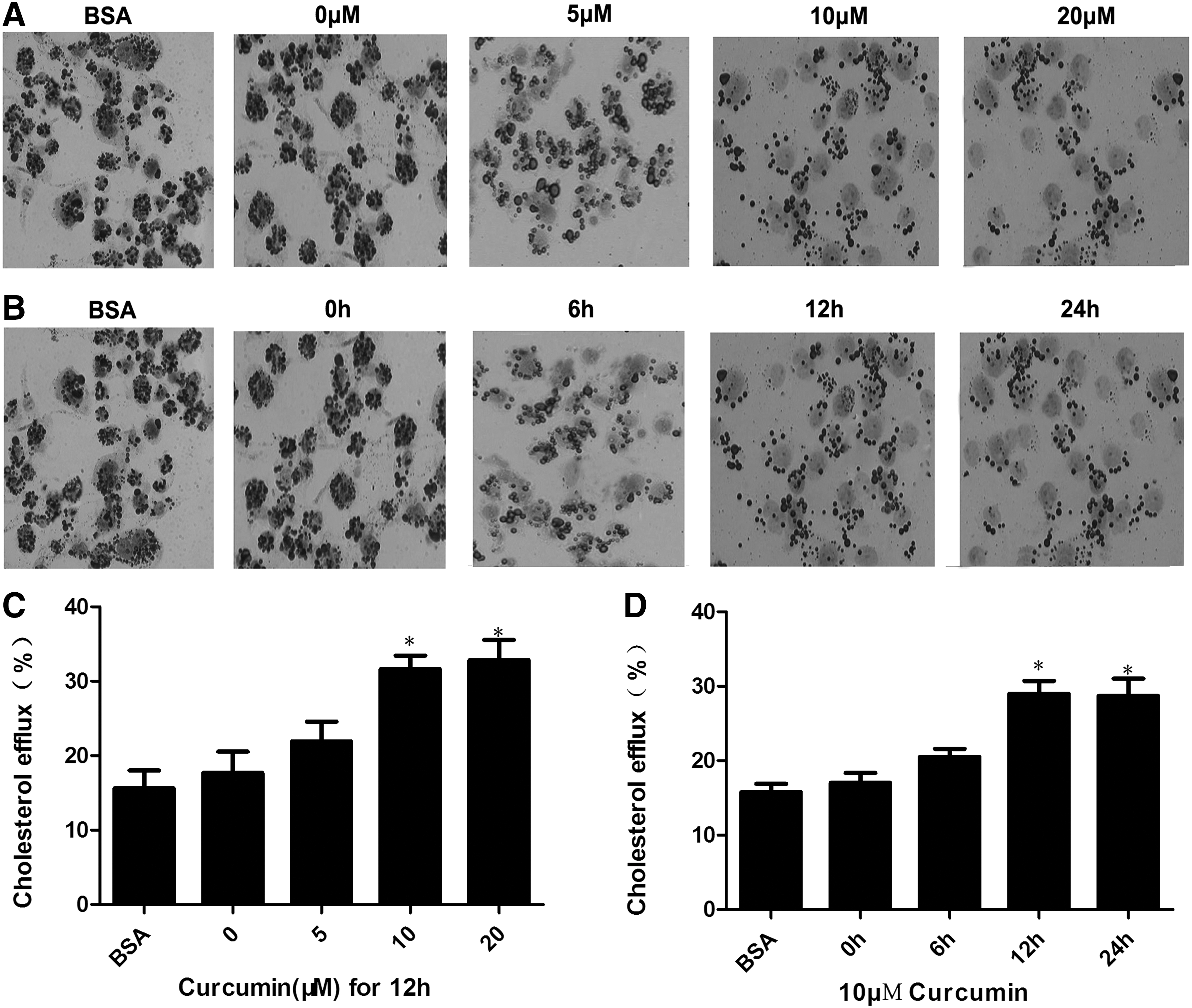
Curcumin reduced lipid droplet accumulation through promotion of cholesterol efflux in foam cells in a dose- and time-dependent manner.
