
Abstract
We have previously found that the aqueous extract of Gracilaria tenuistipitata (AEGT) and its partitioned fractions had antioxidant properties in biochemical assays. Although the butanol-partitioned fraction of AEGT (AEGT-pBuOH) had a stronger antioxidant performance than AEGT, its biological effects are still unknown. In this study, the cellular responses of oral cancer cells to AEGT-pBuOH were monitored in terms of cell viability, cell cycle progression, apoptosis, and oxidative stress responses. In an ATP content assay, the cell viability of oral cancer cells treated with AEGT-pBuOH was dose responsively inhibited (p < 0.005). For flow cytometry, AEGT-pBuOH was also found to dose responsively induce cell cycle disturbance by propidium iodide (PI) staining and to induce apoptosis by annexin V/PI and pan-caspase staining (p < 0.005). In AEGT-pBuOH-treated oral cancer cells, the reactive oxygen species (ROS) was increased and mitochondrial membrane potential was decreased in a dose–response manner (p < 0.005). These results suggest that AEGT-pBuOH inhibited the proliferation and induced apoptosis of oral cancer cells involving the ROS generation and mitochondrial depolarization.
Introduction
O
This article focuses on natural products of red algae, specifically the Gracilaria species, because its abundant bioactive extracts and compounds have been well reviewed (de Almeida et al., 2011; Francavilla et al., 2013). Gracilaria tenuistipitata is distributed in the tropical and subtropical regions of the Western Pacific (Chang and Xia, 1988). It is also commonly cultured in China (Chiang and Lin, 1989) and Taiwan (Ajisaka and Chiang, 1993). Therefore, G. tenuistipitata is easy to collect for both food industry and experimental purposes.
Consistent reports have found that aqueous extract of G. tenuistipitata (AEGT) had the properties of anti-H2O2-induced DNA damage (Yang et al., 2012b), anti-HCV viral replication (Chen et al., 2013), anti-LPS-induced inflammation in vitro, and anti-CCl4-induced hepatoxicity in vivo (Tseng et al., 2014). However, these findings were based on the crude extracts of G. tenuistipitata.
More advanced separation techniques, such as liquid–liquid solvent partitioned extraction (Melwanki and Fuh, 2008; Guleriaa and Kumara, 2009), may improve the identification of several bioactive fractions of AEGT. Recently, we performed a partitioned extraction for AEGT (Yeh et al., 2015) and found that the total phenolic contents (TPC), 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,2-azinobis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging activities, and ferric reducing antioxidant power were higher in AEGT with n-butanol partition (AEGT-pBuOH) than AEGT alone. Although AEGT demonstrated antioxidant properties in a cell model (Yang et al., 2012b), the biological effects of AEGT-pBuOH remain unclear. Moreover, the redox balance of antioxidants is sometimes controversial (Wang and Yi, 2008), and the antioxidants may behave like a double-edged sword in cellular redox state (Bouayed and Bohn, 2010; Seifirad et al., 2014).
Therefore, this study explores the impact of AEGT-pBuOH on oral cancer cells in terms of cell viability, cell cycle progression, apoptosis, and oxidative stress responses.
Materials and Methods
Raw materials and preparation of AEGT-pBuOH
The preparation of AEGT and its nuclear magnetic resonance spectroscopy (NMR) spectrum characterization were described in our previous work (Yang et al., 2012b). AEGT-pBuOH was prepared from AEGT as described (Yeh et al., 2015). In brief, 10 g of the powdered AEGT were dissolved with 500 mL of distilled water and then partitioned with n-butanol three times. Subsequently, the partitioned solvent extracts of AEGT (AEGT-pBuOH) were evaporated under low pressure and stored in a dry box before use. AEGT-pBuOH was dissolved in DMSO for drug treatment, and the concentration of DMSO in all treatments was less than 1%.
Cell cultures
The human oral cancer cell lines Ca9-22 (Yen et al., 2012) and CAL 27 (Huang et al., 2014) were maintained in DMEM (Gibco, Grand Island, NY) and supplemented with 10% fetal bovine serum and penicillin/streptomycin in a humidified atmosphere containing 5% CO2 at 37°C.
