
Abstract
Despite the well-documented therapeutic effects of histone deacetylase inhibitor (HDACi) on various diseases, including arthritis and asthma, the therapeutic effect of HDACi on allergic rhinitis remains unmentioned in the literature. This study investigated the therapeutic effect of sodium butyrate (SoB), a form of HDACi, on mice with allergic rhinitis. The results showed that the expression levels of histone deacetylase 1 (HDAC1), histone deacetylase 3 (HDAC3), and thymic stromal lymphopoietin (TSLP) were significantly upregulated in mice with allergic rhinitis, whereas H3 acetylation at lysine 9 (H3AcK9) was decreased. The intranasal application of SoB inhibited the expression levels of TSLP levels and upregulated the expression of H3AcK9 in a mouse model of allergic rhinitis. Furthermore, SoB treatment significantly decreased the increased levels of ovalbumin-specific IgE and improved clinical symptoms and nasal mucosa epithelial morphology in the mouse model of allergic rhinitis. In addition, we further demonstrated that SoB treatment significantly increased the serum levels of IL-2 and IFN-γ and decreased the serum levels of IL-4 and IL-10, correcting the Th1/Th2 imbalance in the mouse model of allergic rhinitis. Taken together, our study suggests that SoB has the potential to treat allergic rhinitis.
Introduction
H
The HDAC family comprises 11 members that play critical roles in development and tissue homeostasis (Delcuve et al., 2012). Of these HDACs, HDAC1 is highly expressed in inflammation-related diseases such as arthritis (Cantley et al., 2015). It was reported that HDAC3 could be induced by antigen stimulation and plays an important role in regulating allergic inflammation both in vivo and in vitro (Kim et al., 2012). Increased expression of H3 acetylation at lysine 9 (H3AcK9) was associated with inflammation inhibition mediated by zinc (Li et al., 2015). It has been reported that HDACi can notably alleviate airway hyperresponsiveness and reduce the number of bronchial eosinophils, lymphocytes, and mucosal infiltration of inflammatory cells (Ito et al., 2002; Bhavsar et al., 2008; Royce and Karagiannis, 2014). On the other hand, HDACi can attenuate the maturation of dendritic cells (DCs) and regulate the activation of T cells (Kelly-Sell et al., 2012). Thymic stromal lymphopoietin (TSLP) is a key factor involved in mediating the regulatory effect of epithelial cells on DCs in respiratory tract mucosa (Soumelis et al., 2002). TSLP enhances the ability of antigen presenting of DCs, and at the same time, TSLP-activated DCs induce an inflammatory T helper type 2 cell response through OX40 ligand (Ito et al., 2005).
Although increasing evidence has suggested that HDACi has anti-inflammatory function, the effect of HDACi on allergic rhinitis has not been well studied. In this study, we examined the therapeutic effect of sodium butyrate (SoB), a form of HDACi, on nasal symptoms in ovalbumin (OVA)-sensitized mice and investigated the potential mechanism.
Materials and Methods
Chemicals
Ovalbumin, sodium butyrate, and polyclonal rabbit anti-HDAC1 were all purchased from Sigma. Polyclonal rabbit anti-HDAC3 and monoclonal rabbit anti-H3AcK9 were purchased from Cell Signaling Technology, Inc. Other chemicals were all purchased from Sigma unless otherwise specified. Polyclonal rabbit anti-TSLP and anti-GAPDH were purchased from Santa Cruz Biotechnology, Inc.
Animals and trials
Four-week-old female BALB/c mice weighing 10–15 g, involved in this study, were purchased from Shanghai Laboratory Animal Center (SLAC). Mice were raised in pathogen-free environments under a 12-h light/12-h dark cycle at 22°C with free access to food and water and handled in accordance with the Institutional Animal Care and Use Committee of Fourth Military Medical University. The mice were randomly divided into three groups (n = 10 per group), including the control group, the OVA group, and the OVA + SoB group. The last two groups were injected intraperitoneally with mixed OVA solution (20 μg) and alum (2 mg) in a phosphate-buffered solution (PBS, 0.01%, 0.5 mL) on days 1, 3, 6 and 8, while the control group was injected with alum (2 mg) in PBS (0.01%, 0.5 mL) on the same schedule. Next, the mice of the OVA group were treated with OVA challenge (10%, 20 μL) through intranasal instillation once daily from day 15 to day 24, and the OVA + SoB group was given the same OVA challenge 1 h after being given the SoB (100 mg/mL, 20 μL) through intranasal instillation. The control group was treated with PBS (0.01%, 20 μL) through the nasal cavities once daily during the same period. The dose of SoB was chosen on the basis of our previous study (Wang et al., 2013).