THP-1 macrophage-derived foam cells were divided into five groups and cultured at 37°C in media containing BSA, 0, 5, 10, or 20 μM curcumin for 12 h. Cellular cholesterol and CE were extracted as described in the Materials and Methods section. HPLC was performed to determine the levels of cellular TC, FC, and CE. Results are expressed as mean±SD of three independent experiments performed in triplicate.
p<0.05 versus control.
BSA, bovine serum albumin; CE, cholesterol ester; FC, free cholesterol; HPLC, high-performance liquid chromatography; SD, standard deviation; TC, total cholesterol.
THP-1 macrophage-derived foam cells were divided into five groups and cultured at 37°C in media containing BSA or 10 μM curcumin at indicated times. Cellular cholesterol and CE were extracted as described in the Materials and Methods section. HPLC determined the levels of cellular TC, FC, and CE. Results are expressed as mean±SD of three independent experiments performed in triplicate.
p<0.05 versus control.
To investigate how curcumin decreased cellular cholesterol levels, we assessed the cholesterol efflux activity in the presence and absence of curcumin. Curcumin significantly increased the efflux of cholesterol at 10 and 20 μM concentrations (Fig. 2C). In the time course experiment, 10 μM curcumin significantly increased the cellular cholesterol efflux after 12 h of incubation (Fig. 2D). These results suggest that curcumin activated cholesterol efflux and reduced cellular cholesterol accumulation to inhibit foam cell formation.
Curcumin increases ABCA1 expression
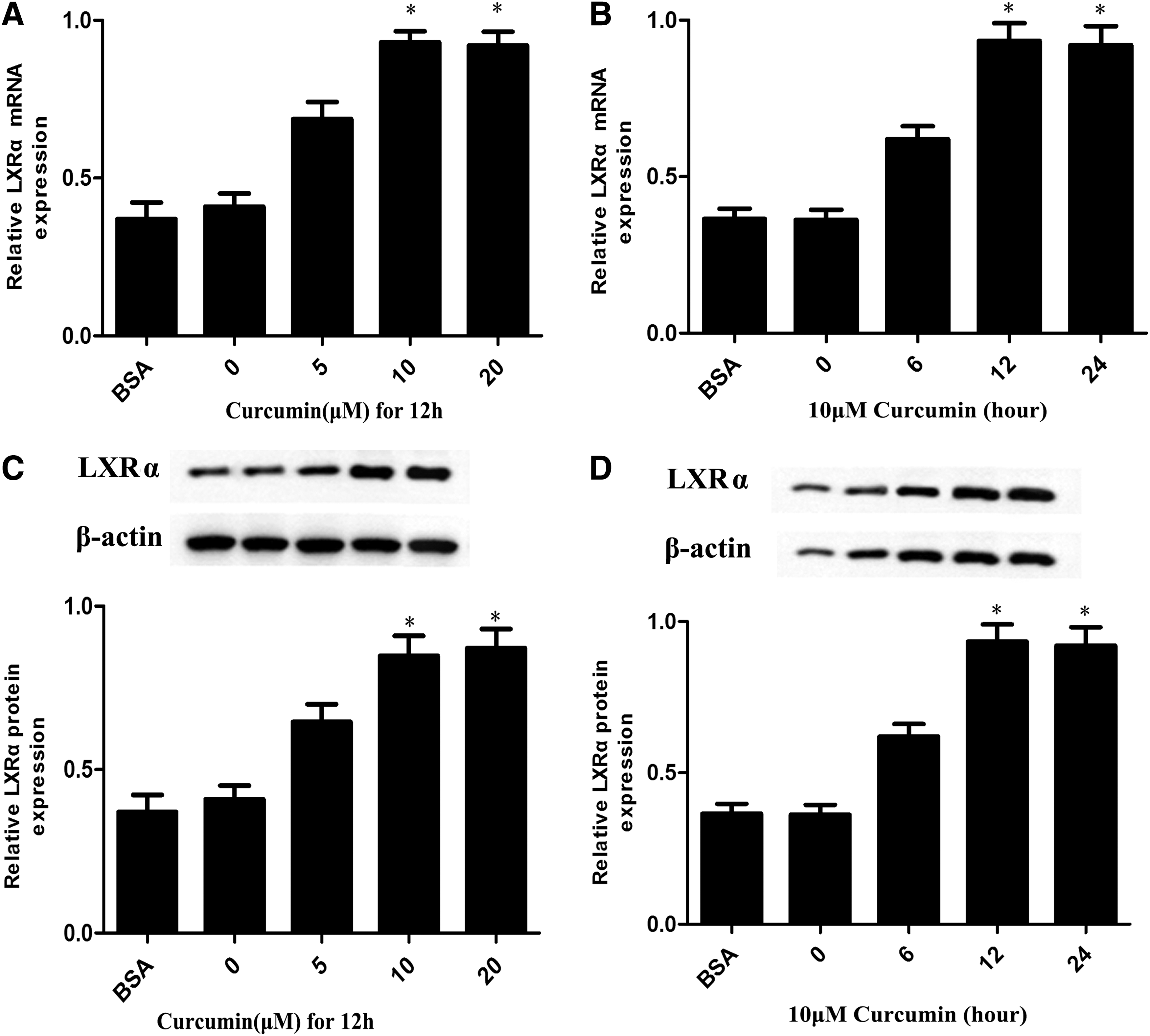
Previous studies demonstrated that ABCA1 is a key player in macrophage reverse cholesterol transport (RCT) and is critical for regulating cellular cholesterol homeostasis during transformation of foam cells (Iizuka et al., 2012; Yu et al., 2013). To study whether ABCA1 is involved in the effect of curcumin on cellular cholesterol levels, we investigated the impacts of curcumin