
Abstract
The variation in cytokine production during bacterial invasion of human intestinal epithelial cells (IECs) is a contributing factor for progression of the infection. A few Salmonella enterica Heidelberg strains isolated from poultry products harbor transmissible plasmids (TPs), including those that encode a type-IV secretion system. Earlier, we showed that these TPs are responsible for increased virulence during infection. This study examines the potential role of these TPs in cytokine production in IECs. This study showed that S. Heidelberg strains containing TPs (we refer as virulent strains) caused decreased interleukin (IL)-10 production in IECs after 1 h infection. The virulent strains induced a high level of tumor necrosis factor-α production under identical conditions. The virulent strains of S. Heidelberg also altered the production of IL-2, IL-17, and granulocyte macrophage colony-stimulating factor compared to an avirulent strain. As a part of infection, bacteria cross the epithelial barrier and encounter intestinal macrophages. Hence, we examined the cytotoxic mechanism of strains of S. Heidelberg in macrophages. Scanning electron microscopy showed cell necrosis occurs during the early stage of infection. In conclusion, virulent S. Heidelberg strains were able to modify the host cytokine profile during the early stages of infection and also caused necrosis in macrophages.
Introduction
S
The intestinal epithelial cells (IECs) serve as a physical and biochemical barrier, which separates the host from the gastrointestinal (GI) commensal microflora (Ley et al., 2006). The human GI tract accommodates around 1014 bacterial cells. These organisms colonize and form a large bacterial community in the lumen, which is indispensable to the health of the host. The intestinal mucosal surface is the primary site for Salmonella attachment and infection initiation. During bacterial invasion, the cell surface molecules of IECs sense the pathogen and respond rapidly to the microbial stimuli by reinforcing cytokine secretion and releasing granules with antimicrobial peptides into the intestinal luminal fluid (Ayabe et al., 2000; Tanabe et al., 2005; Bevins and Salzman, 2011).
Cytokines/chemokines secreted by IECs can recruit proinflammatory cells to the site of infection (Artis and Grencis, 2008). Cytokines that are secreted from macrophages and T cells can also induce the expression of major histocompatibility (MHC I and II) molecules and receptors on the surface of IECs (Hershberg and Mayer, 2000). These coordinated immune responses contribute to the activation of immune cells and production of cytokines, which initiates the appropriate immune response to bacterial infection. Therefore, most of the gut-associated immunological processes begin at the mucosal surface of the intestine. Sometimes, the final outcome of host and pathogen interaction is progression of a wide variety of inflammatory and immune-mediated processes (Berer et al., 2011; Chu et al., 2012).
Lynne et al. (2009) have identified three transmissible plasmids (TPs) that harbor drug resistance genes in S. Heidelberg strains isolated from poultry sources. The plasmid sequence analysis of these S. Heidelberg strains revealed a VirB/D type-IV secretory system (T4SS) core complex (Han et al., 2012) that is similar to Agrobacterium tumefaciens VirB/D T4SS (Cascales and Christie, 2004). A comparison of bacterial genome sequences revealed the presence of T4SS in Helicobacter pylori, Streptococcus suis, Bordetella pertussis, Brucella spp., Legionella pneumophila, and several other pathogenic bacteria (Corbel, 1997). These bacteria employ T4SS to inject virulence factors into host cells (Backert and Meyer, 2006).
The whole genome sequence data from 44 outbreak strains of S. Heidelberg (animals, retail meats, and human clinical isolates) revealed that 21 isolates possessed plasmids that encode T4SS (Hoffmann et al., 2014). The T4SS mediates various functions, which include translocation of proteins or toxins to the host cell, as well as bacterial conjugation (Grohmann et al., 2003). Specifically, the T4SS-expressing bacteria transport plasmids to a recipient bacterium or transfer DNA or protein molecules that allow the exchange of genetic material with the extracellular environment or host cells (Han et al., 2013). Our recent study revealed the increased expression of T4SS-encoding genes in S. Heidelberg during early-stage infection with macrophages (Gokulan et al., 2013). Furthermore, it also showed that S. Heidelberg TPs are responsible for multidrug resistance and increased virulence (Gokulan et al., 2013; Han et al., 2013).
There are limited studies available on IEC cytokine secretion profile during the early stage of infection. The main focus of this study was to evaluate the cytokine production of intestinal epithelial cells upon exposure to S. Heidelberg strains that harbor TPs, including those that encode the VirB/D4-T4SS, and compare it with a strain that lacks these TPs. Specifically, we aimed to address the involvement of IECs in cytokine production at the early stage of bacterial invasion (30 min and 1 h postinfection). In addition, we also analyzed the molecular mechanism of cell death caused by infection with S. Heidelberg that harbors specific TPs. Hereafter, in this study, we refer to strains that harbor TPs as virulent and strain that lack TPs as avirulent. Finally, evaluating the low level of cytokine production in local environment, specifically in intestinal epithelial tissue will be important to understand the epithelial barrier function and molecular mechanism of infection and pathogenesis.