Cell viability assay
The cell viability of AEGT-pBuOH was estimated using the ATPlite Luminescence ATP Detection Assay System (PerkinElmer Life Sciences, Boston, MA) according to the manufacturer's instructions (Wei et al., 2011). AEGT-pBuOH was dissolved in DMSO and diluted in the medium for cell treatment at concentrations of 0, 0.25, 0.5, 1, and 2 mg/mL for 24 h.
Cell cycle analysis
Cell cycle distribution was analyzed according to DNA staining intensity as described (Wang et al., 2010). In brief, cells were treated with 0, 0.25, 0.5, 1, and 2 mg/mL of AEGT-pBuOH for 24 h. After harvesting for 70% ethanol fixation, cells were stained with 50 μg/mL of propidium iodide (PI) (Sigma, St Louis, MO) in Phosphate-buffered saline (PBS) for 30 min at 37°C in darkness. Cell cycle phases were detected by a flow cytometry analysis (BD Accuri™ C6; Becton-Dickinson) using BD Accuri C6 software (version 1.0.264).
Apoptosis assay—annexin V/PI
Apoptosis assay was performed by annexin V (Strong Biotech Corporation, Taipei, Taiwan)/PI (Sigma) as previously described (Ahn et al., 2012; Chiu et al., 2013). Briefly, cells were treated with 0, 0.25, 0.5, 1, and 2 mg/mL of AEGT-pBuOH for 24 h. Cells were then mixed with 10 μg/mL of annexin V-fluorescein isothiocyanate and 20 μg/mL of PI for 30 min before analysis using the BD Accuri C6 flow cytometer (Becton-Dickinson) and its software.
Pan-caspase activity assay
Most caspases are known to selectively bind to peptide Val-Ala-Asp (VAD). Using the reporter fluorescent dye (TF2-VAD-FMK), the generic activation of caspases (caspases-1, -3, -4, -5, -6, -7, -8, and -9) was detected by the Generic Caspase Activity Assay Kit (Abcam, Cambridge, UK) as described previously (Yeh et al., 2012). Briefly, AEGT-pBuOH-treated Ca9-22 cells were suspended in a medium with TF2-VAD-FMK in the cell incubator for 1 h. After PBS washing, cells were resuspended in 0.5 mL of assay buffer and immediately analyzed using the BD Accuri C6 flow cytometer and its software.
Reactive oxygen species assay
Intracellular reactive oxygen species (ROS) status was determined by 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) (Sigma) as previously described (Yen et al., 2012). Briefly, AEGT-pBuOH-treated cells were incubated in the cell incubator with 10 μM of H2DCF-DA in PBS for 30 min. After PBS washing, cells were resuspended in PBS for analysis using the BD Accuri C6 flow cytometer and its software.
Mitochondrial membrane potential assay
Mitochondrial membrane potential (MMP) detection was performed using a MitoProbe™ DiOC2(3) Assay Kit (Invitrogen, San Diego, CA) as described (Yen et al., 2014). Briefly, AEGT-pBuOH-treated cells were suspended in 50 nM of DiOC2(3) in PBS for 30 min in the cell incubator. After PBS washing, cells were resuspended in PBS for analysis using the BD Accuri C6 flow cytometer and its software.
Statistical analysis
All data are presented as mean ± SD. Significance of experimental groups with controls is based on Student's t-test.
Results
Cell viability of AEGT-pBuOH-treated oral cancer cells
At treatments of various concentrations of AEGT-pBuOH for 24 h, the cell viabilities of two oral cancer cell lines (Ca9-22 and CAL 27) were significantly inhibited in a dose–response manner in terms of ATP assays (Fig. 1).

Cell viabilities of two oral cancer cell lines were inhibited by AEGT-pBuOH. Cells were incubated with AEGT-pBuOH (0, 0.25, 0.5, 1, and 2 mg/mL) for 24 h. Cell viability was measured by ATP content. Data are presented as mean ± SD (n = 3). **p < 0.005 against vehicle control (<0.1% DMSO only). AEGT-pBuOH, butanol-partitioned fraction of AEGT.