on the mRNA and protein levels of ABCA1. Quantitative real-time PCR and western blot analysis were performed. Figure 3A, B shows that 10 μM curcumin and incubation time of up to 12 h had a significant effect on ABCA1 mRNA levels. Curcumin significantly increased ABCA1 protein levels at the 10 μM concentration (Fig. 3C) and 12 h incubation time (Fig. 3D). These results provide evidence of curcumin's effect on ABCA1 expression.

Effect of curcumin on the expression of ABCA1 in THP-1 macrophage-derived foam cells.
LXRα is involved in curcumin-induced ABCA1 upregulation
LXRs regulate the expression of ABCA1, which mediates phospholipid translocators and free cholesterol; this phenomenon enables cholesterol efflux from the macrophage to various acceptors, including nascent cholesterol-poor high-density lipoprotein (HDL), which is its central role in the regulation of reverse cholesterol transport (Breevoort et al., 2014). We initially detected the expression of LXRα in cells treated with different curcumin concentrations or in cells treated with 10 μM curcumin at different times. Figure 4A–D shows that curcumin increased the expression of LXRα in a time- and dose-dependent manner. We then investigated whether the effects of LXRα on the upregulation of ABCA1 expression were induced by curcumin in THP-1 macrophage-derived foam cells. We treated the cells with LXRα siRNA or LXRα agonist T0901317. As shown in Figure 5A, treatment with siRNA for LXRα downregulated LXRα expression by 89% (Fig. 5A). Moreover, Figure 5B, C shows that curcumin significantly increased ABCA1 expression, and similar results were also found in LXRα agonist-treated cells, but it is abolished by transfect LXRα siRNA. Meanwhile, cellular cholesterol efflux treated with a combination of curcumin and LXRα siRNA was decreased compared with those treated with curcumin alone (Fig. 5D). Thus, curcumin induced the upregulation of ABCA1 expression through LXRα.
Curcumin increases the expression of LXRα.