Materials and Methods
Bacterial strains
Three S. Heidelberg isolates from poultry products that harbor multiple TPs were selected based on the results of plasmid sequencing (Han et al., 2012). These isolates were numbered 146 (containing VirB/D4 T4SS, IncA/C, an untypeable resistance, and IncI1 plasmids), 163, and 696 (both containing VirB/D4 T4SS, IncA/C, and IncFIB plasmids) (Table 1). S. Heidelberg isolate 819 (Kaldhone et al., 2008), a strain that lacks TPs, was used to compare the immunological and cytotoxicity effects of TPs in virulent strains. In earlier studies, strain 819 demonstrated a less-virulent phenotype, in that it was found to be significantly less able to invade and survive in IECs and macrophages compared to virulent strains (Gokulan et al., 2013). We also used transconjugant strains for cytotoxicity adapted from our previous study (Table 1) (Gokulan et al., 2013). For the studies reported here, a single bacterial colony was used to inoculate 2 mL Luria Broth (LB) media at 37°C with shaking at 250 rpm for overnight incubation. The following day, 300 μL of overnight culture were inoculated into 3 mL fresh LB media with shaking at 250 rpm at 37°C until the culture reached mid log phase (A600 between 0.6 and 0.8).
This table was adapted from our previous publication (Gokulan et al., 2013).
Intestinal epithelial cell culture
Human T84 epithelial cells (CCL-248™; American Type Culture Collection [ATCC]) were grown in Dulbecco's modified Eagle's medium (DMEM/F12 media) containing 2.5 mM
Macrophage cell culture
J774 mouse macrophage cells (TIB-67; ATCC) were grown in 75-mm culture flasks in DMEM supplemented with 2 mM glutamine, 10% FBS, penicillin (100 U/mL), and streptomycin (100 μg/mL). The cells were cultured at 37°C with 5% CO2 in 95% humidity. After growth, cells were detached from the culture flask using a cell scraper and washed with DMEM. The cell pellet was resuspended in enriched DMEM and plated into 24-well culture plates (4 × 105 cells/well). The culture plates were incubated at 37°C with 5% CO2 and 95% humidity for 24–48 h.
Cytokine measurement
The ratio of host cell to bacteria cells, 1:200, was selected based on our previous studies of bacterial invasion and survival using in vitro cell culture and in vivo animal models for Salmonella, Brucella, and Mycobacterial species (Khare et al., 2012; Gokulan et al., 2013). Bacterial cells suspended in DMEM (1 × 107 cells/well) were added to the wells containing epithelial cells. The plates were centrifuged at 500 rpm for 5 min and allowed to incubate for 1 h at 37°C in 5% CO2. To measure the cytokine production, cell culture supernatant was collected at 30 and 60 min postinfection. Cytokine assay was done using Bio-Plex Pro Human Cytokine (Bio-Rad., Inc.); a multiplex biometric ELISA-based immunoassay involving colored microspheres conjugated with a monoclonal antibody specific for the cytokine target proteins was used. The analysis was performed as per the manufacturer's guidelines using cell culture supernatant samples on the Bio-Plex Instrument (Bio-Rad, Inc.). All culture supernatant samples used in this study were analyzed in duplicate.
Visualization of necrosis with acridine orange and ethidium bromide
Macrophages were seeded onto 24-well cell culture plates at a density of 4 × 105 cells per well and incubated for 24 h. After incubation, cells were infected with S. Heidelberg virulent or avirulent strains to evaluate mechanisms for cell cytotoxicity. Noninfected macrophage cells served as negative control. Staining for cell death was performed at 1 h postinfection in separate sets of wells. For staining, media were removed and cells were washed twice with plain media followed by incubation with cell culture media containing gentamicin (50 μg/mL). Macrophages were stained for detection of cell death (autophagy/necrosis) according to the protocol developed by Ribble et al. (2005). Acridine orange (2 μg/mL) and ethidium bromide (2 μg/mL) dye were added to the media of each well and incubated for 15 min. The stained cells were visualized and photographed using an inverted fluorescence microscope (EVOS; Life Technologies).
Scanning Electron Microscopy
T84 cells were infected as described in the earlier section. After infection, cells were transferred in 1.5 mL vials and fixed for Scanning Electron Microscopy (SEM) by freshly made 4% glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA) at 4°C. The fixed samples were dehydrated, dried, mounted and further processed for SEM at the FDA NanoCore Facility (Jefferson, AR).
Statistical methods
Graph Pad (Graph Pad Software, Inc.) was used to calculate statistical differences for cytokine production. The unpaired t-test was used to calculate the statistical significance. Differences were considered statistically significant at p < 0.05.
