
Abstract
NLRP3 inflammasome, which is multiprotein complex that induces the maturity and secretion of proinflammatory interleukin-1β (IL-1β), takes a bridge between the innate and adaptive immune responses to the invading pathogens. It has been shown that porcine reproductive and respiratory syndrome virus (PRRSV) could activate the NLRP3 inflammasome but induce the host's immunosuppression. This study aims to explore whether PRRSV could encode the component to antagonize the NLRP3 inflammasome. The obtained results showed that PRRSV could induce the expression and secretion of IL-1β in early infection through the pathway of NLRP3 inflammasome in porcine alveolar macrophages (PAMs), but the levels of pro-IL-1β mRNA and IL-1β protein decreased to a degree that was similar to the level of the mock-infected group in later infection. This work also found that PRRSV nonstructural protein (nsp) 11 could inhibit the expression of pro-IL-1β mRNA induced by lipopolysaccharide (LPS) and the secretion of IL-1β induced by LPS plus nigericin in PAMs. Furthermore, the mutation studies showed that the endoribonuclease activity was essential for nsp11 to inhibit the secretion of IL-1β. Therefore, it could be indicated that PRRSV could induce the activation of NLRP3 inflammasome, but the virus encoded nsp11 to inhibit this action.
Introduction
T
Porcine reproductive and respiratory syndrome virus (PRRSV), a positive-strand RNA virus, along with equine arteritis virus, lactate dehydrogenase-elevating virus, and simian hemorrhagic fever virus, belongs to the family of Arteriviridae (Meulenberg et al., 1993). As the etiological agent of porcine reproductive and respiratory syndrome, PRRSV was first identified in Europe in 1991 and in the United States in 1992, and could cause enormous economic loss to the pig industry each year (Neumann et al., 2005). PRRSV, mainly replicated in macrophages, elicited poor innate and adaptive immune responses, and able to induce prolonged viremia, could cause persistent infections that last for months after infection (Albina et al.,1998; Van Reeth et al., 1999; Chiou et al., 2000; Lopez-Fuertes et al., 2000).
During the coevolution between the virus and the host, the host cell will immediately upregulate the antiviral molecule and establish the antiviral state to the invading virus, while the invading virus also evolves ways to effectively limit these antiviral actions. For example, virus could induce the expression of the antiviral cytokine-type I interferon and could also encode antagonists to inhibit the expression and function of this cytokine. Additionally, taking another antiviral multiprotein complex, the NLRP3 inflammasome as an example, previous studies have shown that many viruses such as influenza A virus (Thomas et al., 2009), adenovirus (Muruve et al., 2008), and sendai virus (Kanneganti et al., 2006) could activate the NLRP3 inflammasome, but meanwhile, the virus could encode proteins to inhibit the NLRP3 inflammasome. The M013 protein of myxoma virus (Rahman and McFadden, 2011), V Protein of measles virus (Komune et al., 2011), and NS1 protein of influenza A (Stasakova et al., 2005) were the proteins that could suppress the secretion of IL-1β mediated by the NLRP3 inflammasome, and both the deletion of M013 in myxoma virus and the deletion of NS1 in influenza A could lead to the enhancement of the activation of caspase-1 and the secretion level of IL-1β and IL-18. The knockout experiment also showed that Nlrp3−/− and Casp1−/− mice were more susceptible than wild-type mice after infection with a pathogenic influenza A virus, and NLRP3 played a critical role in limiting lung damage resulting from influenza A virus infection (Allen et al., 2009; Thomas et al., 2009). Therefore, the NLRP3 inflammasome will take an important role in antiviral immune response.
Together, PRRSV could activate the NLRP3 inflammasome (Bi et al., 2014), but induce the host's immunosuppression and the persistent infection (Mateu and Diaz, 2008). Therefore, it will be useful to demonstrate the mechanism of how the virus limits the antiviral response of the NLRP3 inflammasome and to understand how the PRRSV induces the immunosuppression by identifying whether PRRSV could encode the component to antagonize the NLRP3 inflammasome.