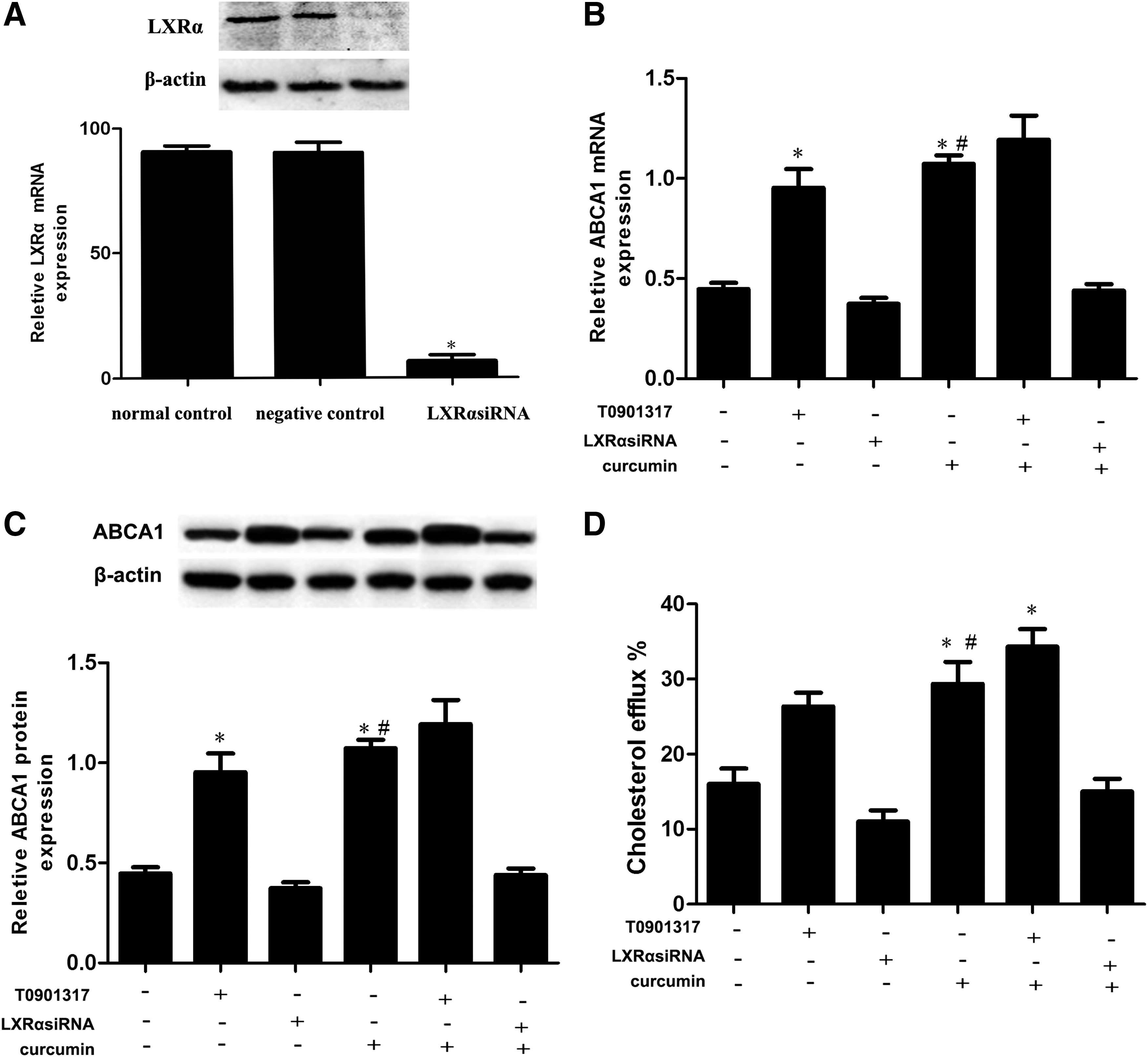
LXRα is involved in the upregulation of ABCA1 expression induced by curcumin.