Results
Early production of proinflammatory cytokines by epithelial cells after infection with S. Heidelberg strains
This study examined the cytokine production by intestinal epithelial cells during infection with TP-containing S. Heidelberg isolated from poultry sources. Cytokine production was analyzed after 30 and 60 min infection with the S. Heidelberg stains listed in Table 1. Interleukin (IL)-10, tumor necrosis factor-α (TNF-α), IL-17, IL-13, IL-2, and granulocyte macrophage colony-stimulating factor (GM-CSF) were evaluated in the cell culture supernatants after infecting with avirulent, as well as virulent, S. Heidelberg strains. The cytokine IL-10 is required for maintenance of gut immune homeostasis, as well as control of the inflammatory process through suppressing the expression of various proinflammatory cytokines to external stimuli (Pan et al., 2013). IECs that were not exposed to S. Heidelberg (control cells) produced a low level of IL-10 at the 30- and 60-min time points (Fig. 1A). The IECs that were exposed to the avirulent S. Heidelberg strain 819, which lacks TPs, also produced a low level of IL-10 at 30 and 60 min postexposure, comparable to that of the control cells (Fig. 1A). In contrast, IECs exposed to the virulent S. Heidelberg strains (146, 163 and 696) harboring TPs induced significantly higher levels of IL-10 at the 30-min time point (p < 0.02, p < 0.05 and p < 0.04, respectively; Fig. 1A). However, at 60 min, the IECs exposed to the virulent strains produced a lower level of IL-10 compared to the avirulent strain, with the exception of strain 696 (p < 0.01) (Fig. 1A). Significant differences were found while comparing the production of IL-10 at 30 min versus 60-min time points, at which strains 146 and 696 produced a statistically significant (p < 0.020 and p < 0.004) lower level of IL-10 at 60 min compared to the 30-min time point (Fig. 1A).
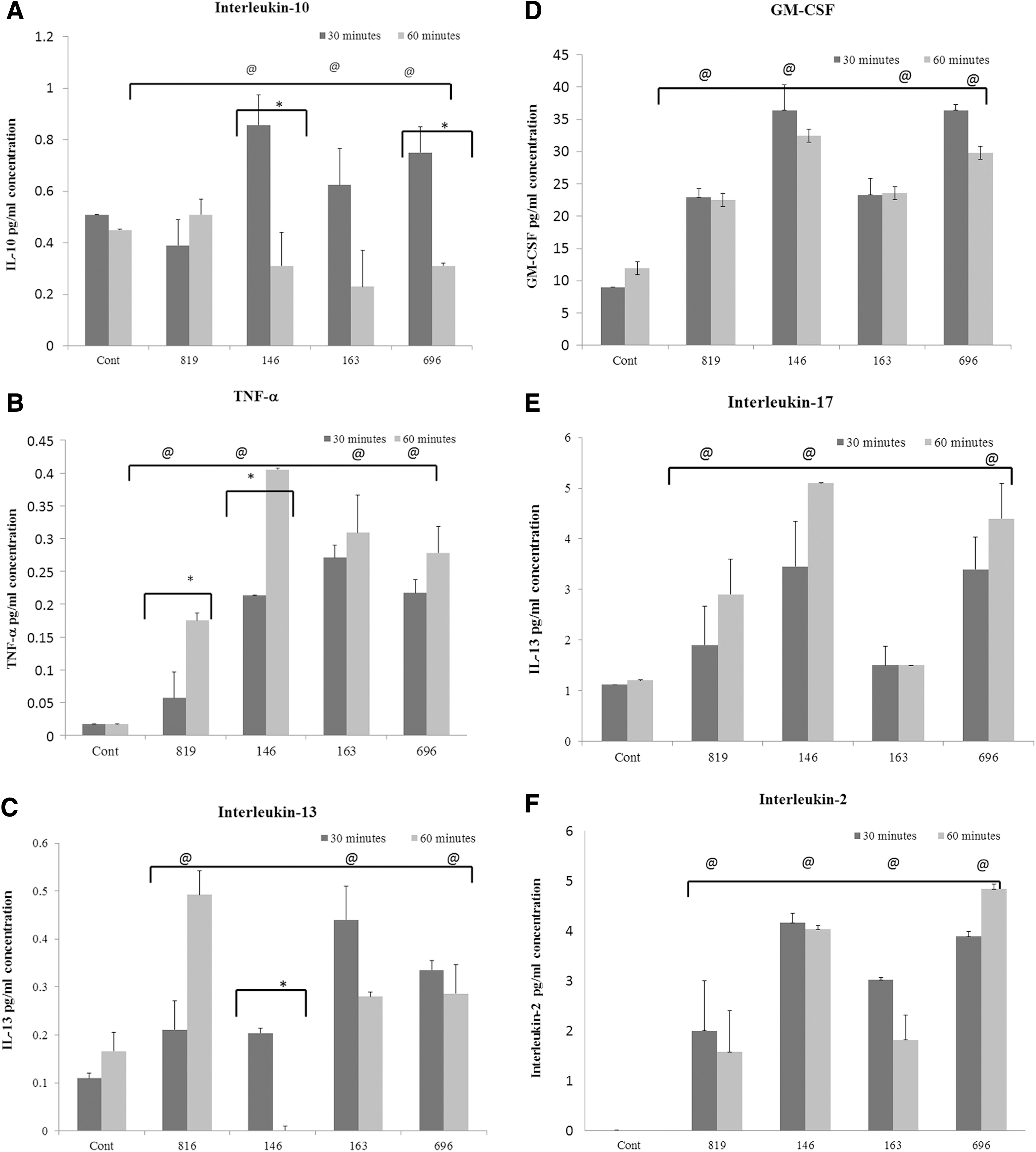
To evaluate whether IEC infection by the virulent and avirulent strains can also alter production of proinflammatory cytokines, the production of TNF-α in IECs was examined. The IECs that were not exposed to S. Heidelberg (control) produced a low level of TNF-α (0.02 pg/mL) and the levels were comparable at both time points (30 and 60 min) (Fig. 1B). Next, we compared TNF-α production between control cells with cells that were exposed to strain 819 that lacks TPs. The results showed that strain 819 induced a higher level of TNF-α than unexposed control cells (p < 0.05). Comparing the 30- and 60-min time points, strain 819 induced a significantly higher (p < 0.001) level of TNF-α production at 60 min (Fig. 1B). The cells exposed to virulent S. Heidelberg strains (146, 163 and 696) also produced a higher level of TNF-α at both time points, which was statistically significant (p < 0.001) compared to unexposed control cells. Next, we compared TNF-α production between cells that were exposed to virulent strains versus avirulent strains. The level of TNF-α production was significantly higher (p < 0.01, p < 0.002, and p < 0.045, respectively) in cells infected with virulent strains in comparison with the avirulent strain (Fig. 1B). A similar pattern was observed at both time points (30 and 60 min).