Materials and Methods
Cells and virus
MARC-145 cells, which were highly permissive for PRRSV infection, and HEK293T cells were maintained in DMEM (Gibco) with 10% heat-inactivated fetal bovine serum (FBS) (Hyclone). Primary porcine alveolar macrophages (PAMs) were obtained by postmortem lung lavage of 6-week-old pigs from PRRSV-negative pigs and cultured in RPMI-1640 medium (Gibco) supplemented with 10% heat-inactivated FBS, 100 units/mL of penicillin, and 100 μg/mL of streptomycin (Hyclone) in cell incubators at 37°C with 5% CO2. The classic type 2 (North American) PRRSV strain BJ-4 was a kind gift from Dr. Hanchun Yang (China Agricultural University).
Antibodies
For all western blot experiments, the antibodies, mouse anti-actin (Beijing Zhongshan Goldenbridge Biotechnology Company), mouse anti-FLAG (Sigma), and anti-mouse IgG HRP-linked second antibody (Abbkine) were used.
Plasmids
The plasmid pcDNA3.1-FLAG-nonstructural protein (nsp) 11 and the catalytically inactive mutant plasmids pcDNA3.1-FLAG-nsp11 H129A (nsp11 H129A) and pcDNA3.1-FLAG- nsp11 H144A (nsp11 H144A) have already been described in our previous work (Shi et al., 2011). The cDNAs encoding porcine NLRP3, ASC, procaspase-1, and pro-IL-1β were obtained by reverse transcription of total RNA from lipopolysaccharide (LPS) (Sigma)-treated PAMs, then the cDNAs were used as the template for polymerase chain reaction (PCR) by using specific primers, and finally the productions of restriction enzyme-digested PCR were inserted into the restriction enzyme digested-pcDNA3.1-FLAG to construct the expression plasmids pcDNA3.1-FLAG-NLRP3, pcDNA3.1-FLAG-ASC, pcDNA3.1-FLAG-procaspase-1, pcDNA3.1-FLAG-pro-IL-1β, and then, all newly prepared plasmids were verified by sequencing. All primers for constructing the plasmids above are listed in Table 1.
IL-1β, proinflammatory interleukin-1β; nsp, nonstructural protein; RT-PCR, reverse transcription-polymerase chain reaction.
siRNA and cell transfection
Transient transfection was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. The sequence (in Table 1) of siRNA targeting NLRP3 or ASC was designed according to Bi et al. (2014), and the siRNAs were transfected by the manufacturer's instructions.
Cell transfection, enzyme-linked immunosorbent assay, and western blot
PAMs (1 × 106 cells/well) were cultured in 24-well plates overnight, and then, the cells were transfected with the indicated plasmids and treated with LPS and/or nigericin (InvivoGen). Forty-eight hours (h) post-transfection, the supernatants were collected for subsequent assay by the enzyme-linked immunosorbent assay (ELISA). The cells were lysed by TRIzol reagent (Life Technologies) for real-time reverse transcription-polymerase chain reaction (RT-PCR).
From the report of the previous study (Komune et al., 2011), the model cells of NLRP3 inflammasome were reconstructed in the HEK293T cells. To identify whether PRRSV protein may inhibit the NLRP3 inflammasome, HEK293T cells grown to approximately 80% confluence in 24-well plates were transfected with plasmids pcDNA3.1-FLAG-NLRP3 (150 ng/well), pcDNA3.1-FLAG-ASC (50 ng/well), pcDNA3.1-FLAG-procaspase-1 (50 ng/well), pcDNA3.1-FLAG-pro-IL-1β (300 ng/well) and pcDNA3.1-FLAG-nsp11, pcDNA3.1-FLAG- nsp11 H129A, pcDNA3.1-FLAG-nsp11 H144A, or pcDNA3.1-FLAG using Lipofectamine 2000. After 24 h, the supernatants were collected and analyzed for IL-1β by ELISA and the cells were harvested and analyzed for nsp11, nsp11 H129A, nsp11 H144A proteins by western blots. Equal proteins were separated on SDS-PAGE gels and transferred to polyvinylidene difluoride membranes (Millipore Company), and then probed with appropriate antibodies. Proteins were detected using an electrochemiluminescence detection system (Cell Signaling Technology).