Cell cycle changes in AEGT-pBuOH-treated oral cancer Ca9-22 cells
Figure 2A shows the DNA content profiles of AEGT-pBuOH-treated oral cancer Ca9-22 cells (0, 0.25, 0.5, 1, and 2 mg/mL) for 24 h. In Figure 2B, the sub-G1 and G1 percentages of AEGT-pBuOH-treated oral cancer cells were significantly higher than in controls, and their G2/M percentages were significantly lower than those of controls.
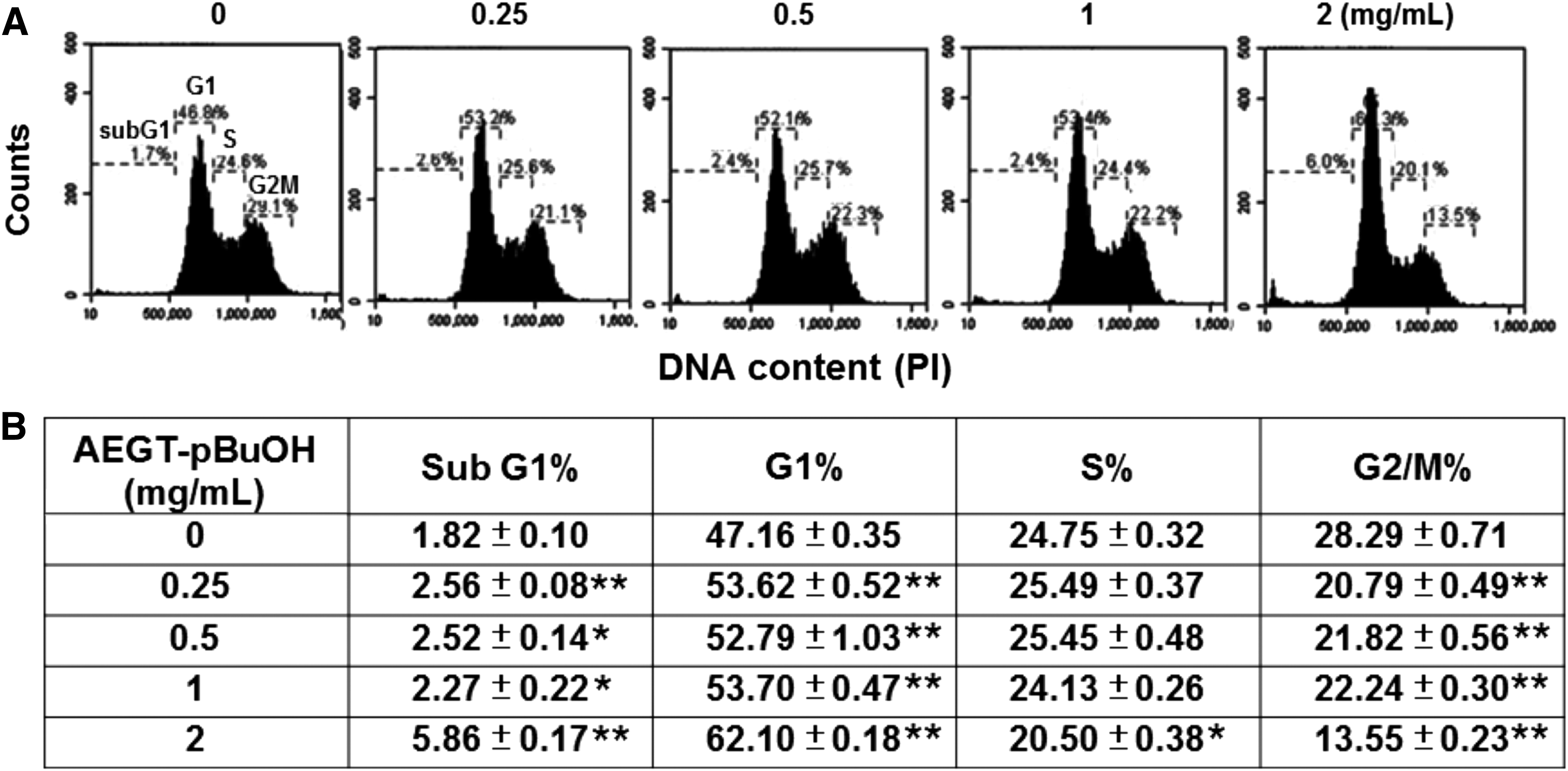
AEGT-pBuOH induced cell cycle changes in oral cancer Ca9-22 cells.