IL-13 plays a dual role in the intestinal tract; at lower concentrations it serves a protective function, whereas at higher concentrations it is associated with pathogenesis of intestinal diseases (Abraham and Cho, 2009). Our results showed that control IECs secreted a low level of IL-13 at both time points, as expected. Cells that were infected with the avirulent strain induced a significantly higher level of IL-13 at both time points; however, the level of secreted IL-13 was significantly higher (p < 0.03) at 60 min than at 30 min (Fig. 1C). IECs infected with virulent strains (163 and 696) induced a significantly higher level of IL-13 (p < 0.0001), with the exception of strain 146 at 30 min. Exposure to the avirulent strain 819 induced a significantly higher level of IL-13 compared to the virulent strains (p < 0.04) (Fig. 1C). The level of IL-13 in cells infected with strain 146 at 60 min was below the detection limit. Overall, the level of IL-13 produced from cells infected with the virulent S. Heidelberg strains decreased from 30 to 60 min postinfection.
This study also investigated the correlation between the production of proinflammatory cytokines and the bacterial pathogenicity of S. Heidelberg strains that are listed in Table 1. GM-CSF has been shown (Ouyang et al., 2008) to promote the maturation and proliferation of macrophages and promote inflammation by upregulating the release of proinflammatory cytokines during bacterial infection. The control IECs produced a basal level of GM-CSF and the levels were comparable at both time points. The cells exposed to the avirulent strain (819) produced a significantly higher level of GM-CSF (p < 0.0001) than control cells after 30 and 60 min exposure (Fig. 1D). The IECs that were exposed to virulent strains (146 and 696) produced a significantly higher level of GM-CSF (p < 0.01 and 0.002, respectively) than strain 819-exposed cells (Fig. 1D). Cells infected with strain 163 produced GM-CSF at a level comparable to cells infected with the 819 strain. A similar result was also observed at 60 min. Comparison of GM-CSF between 30- and 60-min time points showed no statistical difference between treatment groups.
IL-17 is recognized as an important player in the pathophysiology of infectious diseases and immune-mediated GI diseases. It regulates the proinflammatory cytokines, including TNF-α and GM-CSF, during early stage of invasion. Therefore, IL-17 levels were measured to see if there was any correlation between IL-17 and the other proinflammatory cytokines. Infection of IECs with both virulent and avirulent strains produced higher levels of IL-17 compared to uninfected control cells. IECs infected with strains 146 and 696 showed a statistically significant (p < 0.04 and p < 0.01) increase of IL-17 at both time points (Fig. 1E) compared to the avirulent strain. IL-2 is an immunoregulatory cytokine that plays an important role in the generation of immune cells. Cells infected with the avirulent strain produced a low level of IL-2 in comparison with those infected with virulent strains. However, virulent strain 146 induced a significantly higher level of IL-2 (p < 0.08) at 30 min postexposure (Fig. 1F) compared to control cells. The other two virulent strains (163 and 696) also induced IL-2 production, although the levels were less than strain 146. The 60 min postexposure data show that strains 146 and 696 produced a significantly (p < 0.02 and p < 0.08) higher level of IL-2 compared to the avirulent strain (Fig. 1F).