Measurement of the mature IL-1β protein
The cell-free supernatants were collected at the indicated time and were analyzed for IL-1β by an ELISA (R&D Systems) according to the manufacturer's instructions.
RNA extraction and real-time RT-PCR
Total RNA from PAMs was extracted by TRIzol reagent according to the manufacturer's instructions. RNA (1 μg) was reversely transcribed in a 20 μL reaction mixture (TAKARA PrimeScript™ RT Reagent Kit with gDNA Eraser). The cDNA was amplified in a 20 μL reaction mixture containing SYBR green real-time RT-PCR master mix (TAKARA) in a 7500 fast real-time RT-PCR system (Applied Biosystems). Cross-intron primers for real-time RT-PCR are listed in Table 1. The PCR conditions were as follows: initial denaturation for 10 min at 95°C, followed by 40 cycles of 3 s at 95°C, and 34 s at 60°C. A melting curve analysis was performed to ensure specificity of the products. The relative gene expression levels were normalized to the GAPDH level using the 2−ΔΔCT formula (ΔΔCT = ΔCT sample—ΔCT control).
Statistical analysis
Statistical significance was tested by one-way ANOVA followed by Tukey's post-test using GraphPad PRISM software. p-Values of less than 0.05 were considered statistically significant.
Results
PRRSV induced IL-1β in PAMs
Previous studies revealed that the infection of PRRSV induced the expression of porcine IL-1β (Qiao et al., 2011; Zhang et al., 2013) and activated the NLRP3 inflammasome (Bi et al., 2014). To investigate the dynamic changes of the expression of IL-1β during PRRSV infection, primary PAMs were infected with the PRRSV strain BJ-4 at different MOI, and then at different indicated hours of postinfection, the cells were harvested and analyzed for the levels of procaspase-1, NLRP3, ASC, and pro-IL-1β mRNA by real-time RT-PCR, and the supernatants were fetched and analyzed for the mature IL-1β by ELISA. The results showed that the levels of pro-IL-1β and procaspase-1 mRNA and the mature IL-1β were significantly increased at 24 h postinfection in a dose-dependent manner (Fig. 1A, B). This work also found that the levels of pro-IL-1β, procaspase-1 mRNA, and the mature IL-1β were significantly affected by the hours of postinfection. As shown in Figure 1C, D, PRRSV increased the levels of pro-IL-1β, procaspase-1 mRNA, and the mature IL-1β in early infection in PAMs, that is, the levels of pro-IL-1β and procaspase-1 mRNA were peaking at 24 h postinfection and the levels of the secreted IL-1β protein were peaking at 48 h postinfection. Nevertheless, 48 h after being infected with PRRSV, the levels of pro-IL-1β and procaspase-1 mRNA decreased to a degree that was similar to the level in the mock infection group. Correspondingly, the levels of the mature IL-1β in PAMs also decreased in the subsequent infection.

Porcine reproductive and respiratory syndrome virus (PRRSV) could induce the expression and secretion of inflammatory interleukin-1β (IL-1β) in early infection. Porcine alveolar macrophages (PAMs) were infected with PRRSV at different MOI (1.0, 0.1, 0.01), and then, the cells were harvested and analyzed for the levels of procaspase-1, NLRP3, ASC, and pro-IL-1β mRNA by real time reverse transcription-polymerase chain reaction (RT-PCR)
Secretion of IL-1β induced by PRRSV was dependent on the NLRP3 inflammasome
To determine whether the secretion of IL-1β induced by PRRSV was dependent on the NLRP3 inflammasome, the PAMs were infected with BJ-4 PRRSV at different MOI and then treated with glyburide (400 μM) that was a specific inhibitor to the NLRP3 inflammasome (Lamkanfi et al., 2009; Lupfer et al., 2013). As shown in Figure 2A, the glyburide could significantly inhibit the secretion of IL-1β induced by PRRSV.