Apoptosis status of AEGT-pBuOH-treated oral cancer Ca9-22 and CAL 27 cells
Figure 3A and B, respectively, shows the apoptotic profiles of annexin V-PI patterns of the Ca9-22 and CAL 27 cells treated with AEGT-pBuOH (0, 0.25, 0.5, 1, and 2 mg/mL for 24 h). After analysis, the annexin V-positive percentages of AEGT-pBuOH-treated oral cancer Ca9-22 and CAL 27 cells were dose responsively increased (Fig. 3C, D).

AEGT-pBuOH induced apoptosis of oral cancer cells.
Activity analysis of pan-caspase in AEGT-pBuOH-treated oral cancer Ca9-22 and CAL 27 cells
In Figure 4, the activity of a set of apoptosis-related caspases in the AEGT-pBuOH-treated oral cancer Ca9-22 and CAL 27 cells was evaluated by flow cytometry (Yeh et al., 2012). The pan-caspase-positive populations were shifted to the right side when the AEGT-pBuOH concentrations were increased (Fig. 4A, B). After treatment, the pan-caspase-positive percentages of AEGT-pBuOH-treated oral cancer Ca9-22 and CAL 27 cells were dose responsively increased (Fig. 4C, D).
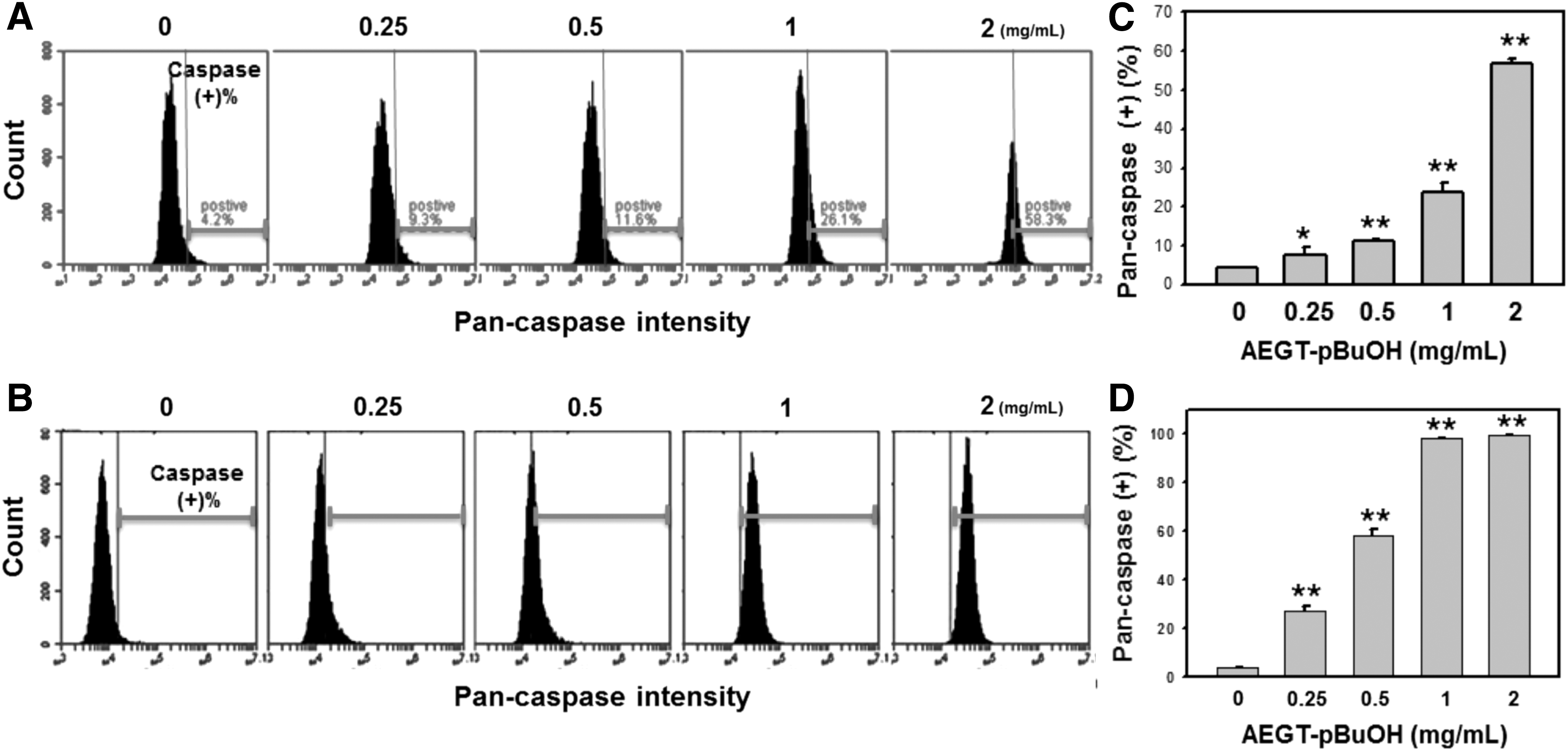
AEGT-pBuOH induced activation of generic caspase in oral cancer cells.
ROS generation of AEGT-pBuOH-treated oral cancer Ca9-22 cells
The ROS changes in the AEGT-pBuOH-treated oral cancer Ca9-22 cells were monitored using flow cytometry (Fig. 5). Figure 5A shows the profiles of ROS-positive percentages of 0, 0.25, 0.5, 1, and 2 mg/mL of AEGT-pBuOH for 24 h. The ROS-positive percentages of AEGT-pBuOH-treated oral cancer Ca9-22 cells were dose responsively increased (Fig. 5B).
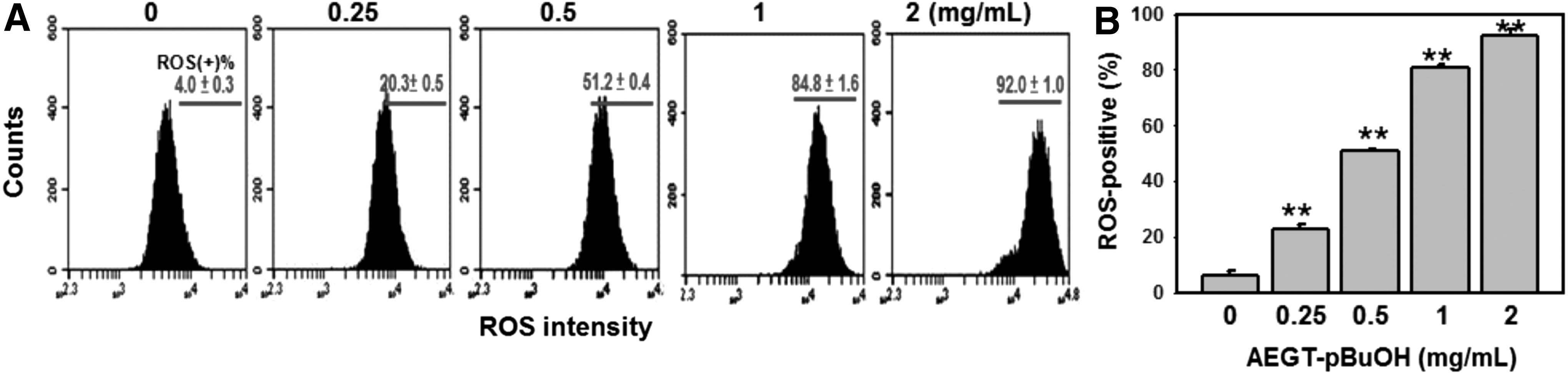
ROS changes in AEGT-pBuOH-treated oral cancer Ca9-22 cells.