Detection of necrosis using acridine orange and ethidium bromide staining
We also examined the cytopathic effects of both virulent and avirulent strains of S. Heidelberg by employing IECs and mouse macrophages as a model system to investigate cell cytotoxicity. The production and accumulation of mucin on the apical surface of the polarized IECs prevented the capture of clear images after staining (data not shown). As a result, in this study, we have described results obtained from macrophages infected with both virulent and avirulent strains of S. Heidelberg. Earlier, we showed that S. Heidelberg strains that harbor TPs induce cytotoxicity in both IECs and macrophages (Gokulan et al., 2013). The control cells, which were not exposed to the bacteria, as well as cells that were exposed to the avirulent strain (819) appeared green in color. Acridine orange permeates through the intact cell membrane and stains the nuclei, which results in live cells appearing green in color (Fig. 2A, B). These results indicate a lack of cell death in control as well as the avirulent strain-treated groups (Fig. 2A, B). The ethidium bromide permeates the nuclei of cells that have lost cell membrane integrity, staining the cells a dark red color. Necrosis is generally visualized as dark red cells with scattered chromatin condensation and loss of plasma membrane integrity. Cells that were infected with virulent strains (that harbored TPs) showed mixed populations of live (green) (60–70%) and cells displaying characteristics of necrosis (dark red) cells (30–40%) at 1 h (Fig. 2D–F). This result suggests that the virulent strains induce cell death by necrosis. At 2 h postexposure, around 90% of cells were stained dark red, indicating the continuous necrosis process (Supplementary Fig. S1; Supplementary Data are available online at
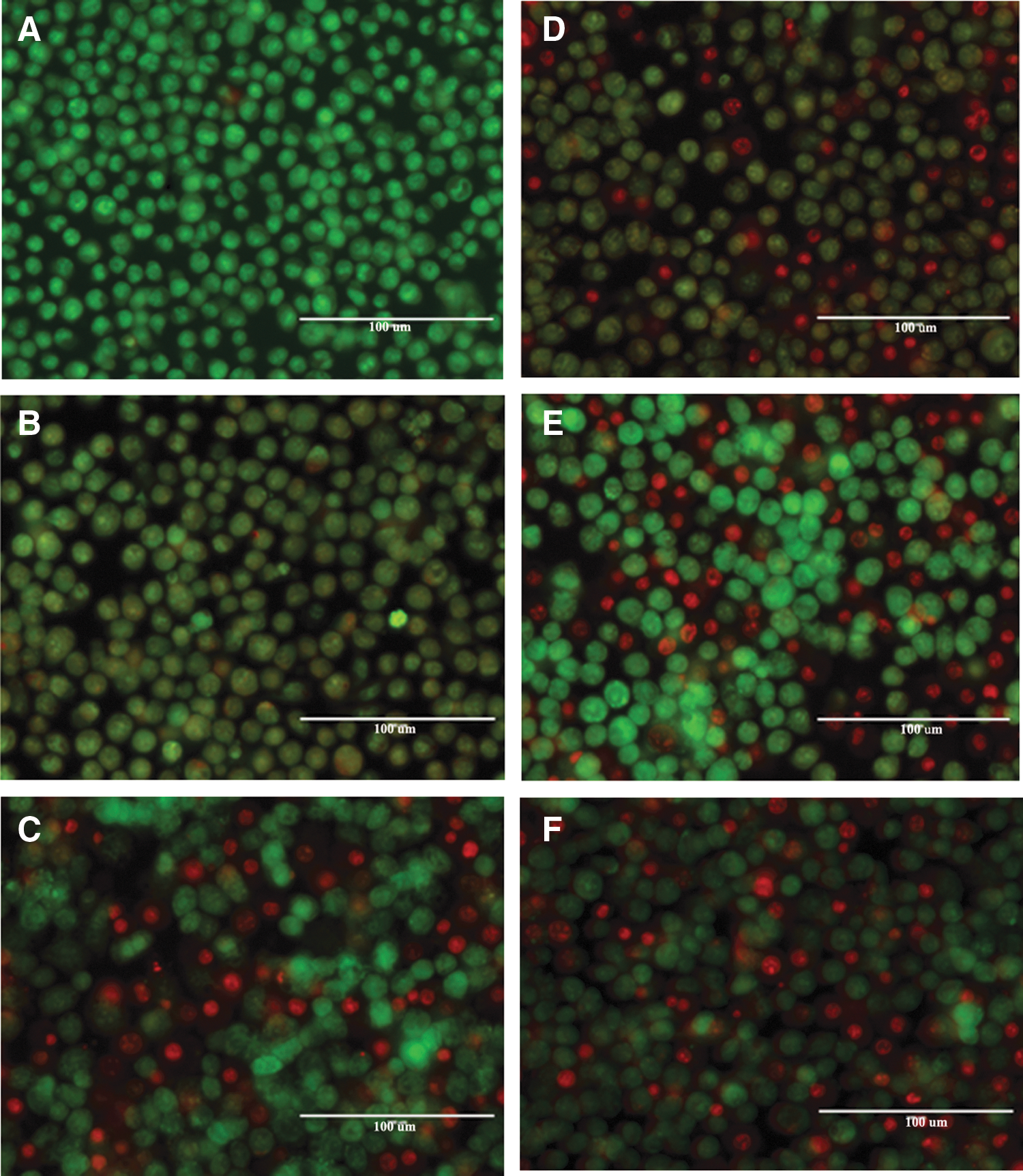
Acridine orange and ethidium bromide staining in macrophages during infection with S. Javiana and S. Heidelberg isolates. Staining of cell death was performed at 1 h postinfection.
We further confirmed the cell cytotoxicity mechanism using scanning electron microscopy (SEM). Necrosis was also analyzed through SEM at various time points (5 and 15 min), where cell necrosis was observed as early as 5 min after infection with virulent strains, in contrast to control and avirulent strains (Fig. 3A–D). The level of necrosis has been increased only in virulent strains, but not in the avirulent strain (in this study, we show representative data for one virulent strain and avirulent strain for 15 min postexposure time point: Supplementary Fig. S2). The SEM images show that cells with disintegrated membranes appear only in those samples exposed to the virulent (TPs-containing) S. Heidelberg strains. Thus, the virulent strains seem to induce cell death in macrophages through necrosis.