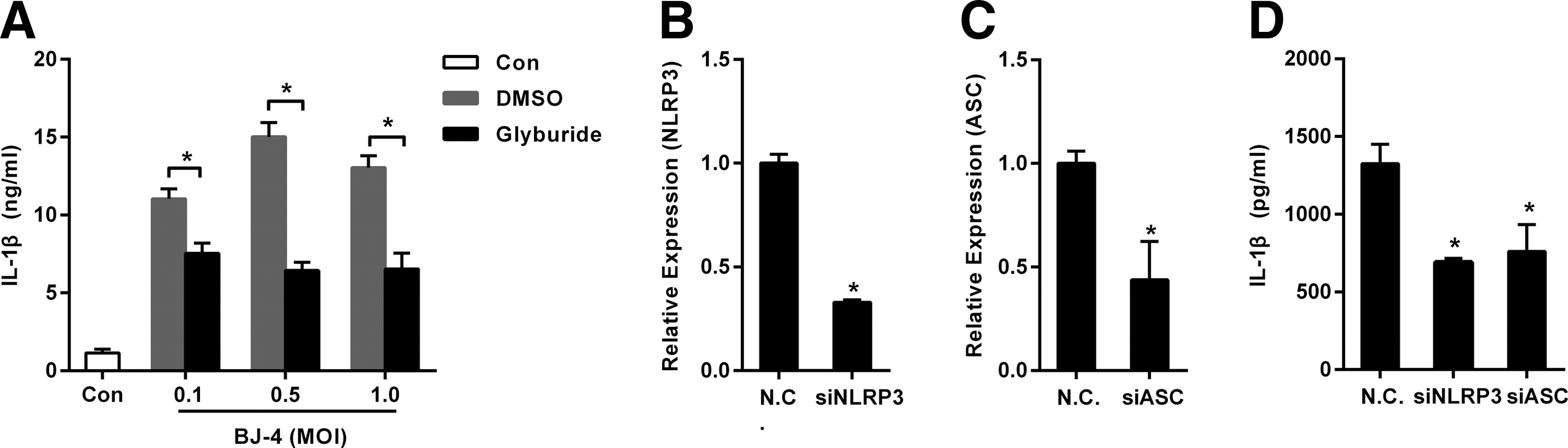
Secretion of IL-1β induced by PRRSV was dependent on the NLRP3 inflammasome.
For confirming the aforementioned results, specific siRNAs (Bi et al., 2014) targeting NLRP3 and ASC were in place of glyburide. As shown in Figure 2B–D, the specific siRNAs could downregulate the expression of NLRP3 and ASC mRNA respectively, and the specific siRNAs could significantly inhibit the secretion of IL-1β induced by PRRSV.
Endoribonuclease activity was essential for nsp11 to inhibit the expression of pro-IL-1β mRNA
The aforementioned results showed that PRRSV could induce the secretion of IL-1β at early infection but could not in later infection. It is attractive to detect which component of the invading PRRSV inhibited the secretion of IL-1β. PAMs were transfected with pcDNA3.1-FLAG (Vector) or pcDNA3.1-FLAG-nsp11 (nsp11). Six hours later, the cells were stimulated with LPS (1 μg/mL), and 42 h later, the cells were harvested and analyzed by real-time RT-PCR. The results in Figure 3A showed that nsp11 could inhibit the expression of pro-IL-1β mRNA induced by LPS.