MMP status of AEGT-pBuOH-treated oral cancer Ca9-22 cells
MMP changes in the AEGT-pBuOH-treated oral cancer Ca9-22 cells were monitored using flow cytometry. Figure 6A shows the profiles of MMP-negative percentages of 0, 0.25, 0.5, 1, and 2 mg/mL of AEGT-pBuOH. The MMP-negative percentages of AEGT-pBuOH-treated oral cancer Ca9-22 cells were dose responsively increased (Fig. 6B).
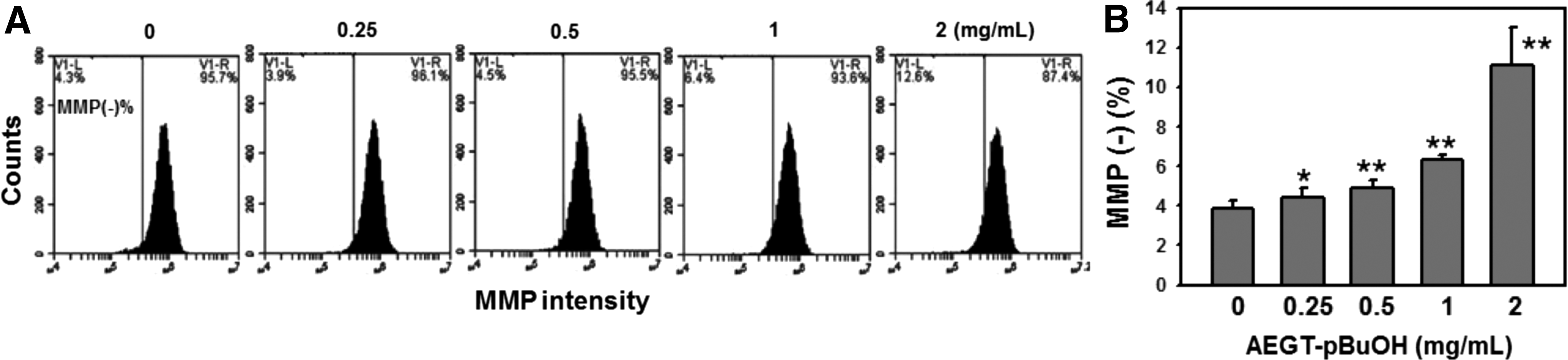
MMP changes in AEGT-pBuOH-treated oral cancer Ca9-22 cells.
Discussion
We previously reported that AEGT had antioxidant effects in biochemical assays and protective effects against H2O2-induced cellular DNA damage, where the AEGT alone was not harmful to lung cancer H1299 cells (Yang et al., 2012b). We also found that AEGT-pBuOH had a stronger antioxidant performance than AEGT in terms of biochemical assays (TPC, DPPH/ABTS scavenging activity, and reducing power) (Yeh et al., 2015). Interestingly, the current study found that AEGT-pBuOH can dose responsively reduce the cell viability of oral cancer Ca9-22 cells in terms of ATP content analysis. This result suggested that extracts with different reagents may have different natural bioactive products (Yang et al., 2012a). Moreover, it also suggested that higher concentrations of natural antioxidant products may interfere with the redox homeostasis and may be suitable for use in cancer treatment (Montero and Jassem, 2011; Glasauer and Chandel, 2014; Ribas et al., 2015). Consistent with our finding, previous studies reported that the roles of antioxidants may depend on its concentration. For example, beneficial effects may appear at physiologic concentrations of antioxidants, but deleterious effects may appear at high concentrations (Bouayed and Bohn, 2010).