Serological tests
Blood samples were acquired by using cardiac puncture under ether anesthesia after the behavior trial. Serum was obtained by centrifugation (8000 g, 4°C) for 15 min and then kept at −80°C until the tests. The OVA-sIgE Elisa Kit (HCB) was used to detect the OVA-specific IgE according to the manufacturer's instructions. The level of each OVA-sIgE was calculated from the equation of standard curve.
Evaluation of cytokines
Serum levels of IL-2, IFN-γ, IL-4, and IL-10 were measured by using ELISA kits (R&D Systems) according to the manufacturer's instructions.
Histological assay
All mice were sacrificed, and nasal mucosa tissue samples were fixed with 10% formaldehyde, embedded in paraffin, and then cut into 5-μm sections. The tissue sections of each group were dried at room temperature overnight and stained with Alcian Blue and Periodic acid–Schiff reagent (AB-PAS) for counting the number of epithelial goblet cells. Meanwhile, some samples were stained with Giemsa to allow for counting the number of eosinophil granulocytes. Sections were coded and randomly analyzed by two blinded researchers with an electron microscope (Olympus).
Western blot analysis
Proteins were obtained from the nasal mucosa tissues of each mouse 6 h after the final intranasal provocation by using lysis buffer. Proteins were extracted using a protein extraction kit (Applygen Technologies), and the concentrations in different samples were measured by the Bio-Rad protein assay kit (Bio-Rad). Equivalent amounts of proteins from each sample were isolated by 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then electrophoretically transferred to a nitrocellulose membrane (Bio-Rad). The membranes were blocked by 2.5% nonfat milk at 37°C for 1 h and then incubated with anti-HDAC1, anti-HDAC3, anti-TSLP, anti-H3AcK9, and anti-GAPDH antibodies overnight at 4°C. After exposure to horseradish peroxidase-conjugated secondary antibodies for 1 h at room temperature, the blots were visualized by using the enhanced chemiluminescence method.
Statistical analysis
Results are expressed as mean ± standard deviation. The significance of differences was evaluated by one-way analysis of variance using scientific statistic software SPSS, version 11.5 (SPSS, Inc.). A p value <0.05 was considered statistically significant.
Results
Effects of OVA challenge on HDAC1, HDAC3, H3AcK9, and TSLP in nasal mucosa
To examine histone acetylation alteration in allergic rhinitis, we detected the expression patterns of HDACs in OVA-challenged mice. As shown in Figure 1A, the HDAC1 and HDAC3 of nasal mucosa were significantly upregulated in the OVA-challenged mice compared with that of control group (p < 0.05) (Fig. 1D). Meanwhile, the expression of H3AcK9 in nasal mucosa of OVA-challenged mice decreased significantly (p < 0.01) compared with the control group (Fig. 1B, E), suggesting a decrease of the histone acetylation level. Moreover, the expression of TSLP in OVA-sensitized mice was increased compared with that in the control group and this reached remarkable significance (p < 0.01), as shown in Figure 1C and F.