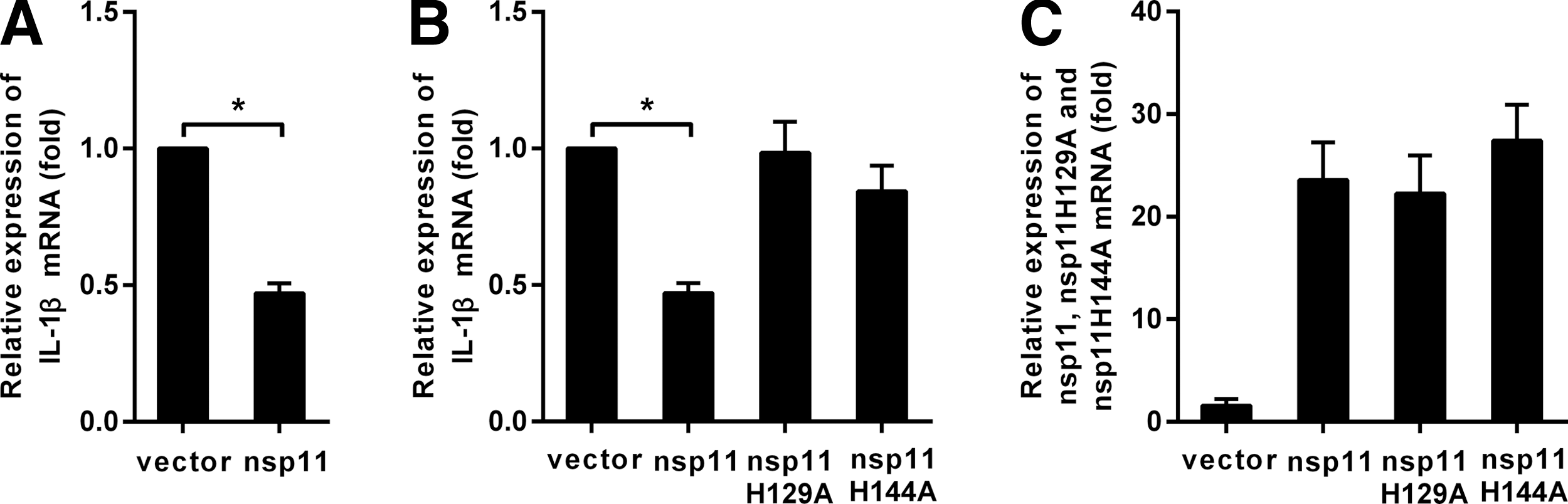
Endoribonuclease activity is essential for nonstructural protein (nsp) 11 to inhibit the expression of pro-IL-1β mRNA.
PRRSV nsp11 was an endoribonuclease (NendoU) protein, His-129, His-144, and Lys-173 were the catalytic centers, and mutating one of the three amino acids could abolish its enzyme activity (Nedialkova et al., 2009) [replacing His-129 and His-144 with Ala (nsp11 H129A, nsp11 H144A)]. PAMs were transfected with pcDNA3.1-FLAG-nsp11, pcDNA3.1-FLAG- nsp11 H129A, pcDNA3.1-FLAG-nsp11 H144A, or pcDNA3.1-FLAG, and then, the cells were stimulated with LPS, and 42 h later, the cells were harvested to detect the mRNA of pro-IL-1β. As shown in Figure 3B, nsp11 H129A and nsp11 H144A failed to block the expression of pro-IL-1β mRNA induced by LPS.
Endoribonuclease activity was essential for nsp11 to inhibit the secretion of IL-1β
Nigericin, a pore-forming toxin that has been shown to promote P2X7 receptor-independent potassium efflux treatment, could activate the NLRP3 inflammasome in LPS-primed dendritic cells (Elinav et al., 2011).