Upregulation of ABCA1 expression induced by curcumin media through the AMPK-SIRT1 signaling pathway
AMPK is an energy sensor of metabolic homeostasis. Previous studies documented that AMPK activation plays an important role in atherosclerosis treatment through enhanced cholesterol efflux from foam cells (Li et al., 2010). SIRT1 is a cytoplasmic transcription factor that is activated by AMPK in response to cytokines (Li et al., 2007). It is a positive regulator of LXR proteins, serves as a nuclear receptor that functions as a cholesterol sensor, and regulates whole-body cholesterol as well as lipid homeostasis (Han et al., 2014). Numerous studies indicate that curcumin could activate AMPK in adipocytes and macrophages (Cao et al., 2014; Priyanka et al., 2014). In this study, THP-1 macrophage-derived foam cells were treated with 10 μM curcumin for 12 h. Western blot analysis showed that the expression of p-AMPK and p-SIRT1 protein increased after adding curcumin. The results demonstrate that curcumin can activate AMPK-SIRT1 signaling in THP-1 macrophage-derived foam cells (Fig. 6A–C). THP-1 macrophage-derived foam cells were treated with the specific inhibitors of AMPK to further investigate whether AMPK activation is involved in curcumin-induced ABCA1 expression. As shown in Figure 6D, E, cells treated with AMPK inhibitor downregulated LXRα and ABCA1 expression compared with cells treated with curcumin only. Meanwhile, cellular cholesterol efflux treated with AMPK inhibitor was significantly decreased compared with cells treated with curcumin only (Fig. 6F). p-SIRT1 expression was downregulated after treatment with AMPK inhibitor (Fig. 6E). Similar results were found when a genetic approach by transient AMPK knockdown was used. This result reveals that curcumin could increase ABCA1 expression and enhance cholesterol efflux through the AMPK-SIRT1 signal.
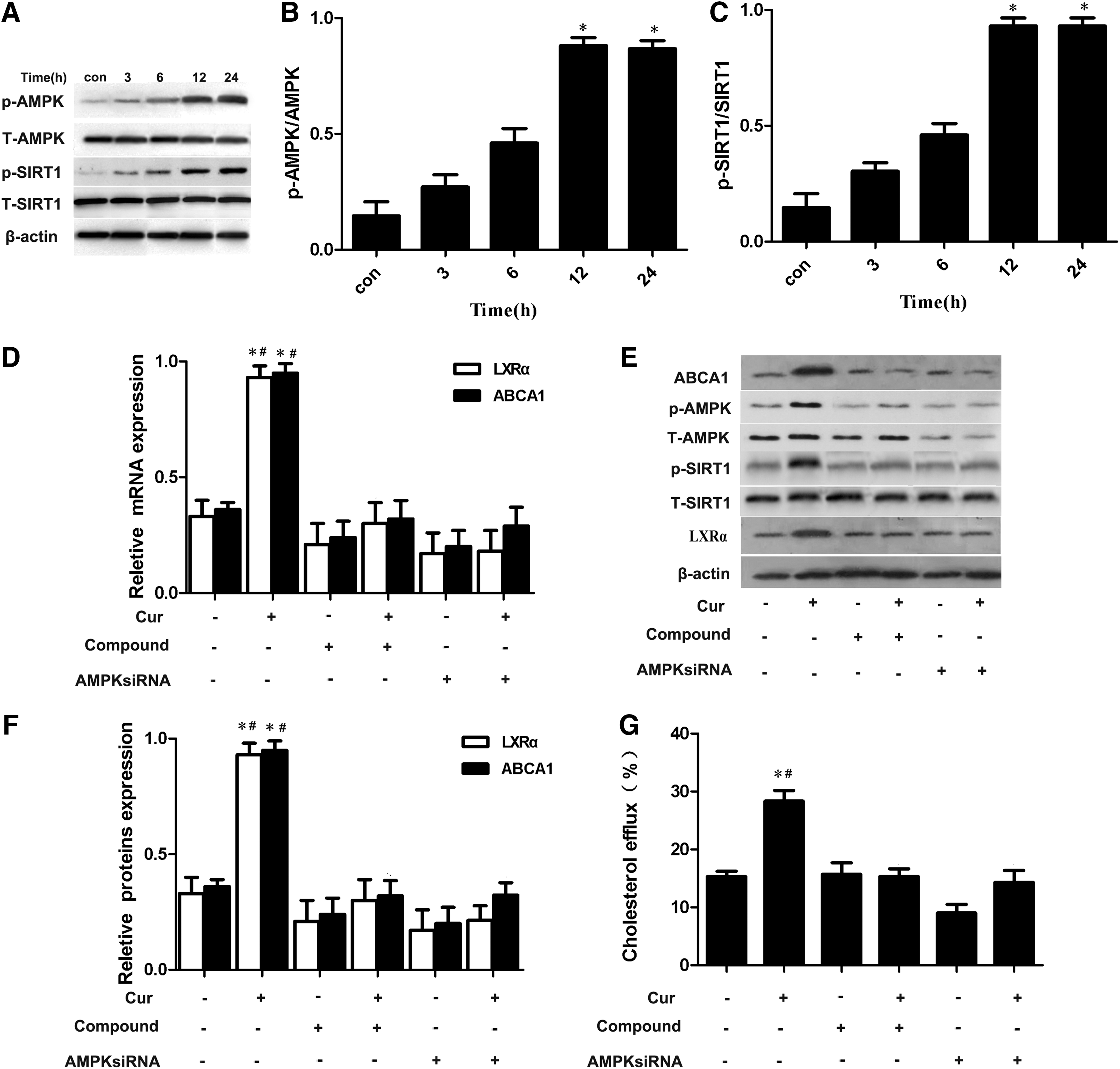
Curcumin-downregulated ABCA1 expression depended on AMP-activated protein kinase (AMPK) activation.