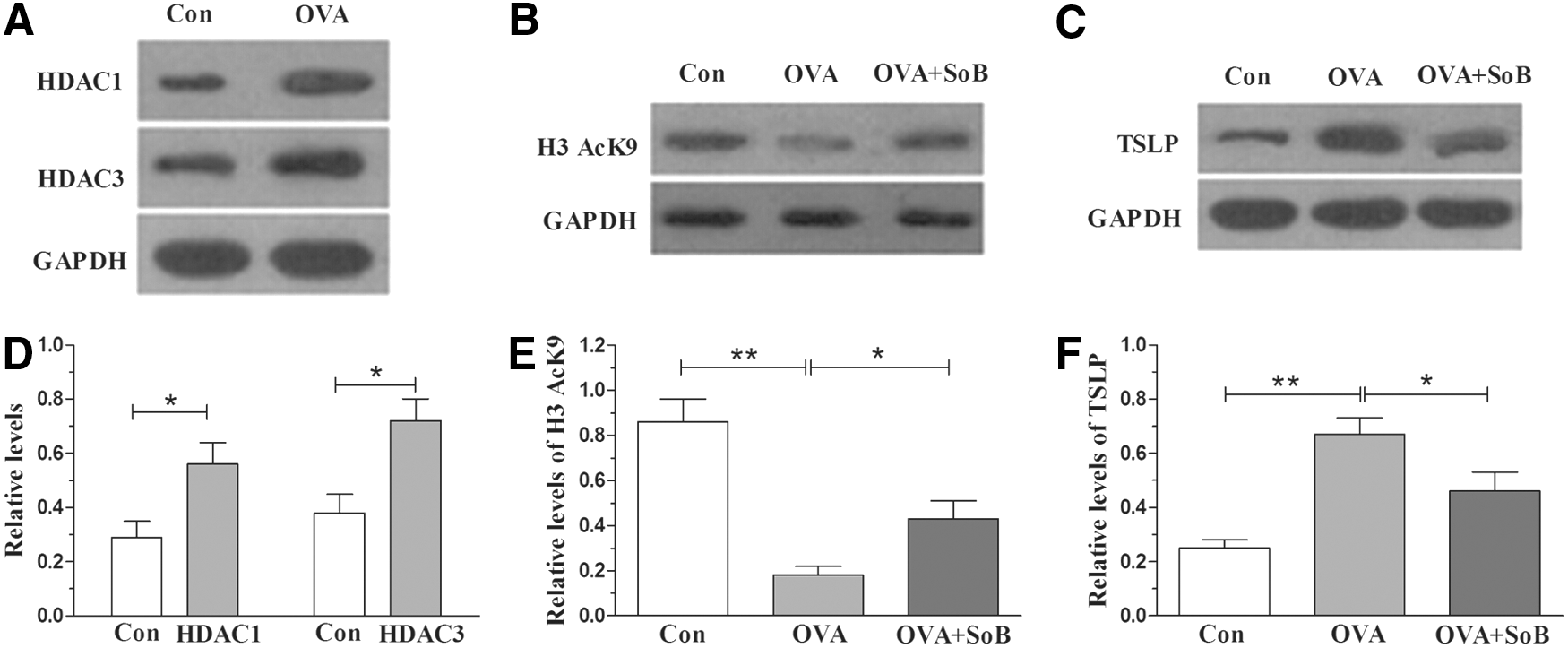
The protein levels of HDAC1, HDAC3, H3AcK9, and thymic stromal lymphopoietin (TSLP) in mouse nasal mucosa detected by Western blot analysis.
Effects of SoB administration on H3AcK9, TSLP, OVA-specific IgE, and behavior scores
SoB treatment significantly increased the level of H3AcK9 (p < 0.05) (Fig. 1B, E) and decreased the level of TSLP (p < 0.05) (Fig. 1C, F) compared with the OVA group, revealing the vital roles of SoB on the regulation of histone acetylation. To investigate the effect of SoB on the mouse model of allergic rhinitis, we detected the expression levels of OVA-specific IgE. There was significant difference in the OVA-specific IgE between the OVA group and the OVA + SoB group (p < 0.01), in which SoB treatment significantly decreased the increased levels of OVA-specific IgE (Fig. 2A). In addition, the behavior scores of the OVA + SoB group were also lower than those of the OVA group (p < 0.01, Fig. 2B), displaying the therapeutic effect of SoB on mice with OVA-induced allergic rhinitis.