PAMs were transfected with plasmids pcDNA3.1-FLAG-nsp11, pcDNA3.1-FLAG- nsp11 H129A, pcDNA3.1-FLAG-nsp11 H144A, or pcDNA3.1-FLAG, and then, the cells were stimulated with LPS plus nigericin. As shown in Figure 4A, B, nsp11 could inhibit the production of mature IL-1β protein induced by LPS plus nigericin, whereas nsp11 H129A and nsp11 H144A could not.
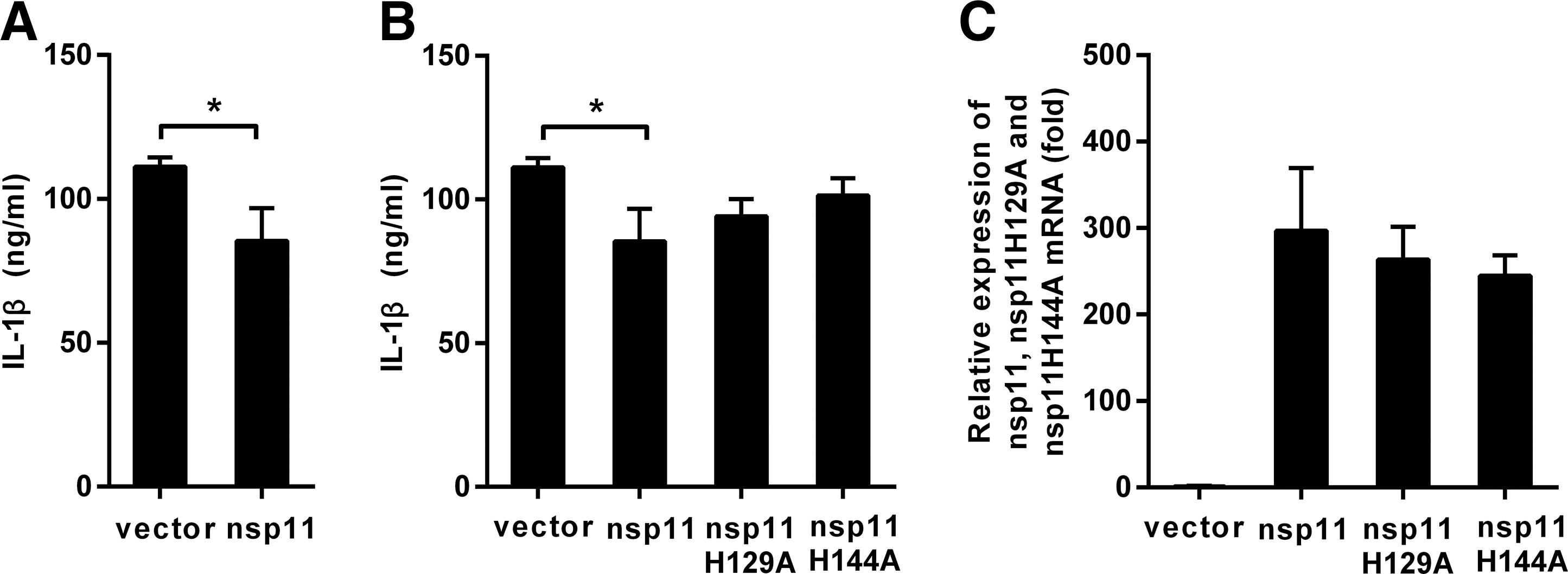
Endoribonuclease activity is essential for nsp11 to inhibit the secretion of IL-1β.