We also treated THP-1 macrophage-derived foam cells with specific inhibitors of Sirt1 to further investigate whether Sirt1α activation is involved in curcumin-induced ABCA1 expression. As shown in Figure 7A–C, treatment with SIRT1 inhibitor suppressed LXRα activity and ABCA1 expression compared with treatment with curcumin only. At the same time, cellular cholesterol efflux treated with SIRT1 inhibitor was significantly decreased compared with cells treated with curcumin only (Fig. 7D). Furthermore, similar results were found when a genetic approach by transient SIRT1 knockdown was used. These results demonstrate that curcumin increased ABCA1 expression through AMPK-SIRT1-LXRα.
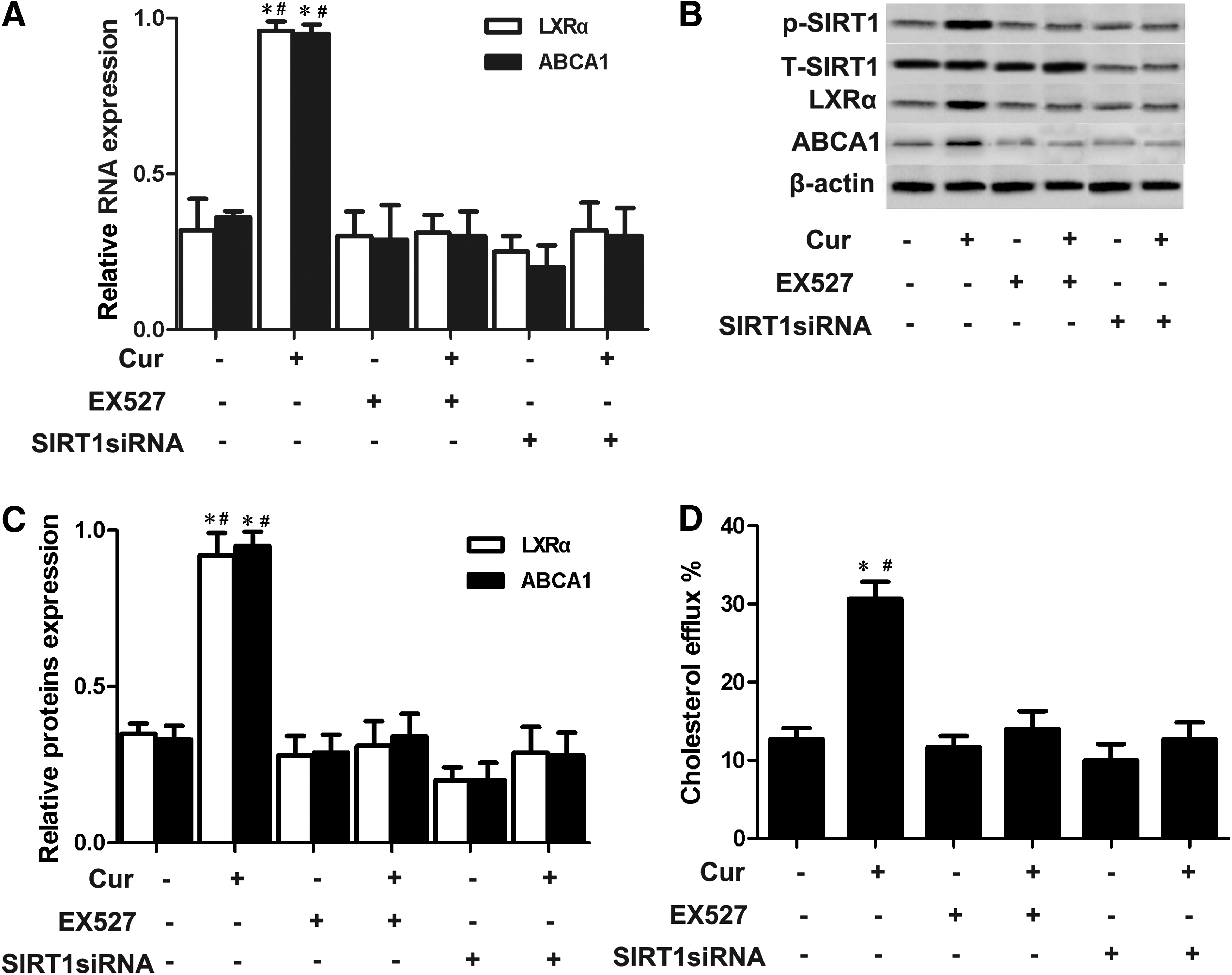
Curcumin-downregulated ABCA1 expression depended on SIRT1 activation.
Discussion
Curcumin is a polyphenolic compound with a multitude of beneficial actions for lipid metabolism and atherosclerosis (Kang et al., 2013; Liu et al., 2014). Zhao et al. (2012) reported that curcumin has suppressed foam cell formation by inhibiting SR-A-dependent ox-LDL internalization and promoting ABC-dependent cholesterol efflux. However, the efficacy and possible molecular mechanism of curcumin involved in cholesterol metabolism of macrophage-derived foam cells remained unclear. Maintenance of cellular cholesterol homeostasis is important for normal human physiology. ABCA1 mediates the secretion of cellular-free cholesterol and phospholipids to lipid-free or lipid-poor apoA to form HDL, playing a central role in maintaining cholesterol homeostasis and antiatherogenesis (Zhao et al., 2008; Liu et al., 2013). Therefore, in the current study, we focus on cholesterol efflux maintaining cholesterol homeostasis in macrophage-derived foam cells. Curcumin decreased intracellular lipid and enhanced cholesterol efflux by upregulating ABCA1 expression. These results are the same as in Zhao et al.'s (2012) study; however, the mechanisms are different. Our findings strongly suggest that curcumin has a novel effect in maintenance of cholesterol homeostasis and an improved mechanism on atherosclerosis protection. In this study, we delineated the underlying molecular mechanism.