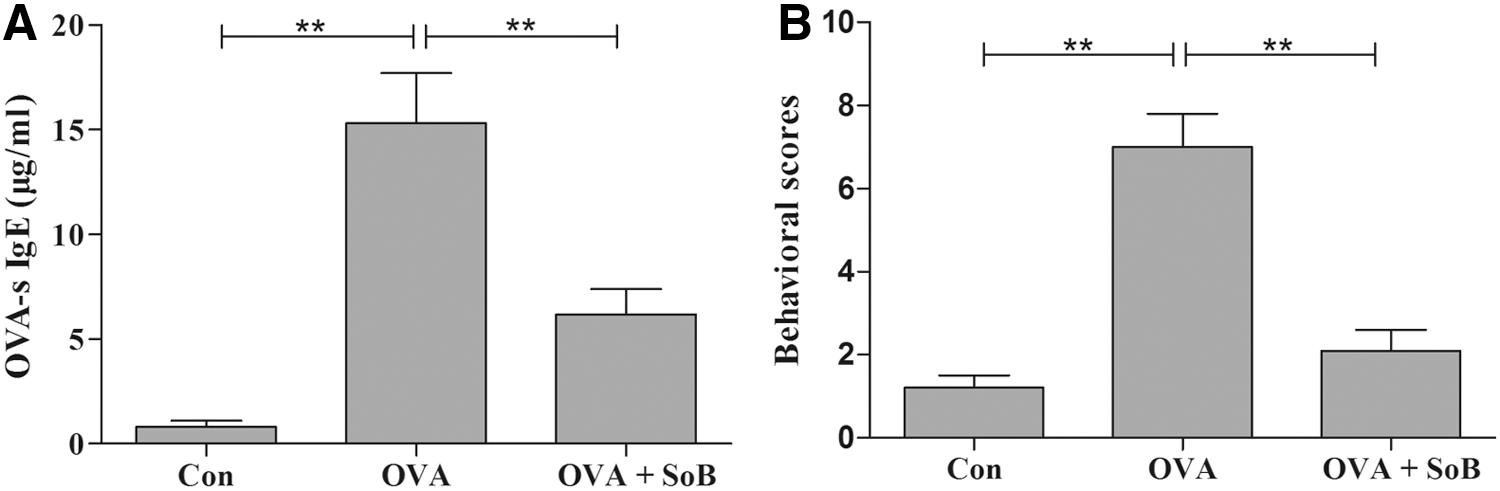
Effects of SoB on OVA-sIgE and behavior scores.
Comparison of histopathological changes of nasal mucosa
To further confirm the therapeutic effect of SoB on allergic rhinitis, we investigated the alteration of goblet cells and eosinophils in nasal mucosa tissues. The results showed that the healthy mice in the control group showed normal nasal mucosa epithelial morphology without goblet cells (Fig. 3A). In the OVA group, abundant goblet cells (Fig. 3B) in nasal mucosa tissues of mice with allergic rhinitis were observed. Interestingly, treatment with SoB apparently decreased the number of goblet cells and reversed the abnormal nasal mucosa epithelial morphology (Fig. 3C). Similarly, the amount of eosinophilic infiltration was also increased in OVA groups compared with the control group, which was apparently attenuated by SoB treatment (Fig. 3D–F).
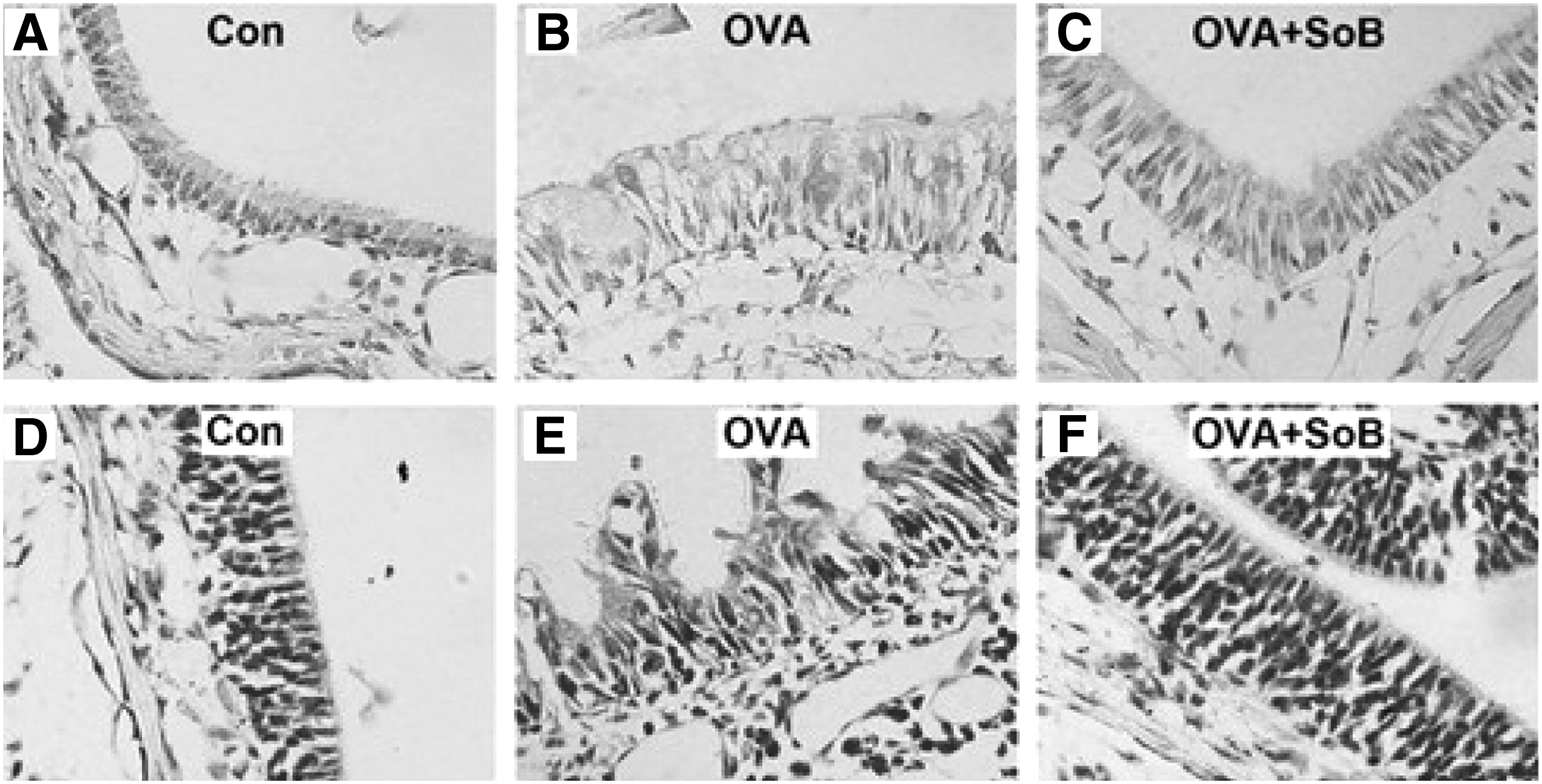
Histological changes in nasal mucosa with AB-PAS or Giemsa staining.
Effects of SoB administration on TH1/TH2 cytokines
Previous study has demonstrated that the ratio of TH1 to TH2 cells is unbalanced, favoring TH2 in allergic rhinitis (Deng et al., 2015). To investigate whether SoB has an immunoregulatory role on balancing the ratio of TH1/TH2 in the mouse model of allergic rhinitis, we detected the serum levels of TH1/TH2 cytokines in each group. The results showed that TH1 cytokines, including IL-2 (Fig. 4A) and IFN-γ (Fig. 4B), were significantly lower in the OVA group than those of the control group, whereas TH2 cytokines, including IL-4 (Fig. 4C) and IL-10 (Fig. 4D), were significantly higher in the OVA group than those of the control group. Notably, treatment with SoB markedly increased the serum levels of IL-2 (Fig. 4A) and IFN-γ (Fig. 4B) and decreased the serum levels of IL-4 (Fig. 4C) and IL-10 (Fig. 4D) compared with the OVA group. These data indicate that SoB appears to correct the TH1/TH2 balance to normal physiological levels.