To confirm the inhibitory function of nsp11 to the NLRP3 inflammasome in PAMs, NLRP3 inflammasome was successfully reconstructed using HEK293T cells, which were deficient in the endogenous inflammasome (Bryan et al., 2009). The HEK293T cells were transiently transfected with plasmids encoding components of the NLRP3 inflammasome-NLRP3, ASC, procaspase-1, pro-IL-1β, and the increasing amounts of expression plasmid pcDNA3.1-FLAG-nsp11 or the control plasmid pcDNA3.1-FLAG. And 24 h later, the cell-free supernatants were collected and analyzed for IL-1β by ELISA. The results in Figure 5A showed that reconstitution of the NLRP3 inflammasome led to the constitutive secretion of IL-1β (Chuang et al., 2011) and the dose-dependent inhibition of the secretion of IL-1β mediated by the NLRP3 inflammasome with increasing concentrations of nsp11. Finally, HEK293T cells were cotransfected with plasmids encoding components of the NLRP3 inflammasome, the expression plasmids pcDNA3.1-FLAG-nsp11, pcDNA3.1-FLAG-nsp11H129A, pcDNA3.1-FLAG-nsp11 H144A, or pcDNA3.1-FLAG. Twenty-four hours later, the cell-free supernatants were collected and analyzed for IL-1β by ELISA. As shown in Figure 5B, nsp11 significantly inhibited the secretion of IL-1β mediated by the NLRP3 inflammasome, whereas the mutants of nsp11 that inactivated its endoribonuclease activity could not inhibit the secretion of IL-1β mediated by the NLRP3 inflammasome.
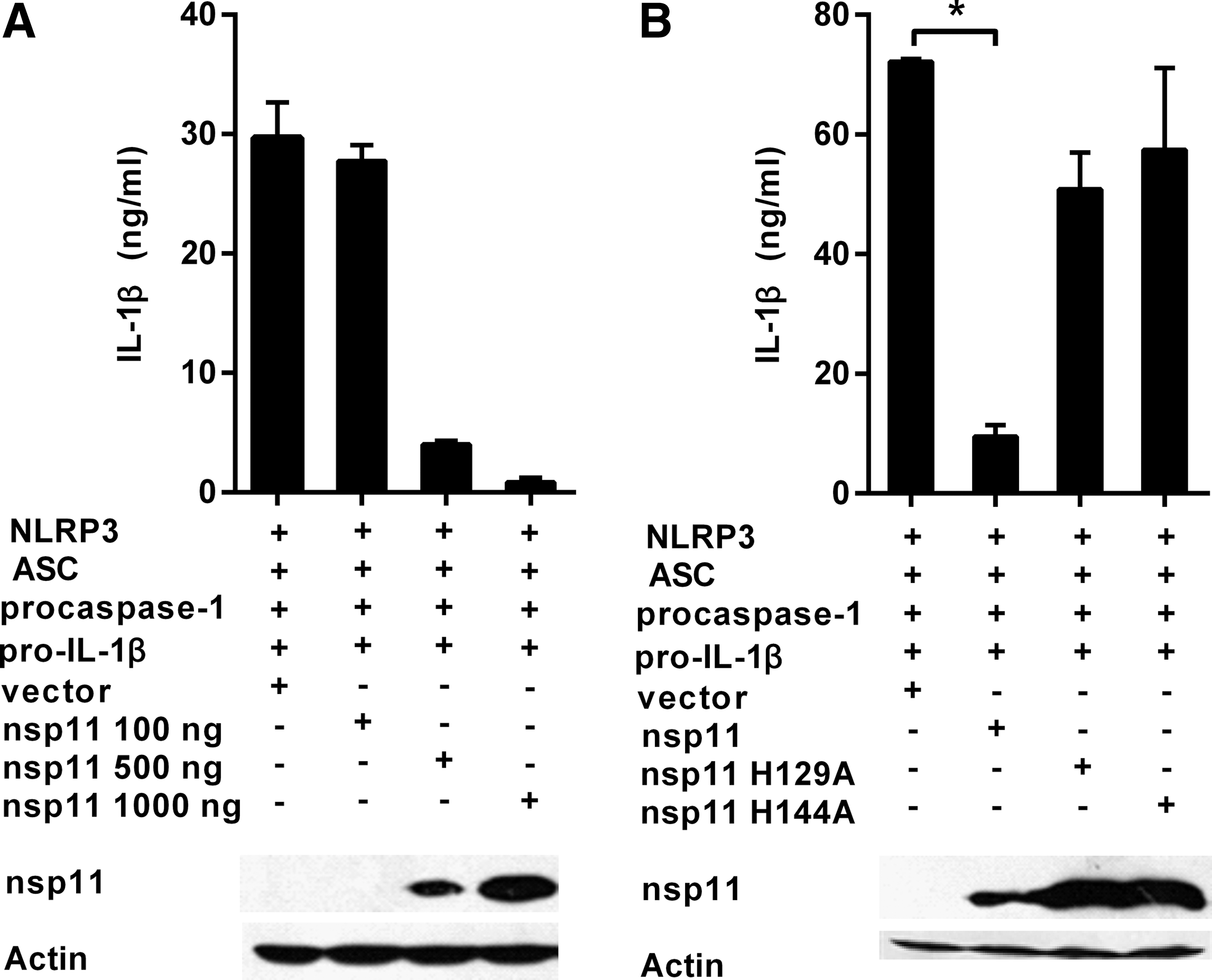
PRRSV nsp11 inhibited the secretion of IL-1β mediated by the NLRP3 inflammasome in HEK293T cells.
Discussion
The innate immune system plays an important role in confronting pathogenic and nonpathogenic insults and in instructing the development of efficient adaptive immune responses (Ciraci et al., 2012). The proinflammatory cytokine IL-1β, which can be activated by caspase-1-dependent NLRP3 inflammasome, is a critical step for an effective innate immune response (Thomas et al., 2009; Yamazaki and Ichinohe, 2014).