SEM imaging of macrophages.
Discussion
In this study, we aimed to address the immunological responses of IECs during the initial phase of bacterial infection. Our results show that strains of S. Heidelberg containing specific TPs induced different immune responses upon infection of IECs compared to those without TPs. The results demonstrated that T84 IECs produced lower levels of IL-10 when infected with strains of S. Heidelberg that harbored TPs. Like other pathogenic bacteria, virulent strains of S. Heidelberg have evolved and equipped themselves with several strategies to modulate the host immune response, including T4SS and the secretion of virulence factors. S. Heidelberg could possibly employ T4SS to directly inject virulence factors into host cells and these molecules could alter host cell signaling cascades for cytokine production.
The decreased level of IL-10 production could be attributed to increased inflammatory cytokine production. The decreased IL-10 may also compromise IEC barrier function and tight cell junction integrity. An in vivo study has shown that deficiency of IL-10 is associated with increased intestinal permeability, increased inflammation, and intestinal dysfunction (Madsen et al., 1999). Our earlier study has shown that virulent S. Heidelberg strains increase their expression of T4SS genes in the early phase of macrophage infection and alter host cell signaling pathways that include mRNA expression of cytokines, chemokines, and signal transduction molecules (Gokulan et al., 2013). S. Heidelberg infections dampen the secretion of anti-inflammatory cytokine IL-10, which may facilitate a successful invasion and persistence in host cells. An ex vivo tissue explant study demonstrated that production of endogenous IL-10 in mucosal tissue is important in preventing morphological changes and maintaining cellular integrity (Pan et al., 2013). IL-10 is an essential cytokine that is required to prevent the deleterious effects of inflammation in the gut (Shkoda et al., 2007; Pan et al., 2013).
The same group showed that flow cytometric analysis revealed that a certain percentage of epithelial cells secrete IL-10 (Pan et al., 2014). It also has inhibitory capability to secrete Th1 cytokines and impacts the shaping of the gut immune system. Inoculation of pathogens in germ-free mice (Matsumoto et al., 1992) as well as in IL-10-deficient mice resulted in the animals developing spontaneous chronic enterocolitis (Kuhn et al., 1993), intestinal inflammation, and irritable bowel syndrome (Wallden et al., 2012). Other studies suggest that deficiency of IL-10 is associated with intestinal inflammation in ulcerative colitis and inflammatory bowel disease in humans (Franke et al., 2008; Anderson et al., 2009). The mRNA expression data from our previous study revealed that S. Heidelberg strains with TPs altered the profile of cytokines, chemokines, and signaling pathways in mouse macrophages (Gokulan et al., 2013). To substantiate the protein secretion data, the cytokine mRNA expression analysis was performed and the results revealed that there was a difference in gene expression level between proinflammatory (TNF-α) and anti-inflammatory cytokines (IL-10) (Supplementary Fig. S3 and Supplementary Table S1).
We observed decreased levels of IL-10 in this study; hence, we expected reduced immunosuppressive function with increased production of TNF-α. Our results showed that increased levels of TNF-α is present only in cells that are exposed to the virulent strains of S. Heidelberg. The increased level of TNF-α indicates that virulent S. Heidelberg strains may trigger the proinflammatory response upon infection. This proinflammatory cytokine production could promote epithelial barrier disruption, bacterial adhesion, invasion, and persistence in the host. This evidence further supports our earlier finding that S. Heidelberg strains harboring TPs are highly invasive and persistent in macrophages and IECs (Gokulan et al., 2013). Several studies have shown that the elevated level of TNF-α is a major mediator for intestinal epithelial barrier dysfunction (Wang et al., 2012; Feng and Teitebaum, 2013). In vivo, these changes in cytokine expression may compromise epithelial barrier function, allowing bacteria and bacterial toxins to cross to the lamina propria and activate inflammatory cytokine secretion.
In this study, we observed that infection with virulent strains of S. Heidelberg increased the production of IL-17 in IECs. Activated Th-17 cells are the main source of IL-17 (Ouyang et al., 2008); however, other cell types, including innate immune cells, Paneth cells, and epithelial cells, are also capable of producing IL-17 (Cua and Tato, 2010). A high level of IL-17 is correlated with pathogenesis of immune-mediated diseases (Fossiez et al., 1998) and may play an important role in recruiting inflammatory cells during bacterial infection (Iwakura and Ishigame, 2006). Previously, it has been reported that the early stage of H. pylori infection resulted in increased level of IL-17 (Fox and Wang, 2007). IL-17 is also capable of upregulating gene expression of proinflammatory cytokines (TNF-α and GM-CSF) during bacterial infection. The secreted IL-17 activates the epithelial cells, which results in production of TNF-α and other proinflammatory cytokines at the site of infection (Iwakura and Ishigame, 2006; Caruso et al., 2008).