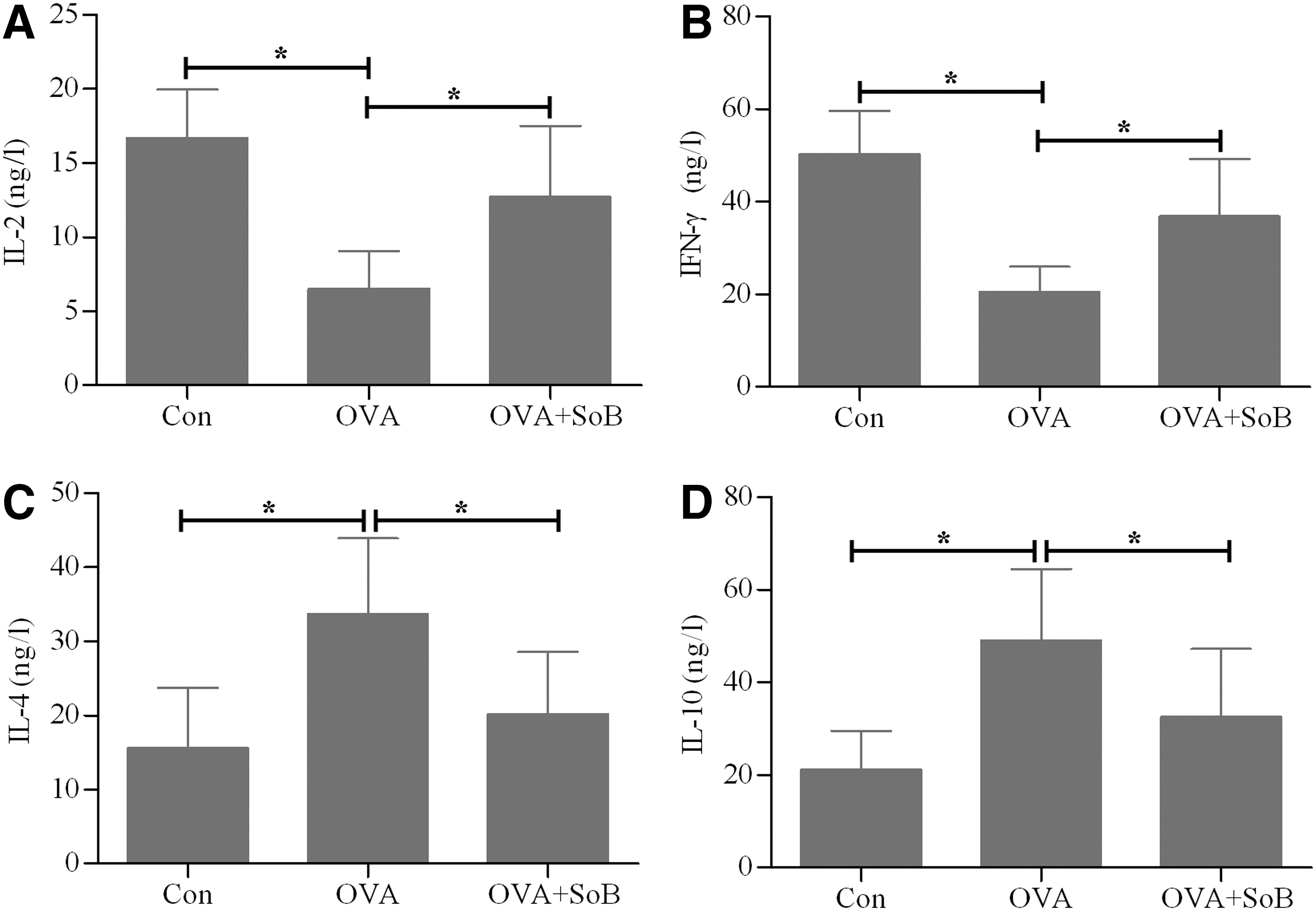
Effects of SoB administration on TH1/TH2 cytokines. The serum concentrations of
Discussion
Allergic rhinitis is defined as inflammation of the nasal mucosa, characterized by local infiltration of eosinophils, increased IgE, and major symptoms, including nasal rubbing, nasal blockage, sneezing, rhinorrhea, and lacrimation (Shinmei et al., 2009; Zheng et al., 2011; Jung et al., 2013). The main steps for managing allergic rhinitis involve avoiding various trigger factors and administering symptomatic treatment with drugs (Jung et al., 2013). Up until now, the two major drug classes recommended for the treatment of allergic rhinitis have been intranasal steroids and antihistamines, which have transient curative effects and unwanted side effects (Hiromura et al., 2007). Accordingly, other therapeutic strategies are required for patients suffering from allergic rhinitis (Fukui et al., 2009). Kuyama et al. (2010) reported that 5-aminosalicylate had a noteworthy effect on allergic rhinitis in mice. Human chemokine-like factor 1 was also found to be effective against allergic rhinitis when used as an intranasal treatment (Zheng et al., 2011). Even so, novel therapeutic strategies against allergic rhinitis are still warranted.