This work showed that PRRSV-infected PAMs secreted IL-1β through the activation of the NLRP3 inflammasome, which was consistent with the results in the previous studies (Lunney et al., 2010); however, an interesting result could be found in this work (Fig. 1), that is, the levels of pro-IL-1β and procaspase-1 mRNA and the mature IL-1β protein in later infection with PRRSV were lower than that in early infection with PRRSV in PAMs. The aforementioned results indicated that PRRSV could activate NLRP3 inflammasome, while simultaneously the virus may limit this antiviral response through a certain mechanism. Therefore, this work further explored the mechanism of how PRRSV inhibited the NLRP3 inflammasome activation and found another promising result that PRRSV encoded nsp11 to inhibit the NLRP3 inflammasome activation (Figs. 3 –5). IL-1β plays an important role in both the innate and adaptive immune response, and also in the pathological mechanism, therefore, it is reasonable that viruses antagonize the production of IL-1β by a certain mechanism. For example, some viruses encoded proteins that inhibit the production of IL-1β, such as the influenza virus NS-1 protein, V protein of MV, and so on (Stasakova et al., 2005; Komune et al., 2011).
Our and other previous studies have shown that nsp11 was an IFN antagonist and this inhibitory action was dependent on its endoribonuclease activity (Shi et al., 2011), while the similar results were also obtained in this study that nsp11 was an antagonist to the production of IL-1β and this inhibitory action was also dependent on its endoribonuclease activity, which indicated that nsp11 may take an important role in PRRSV immunosuppression, and it may be logical to take the nsp11 as a target for the development of the vaccines against PRRSV and the methods for controlling PRRSV.
In conclusion, this work suggested that PRRSV-infected PAMs secreted IL-1β through the activation of the NLRP3 inflammasome, but the virus also encoded its antagonist nsp11 to prevent the production of IL-1β, which indicated that PRRSV had evolved ways to limit this antiviral response, so this work was useful to understand how the PRRSV induced the immunosuppression.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant no. 31302073), a 973 Program (No.2014CB542700), the key project of National Natural Science Fund (No.31490600), the Doctoral Starting Up Foundation of Henan Normal University (5101049170153), and another National Foundation of China (grant no. 31472177).
Disclosure Statement
No competing financial interests exist.