Our study shows that elevated levels of TNF-α and GM-CSF were found only in IECs infected with virulent strains. This effect could be correlated with the high level of IL-17 production by IECs infected with virulent strains with the exception of strain 163. IL-13 plays an important role in innate immune response. IL-13 is mainly produced by CD4 T cells (Fuss et al., 2004) and it is also secreted by several other immune cells, including gut-associated immune cells (Koyasu and Moro, 2011). In the gut immune system, IL-13 acts on the goblet cells to produce mucin, to attract eosinophils, and IgE production (Wu et al., 2011). Recent studies showed a direct connection of IL-13 with allergy and inflammatory disease, which is also required to fight against gut parasitic pathogen nematode (helminthes) (Bancroft et al., 1998). An in vivo experimental study has shown that IL-13 plays a homeostatic role in the mouse gut; specifically, it can limit IL-17-induced inflammation by blocking IL-17 production, as well as by increasing production of anti-inflammatory cytokine IL-10 (Newcomb et al., 2011, 2012). In addition, endogenously secreted IL-13 protects the gut against excessively secreted IL-17, as well as limiting the colitogenic process (Farmer et al., 2011).
In this study, we observed decreased level of IL-13 production at 60 min compared to 30 min postinfection, which can be explained in several ways; (1) virulent bacteria can modulate the signal transduction pathways on cytokine production in epithelial cells, (2) virulent strains induces cell necrosis that can lead to a low level of IL-13 production, (3) the proinflammatory cytokines may play a role by limiting the secretion of IL-13 and IL-10, and (4) bacterial toxins also may play a role in altering signal transduction pathways. In an avirulent strain, the above-mentioned events may occur at a low level; which can be correlated to a high level of IL-13 observed at 60 min in comparison to that at 30 min postinfection. A low level of IL-13 and IL-10 together could be also a contributing factor for bacterial invasion in virulent S. Heidelberg strains.
The GM-CSF, a hematopoietic growth factor, which promotes the proliferation and differentiation of myeloid progenitor cells could be induced by IL-13 and TNF-α. GM-CSF is secreted by various immune cells, including Paneth cells and epithelial cells (Hamilton, 2002; Fukuzawa et al., 2003). It has been reported that GM-CSF signaling is required for maintaining intestinal homeostasis, barrier function, and potentiating IEC proliferation (Fukuzawa et al., 2003). In contrast, several studies have shown that a high level of GM-CSF is responsible for the bacterial pathogenesis. For example, Salmonellosis is directly correlated with GM-CSF, because it potentiates the release of proinflammatory cytokines that include TNF-α, IL-1, and IL-6 (Metcalf, 1991). Earlier it has been shown that, in the absence of GM-CSF, a decreased level of proinflammatory cytokines was observed during S. typhimurium infection, which resulted in delay of onset of inflammatory pathology (Coon et al., 2009).
Our result shows that a high level of GM-CSF was observed in cells infected with virulent strains, except strain 146. The difference observed in strain 146 may be correlated with the absence of IncFIB plasmid that harbors virulence and antimicrobial strains. In contrast, the other virulent strain harbors IncFIB, IncA/C, and VirB/D4 plasmids. The high level of GM-CSF can therefore most likely be attributed to the synergistic effect of IL-13 and TNF-α. Together, these results suggest that secreted IL-17 and GM-CSF by IECs could induce proinflammatory cytokines during the early stages of S. Heidelberg invasion. This observation is in line with earlier reported human studies, in which H. pylori infection enhanced production of both IL-17 protein and transcriptome in gastric mucosa and lamina propria (Kabir, 2011). The increased levels of IL-17, TNF-α, and other proinflammatory cytokines may be a contributing factor for bacterial colonization and survival in macrophages and IECs (Gokulan et al., 2013). There are limited studies available on IEC cytokine profile during the early stages of bacterial infection.
Our study shows that IECs infected by S. Heidelberg secretes very low levels of IL-2. In spite of this, the virulent strain-infected cells produced two- to threefold higher than the avirulent strain, at the same time a decreased level of IL-10 was observed. Earlier studies have shown that αβ intestinal epithelial lymphocytes isolated from celiac disease produced high level of IL-2 and reduced level of IL-10, while comparing healthy versus pathological samples (Kolkowski et al., 2006). Mice infected with S. Dublin induced enhanced proinflammatory cytokines, including IL-2 (Eckmann et al., 1996). This study provides evidence for the relative contribution of IECs to the proinflammatory immune response during invasion by pathogenic S. Heidelberg.
Cell cytotoxicity is caused by chemical agents, drugs, secreted cytokines, and pathogenic microorganisms. The cell cytotoxicity event and morphological changes typically can be triggered by several ways that include necrosis, apoptosis, and autophagy. Host immune cells have the capability to kill invading pathogens by phagocytic mechanisms. To counteract this defense mechanism, bacteria can employ secretory systems to inject toxins into host cells, allowing pathogenic organisms to escape and further initiate infection. These toxins may prevent the host-mediated defense mechanisms and activate cytotoxic effects by necrosis. Recently, we demonstrated that virulent gene cytolethal distending toxin B (cdtB) in nontyphoidal S. Javiana induced cell cytotoxicity through autophagy (Williams et al., 2015a) (Supplementary Fig. S4). Interestingly in this study, the cytotoxicity mechanism induced by S. Heidelberg harboring TPs is by cell necrosis. Cytotoxicity results indicate that S. Heidelberg strains that harbor TPs (146, 163 and 696) are more virulent than strain 819 that lack TPs. Macrophages infected with virulent S. Heidelberg showed a significantly increased (80–90%, data not shown) number of necrotic cells at 2 h postinfection (Supplementary Fig. S1). Thus, cytotoxicity was further confirmed by SEM that clearly showed the loss of membrane integrity and release of intracellular components that is the signature event of necrosis (Supplementary Fig. S2). Overall these results support that S. Heidelberg strains that harbor TPs are more virulent compared to strains that lack TPs.