The role of histone acetylation in asthma has been reported. In addition, histone acetylation as a potential therapeutic target in asthma and allergies has aroused considerable interest (Ito et al., 2002; Bhavsar et al., 2008; Royce and Karagiannis, 2014). On the other hand, allergic rhinitis and asthma are manifestations of the same disease entity (Linneberg et al., 2002; Leynaert et al., 2004), such as an infiltration of eosinophils and an elevated number of activated Th2 cells (Ito et al., 2006; Bhavsar et al., 2008). Therefore, for the first time, we demonstrated the therapeutic effects of HDACi on allergic rhinitis using SoB.
The importance of TSLP in the development and progression of allergic rhinitis is becoming more appreciated (Ziegler, 2010). The mRNA and protein levels of TSLP are higher in the nasal mucosa of allergic rhinitis patients compared with healthy controls, indicating the underlying role of TSLP in the pathogenesis of allergic rhinitis (Mou et al., 2009). Similarly, high levels of TSLP expression were detected in the nasal epithelium of mice with allergic rhinitis. Furthermore, the mast cells were shown to regulate the expression of TSLP in allergic rhinitis, which suggests that mast cells play a vital role in the TSLP levels of the nasal mucosa and subsequent development of allergic rhinitis (Miyata et al., 2008).
In the present study, a nasal challenge with OVA led to increased HDAC1 and HDAC3, which may be involved in the pathogenesis of allergic rhinitis. HDAC1 and HDAC3 have been suggested to be extensively involved in allergic inflammation-related diseases (Kim et al., 2012; Cantley et al., 2015). The levels of TSLP through an OVA challenge had a trend of high expression accompanied by HDAC1 and HDAC3. Consequently, we deduced that HDAC1 and HDAC3, as upstream regulatory molecules of TSLP, may have modulatory roles in allergic rhinitis based on the fact that TSLP plays a critical role in initiating allergic immune responses (Soumelis et al., 2002; Wang et al., 2007; Siracusa et al., 2013; Yao et al., 2013), although there is a paucity of evidence to support this interesting view. Furthermore, it is noteworthy that the HDAC inhibitor, SoB, suppressed the expression of TSLP, leading to improvement of clinical symptoms in allergic rhinitis.
Interestingly, we found that SoB played an important role in maintaining the immunologic balance of TH1/TH2 cells in the mouse model of allergic rhinitis. Previous studies have suggested that butyrate is the end product of gut microbiota anaerobic fermentation of dietary fiber that plays critical roles in the development of the host immune system (Round and Mazmanian, 2009; Cerf-Bensussan and Gaboriau-Routhiau, 2010). Butyrate secreted by bacteria from the phyla, Bacteroidetes and Firmicutes, can be found at high concentrations in the large intestine (Louis and Flint, 2009). Intriguingly, patients with inflammatory disease such as inflammatory bowel disease have reduced numbers of bacteria that produce butyrate (Sokol et al., 2009). Abrahamsson et al. (2012) reported that low intestinal microbial diversity during the first month of life was associated with subsequent atopic eczema. Atopic dermatitis patients show an enrichment of a subspecies of the major gut species, Faecalibacterium prausnitzii, and have a decreased level of butyrate in the fecal samples (Song et al., 2015). Thus, gut microbiota diversity has been suggested to play an important role in atopic disease (Azad et al., 2013; Lu and Ni, 2015). Considering that gut microbiota are the main source of butyrate in vivo, gut microbiota may be used as potential targets for intervention against allergic diseases.
In addition, we found that SoB significantly upregulated the protein expression of H3AcK9, which was capable of inhibiting inflammatory processes (Li et al., 2015). Our results also showed that SoB corrected the TH1/TH2 balance to normal physiological levels in the mouse model of allergic rhinitis. In summary, the HDAC inhibitor, SoB, improved the clinical symptoms of allergic rhinitis in OVA-challenged mice. This evidence demonstrates that SoB is a potential therapeutic agent for allergic rhinitis. Despite the obvious therapeutic effect on mice, the underlying pharmacological mechanisms of SoB for the treatment of allergic rhinitis need to be further defined in future preclinical research.
Footnotes
Acknowledgment
This study was supported by the National Natural Science Foundation of China (No. 81271069).
Disclosure Statement
No conflicts of interest, financial or otherwise, are declared by the authors.