LXRs, originally described as regulators, increase the expression of ABCA1 and RCT in ApoE−/− knockout mice (Parikh et al., 2014). LXRα activation by synthetic activator T0901317 rescued reductions in ABCA1-mRNA and increased ABCA1 expression and apoA-I-mediated cholesterol efflux in the presence and absence of atorvastatin (Zhao et al., 2008; Qiu and Hill, 2008). This finding can be supported by similar findings in glomerular mesangial cells, Caco-2, and macrophages. Moreover, our previous studies demonstrated that LXRα is activated by FGF21, which increases ABCA1 expression through the LXRα pathway (Lin et al., 2014). The presence of LXRα-binding sites in ABCA1 promoter was reported, and LXRα deficiency abolished curcumin's effect on ABCA1 expression in adipocytes, suggesting that ABCA1 is one of the target genes of LXR. Therefore, we tested the effect of curcumin on LXRα expression to identify whether the LXRα expression was induced by curcumin during the course of upregulating of ABCA1 expression. Results show that curcumin activated LXRα and significantly upregulated ABCA1 expression. In addition, LXRα expression was inhibited by LXRα siRNA, which decreased the upregulation of curcumin on ABCA1 expression. Meanwhile, cellular cholesterol efflux was completely reversed by LXRα siRNA. This result is consistent with previous findings that curcumin upregulated ABCA1 protein expression, enhanced cholesterol efflux, and led to reduced lipid accumulation in THP-1 macrophage-derived foam cells, demonstrating that LXRα was involved in ABCA1 expression by curcumin in THP-1 macrophage-derived foam cells.