Conclusion
This study revealed that S. Heidelberg strains containing specific TPs (virulent strains) induced proinflammatory cytokines and decreased anti-inflammatory cytokines during the early stage of infection in intestinal epithelial cells. In contrast, the avirulent strain was unable to alter the anti-inflammatory cytokine IL-10, resulting in lower levels of proinflammatory cytokines. This study also demonstrates that S. Heidelberg virulent strains cause host cell cytotoxicity through necrosis. Based on these observations, we have proposed a model (Fig. 4A) for disease progression during infection of S. Heidelberg that harbors TPs. The avirulent strain (lacking TPs and T4SS) may cause less inflammation due to the high level of anti-inflammatory cytokines. On the other hand, the virulent strains may be introducing toxins into intestinal epithelial cells through the T4SS, which may alter the epithelial cell permeability and lead to activation of immune cells, which results in the production of multiple proinflammatory cytokines, inflammation, and compromised host immune response.
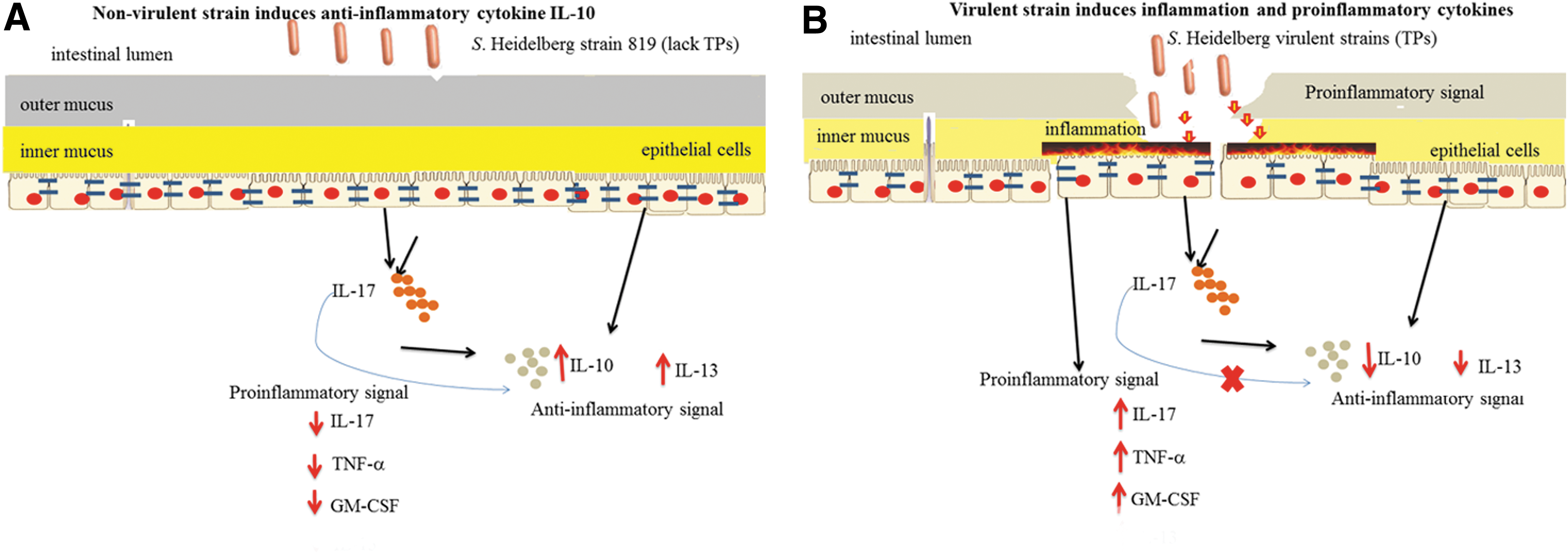
Intestinal epithelial cells (IECs) serve as a physical and biochemical barrier that separates the host from the intestinal microflora. The IECs can sense the pathogen and respond rapidly to the microbial stimuli and reinforce their barrier function by secreting cytokines and antimicrobial peptides into the lumen.
Footnotes
Acknowledgments
The authors would like to thank Dr. John Sutherland and Dr. Rajesh Nayak for their review of the article. Dr. K.G. is supported by FDA Commissioner Fellowship Program and Oak Ridge Institute for Science and Education. K.W. is supported by Oak Ridge Institute for Science and Education. Authors would like to thank Dr. Angel Paredes, NanoCore NCTR for SEM imaging.
Disclaimer
The findings and conclusions presented in this article are those of the authors and do not necessarily represent the views of the US Food and Drug Administration.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.