Many studies have reported the anticancer effects of butanol partition extracts. For example, IC50 value for butanol partition extracts from rhizomes of Agapetes megacarpa was 340 μg/mL for breast cancer MCF-7 cells after 24 h of treatment based on a sulforhodamine B assay (Jariya Alongkornsopit et al., 2014). IC50 values for butanol partition extracts of Marchantia convoluta were greater than 200 μg/mL for lung cancer H1299 and liver cancer HepG2 cells based on an MTT assay (Xiao et al., 2006). In our current study, the ATP assay-based IC50 values for AEGT-pBuOH at 24 h of treatment were 220 and 540 μg/mL for oral cancer CAL 27 and Ca9-22 cells, respectively. In contrast, AEGT alone was not harmful to the lung cancer H1299 cells (Yang et al., 2012b). Similarly, we previously reported that the IC50 value for the ethanolic extracts of G. tenuistipitata (EEGT) (Yeh et al., 2012) was 1610 μg/mL for oral cancer Ca9-22 cells after 24 h of treatment based on an MTS assay. The major different components in the two 1H NMR spectra of AEGT-pBuOH and EEGT were observed in the regions I and II (see Supplementary Fig. S1; The characterization of NMR spectrum analysis for AEGT-pBuOH and EEGT; Supplementary Data are available online at
In the current study, the antiproliferative effect of AEGT-pBuOH was coupled with cell cycle disturbance. Although the sub-G1 accumulation was weak, it still showed a significant difference. Similarly, few sub-G1 populations have been reported in (−)-anonaine-treated lung cancer cells with apoptosis at 24 and 48 h (Chen et al., 2011). A possible explanation is that DNA damage may appear predominantly in G2/M phases and the DNA contents may move to S or G1 peaks, but the DNA damage is insufficient to generate a massive sub-G1 increase when apoptosis occurs. This speculation was partly supported by changes in the cell cycle distribution (Fig. 2). Moreover, the apoptotic-like effect of the sub-G1 accumulation of AEGT-pBuOH-treated oral cancer Ca9-22 cells had been further validated by annexin V/PI (Fig. 3). Since both the extrinsic and intrinsic pathways of apoptosis were mediated by several caspases (Storey, 2008; Cha and Kim, 2012), the general caspase activities (caspases-1, -3, -4, -5, -6, -7, -8, and -9) were also examined. The AEGT-pBuOH-induced apoptosis was further validated by pan-caspase activation (Fig. 4). Many natural product experiments also demonstrate that ROS modulation is an effective strategy for cancer treatment (Chiu et al., 2009; Yen et al., 2010, 2012). Similarly, we found that AEGT-pBuOH induced ROS generation and the mitochondrial membrane depolarization of oral cancer Ca9-22 cells (Figs. 5 and 6). Therefore, these results suggested that oxidative stress may be involved in the antiproliferative and apoptotic effects of AEGT-pBuOH against oral cancer cells. This warrants further investigation of the role of oxidative stress evidenced by the N-acetylcysteine (Sunitha et al., 2013; Venkataramana et al., 2014). Moreover, oxidative stress may induce changes in endoplasmic reticulum stress (Farooqi et al., 2015) and autophagy (Farooqi et al., 2014). These changes in AEGT-pBuOH-treated oral cancer cells may be further examined in future.
Conclusions
In the current study, we found that AEGT-pBuOH can inhibit proliferation and induce apoptosis of oral cancer cells involving oxidative stress. These results suggest that AEGT-pBuOH is a potential marine natural product for anti-oral cancer treatment.
Footnotes
Acknowledgments
This work was supported by funds of the Ministry of Science and Technology (MOST 104-2320-B-037-013-MY3 and MOST103-2314-B-037-010-MY3), the National Sun Yat-sen University-KMU Joint Research Project (grant no. NSYSU-KMU 104-p036), the Kaohsiung Municipal Ta-Tung Hospital (kmtth-103-026), the Kaohsiung Medical University Hospital (KMUH103-3M39), the Health and Welfare Surcharge of Tobacco Products, the Ministry of Health and Welfare, Taiwan, Republic of China (MOHW104-TDU-B-212-124-003 and MOHW105-TDU-B-212-134005), and ChiMei-KMU Joint Project (104CM-KMU-02). We also thank Mr. Yi-An Chung for his technical support for flow cytometer.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.