The PPARγ-LXRα-ABCA1 axis is a major pathway to induce cholesterol efflux in macrophages (Chawla et al., 2001) as in adipocytes (Wu et al., 2009). One study has demonstrated the similar activating effect of curcumin on PPARγ and LXRα in the human hepatoma cell line, HepG2 (Zhai et al., 2015). However, in the present study, attention is not paid to this axis. AMPK is a major metabolic energy sensor and master regulator of metabolic homeostasis (Xu and Si, 2010). Numerous studies demonstrated that AMPK exerts protective effects on atherosclerosis, apart from the regulation of macrophage foam cell formation (Li et al., 2010). AICAR is an agonist that activates AMPK, which increases ATP-binding cassette transporters, ABCG1 and ABCA1, to promote cholesterol efflux from macrophage-derived foam cells as well as reduce plaque formation in ApoE−/− knockout mice (Wong et al., 2009). In addition, curcumin can be activated by AMPK to downregulate EMMPRIN, MMP-9, and MMP-13 expression and inflammatory factors in macrophages (Cao et al., 2014). We speculate that curcumin may activate AMPK signaling to upregulate ABCA1 expression, and our data demonstrate that curcumin activated AMPK, increased ABCA1 expression, and enhanced cholesterol efflux. THP-1 macrophage-derived foam cells incubated with AMPK inhibitor or siRNA blocked the effect of curcumin toward ABCA1 expression and cellular cholesterol efflux. This result suggests that AMPK was involved in the effect of curcumin on ABCA1 expression.
AMPK enhances NAD+-dependent type III deacetylase SIRT1 activity, resulting in modulation of the activity of downstream SIRT1 targets (Zhu et al., 2014). Our results also indicate that AMPK activation could increase SIRT1 activity. Li et al. (2007) reported that SIRT1 is a positive regulator of LXR to upregulate its activity and is important for cholesterol homeostasis. SIRT1−/− cells exhibit defective cholesterol efflux and reduce ABCA1 gene expression (Li et al., 2007). Zeng et al. (2013) reported that SIRT1 also upregulates the expression of LXR and its target genes to prevent atherosclerosis. Our results found that curcumin activated SIRT1, and SIRT1 inhibitor and siRNA-treated cells decreased the upregulation of curcumin on ABCA1 expression and LXRα activation. At the same time, cellular cholesterol efflux was also completely reversed by SIRT1 inhibitor. All these results indicate that increased ABCA1 and enhanced cholesterol efflux expression required SIRT1 activation by curcumin. FOXO3a, a transcription factor, is an important target of SIRT1. SIRT1 deacetylates FOXO3a, triggers transcription of FOXO3a-dependent stress resistance genes, and contributes to increased longevity (Das et al., 2014). However, recent research has found that curcumin reduces macrophage lipid accumulation in LDL receptor knockout mice fed a high-fat diet for 4 months through the SIRT1- FOXO3a axis. FOXO response element is the working part for FOXO3a (Mei et al., 2007). So, we supposed that FOXO3a may be involved in ABCA1 expression through SIRT1 activation. However, in the recent study, we do not investigate it, and we would be investigating it in the future.
In summary, we described the protective effects of chronic supplementation with curcumin against atherosclerosis development. Curcumin enhanced cholesterol efflux by upregulating ABCA1 expression through AMPK/SIRT1/LXRα signaling in THP-1 macrophage-derived foam cells. Our results provide further support to the beneficial effects of curcumin on attenuating the progression of atherosclerosis as well as identifying a potential novel therapeutic target for atherosclerosis prevention.
Footnotes
Acknowledgment
This study was supported by the Innovative Research Team for Science and Technology in Higher Educational Institutions of Hunan Province, Natural Science Foundation of China (No. 81070221), the Visiting Scholar Foundation of Key Laboratory for Biorheological Science, Technology (Chongqing University) of Ministry of Education (2010), the medical foundation of Hui Zhou (2014Y149), and the Medical Research Foundation of Guangdong Province (A2014810).
Disclosure Statement
No competing financial interests exist.