
Abstract
The chromatin remodeling activity of mammalian SWI/SNF complex is carried out by either Brahma (BRM) or Brahma-related gene (BRG-1). The BRG-1 regulates genes involved in cell proliferation, whereas BRM is associated with cell differentiation, and arrest of cell growth. Global modifications of histones and expression of genes of chromatin-remodeling subunits have not been studied in in vivo model systems. In the present study, we investigate epigenetic modifications of histones and the expression of genes in thioacetamide (TAA)-induced liver injury and regeneration in a mouse model. In the present study, we report that hepatocyte proliferation and H3S10 phosphorylation occur during 60 to 72 h post TAA treatment in mice. Furthermore, there was change in the H3K9 acetylation and H3K9 trimethylation pattern with respect to liver injury and regeneration phase. Looking into the expression pattern of Brg-1 and Brm, it is evident that they contribute substantially to the process of liver regeneration. The SWI/SNF remodeler might contain BRG-1 as its ATPase subunit during injury phase. Whereas, BRM-associated SWI/SNF remodeler might probably be predominant during decline of injury phase and initiation of regeneration phase. Furthermore, during the regeneration phase, BRG-1-containing remodeler again predominates. Considering all these observations, the present study depicts an interplay between chromatin interacting machineries in different phases of thioacetamide-induced liver injury and regeneration.
Introduction
L
There are various methods used to induce liver injury and regeneration to generate liver regeneration model. It includes partial hepatectomy and portal branch ligation (Bucher and Swaffield, 1973), LPS-dependent models for liver regeneration (Santoni-Rugiu et al., 2005), and pharmacological models (Palmes and Spiegel, 2004). In contrast to surgical animal models, chemically induced liver injury models are easier to establish and provide a better way to study many liver-associated diseases in humans (Palmes and Spiegel, 2004).
The drug-induced injury response produces mitogenic signals, such as cytokines and growth factors, which are responsible for reentry of quiescent hepatocytes into cell cycle (Michalopoulos and DeFrances, 1997). Several studies suggest that hepatocyte division begins at 12 h posttreatment, however, DNA synthesis starts later in the nonparenchymal cells (Taub et al., 1999). These events of cell cycle progression and cell division are regulated by a cascade of genes expressed at different stages of liver injury and regeneration process. It also includes various transcription factors and cofactors associated with gene promoters, which lead to the activation of gene transcription, specifically the chromatin remodelers.
The regulation of gene expression at the level of chromatin primarily involves two events, epigenetic modification of histones, and ATP-dependent chromatin remodeling. The two mechanisms operate in concert with each other. The mammalian SWI/SNF chromatin remodeling complex known as BRG-1-associated-factor (BAF) complex consists of two homologues having the ATPase subunit either BRM or BRG-1 (Mohrmann and Verrijzer, 2005). These ATP-dependent complexes coordinate the disruption of nucleosomes for binding of several transcription factors that enable gene activation in a permissive chromatin environment (Devine et al., 1999; Muchardt and Yaniv, 1999a). Although BRM and BRG-1 are 75% identical, their ratio differs during the cell growth and proliferation (Muchardt et al., 1996, 1998; Schnitzler et al., 1998; Sudarsanam and Winston, 2000).
There are reports available on the essential role of BRG-1 at critical developmental stages in BRG-1 null mice (Bultman et al., 2000), whereas BRM null mice showed some minor abnormalities in relation to animal size and uncontrolled cell growth in fibroblasts (Reyes et al., 1998). BRG-1 is reported to show compensatory effects for BRM in cell cycle control (Santoni-Rugiu et al., 2005). However, few studies are based on the functional switch of BRG-1 and BRM at the same promoter between activation and repression (Zhang et al., 2007). This ligand-dependent reprogramming of the SWI/SNF complex allows it to differentially serve as either a corepressor or a coactivator of transcription at the same promoter. Epigenetic modifications of histones have distinctively been associated with different states of cellular activities. Acetylation of histone H3 at lysine 9 (H3acK9) is often associated with transcriptionally active chromatin and recognized by bromodomain of proteins (Kouzarides, 2007).
Recruitment of these proteins facilitates binding of other cofactors which leads to gene transcription. Trimethylation at H3K9 (H3me3K9) is a marker for transcriptionally repressed chromatin and recognized by chromodomains of proteins and help in recruitment of heterochromatin protein 1 (HP1) to generate heterochromatin state of gene (Nielsen et al., 2001). Whereas, phosphorylation of histone at serine 10 (H3phS10) is associated with chromosome condensation and is a marker of M-phase of the cell cycle (Yun, 2011). These phosphorylated histones are recognized by proteins containing phospho-Ser/Thr-binding domain to regulate cell cycle progression (Ruthenburg et al., 2007; Reinhardt and Yaffe, 2013).
Liver regeneration includes transient upregulation of genes, particularly which are involved in protein synthesis and cell growth (Haber et al., 1993). Many of these transient expressions may remain upregulated throughout the proliferative phase of liver regeneration. Hence, it was hypothesized that the epigenetic mechanisms must be operative during this proliferative/regenerating phase.
In the present study, selected epigenetic modifications of histone H3, such as H3acK9, H3me3K9, and H3phS10, were studied during liver injury and regeneration. The expression of genes, such as alpha-fetoprotein (Afp), hepatocyte nuclear factor-4α (Hnf-4α), liver regeneration factor (Lrg-21), and specifically, the SWI/SNF chromatin remodeler subunits Brg-1 and Brm genes, were studied. There are very few reports available on BRG-1-mediated G1 cell cycle arrest in partial hepatectomy liver regeneration model (Strobeck et al., 2002). However, there are no such reports available on direct association of BRG-1 and BRM subunit of SWI/SNF chromatin remodeling complex and model systems of liver regeneration in vivo. In the present study, TAA-induced liver injury and regeneration model was chosen to observe the association of BRG-1 or BRM subunit of SWI/SNF chromatin complex, if any, during liver injury and regeneration process based on gene expression.
Since TAA has a relatively short half-life and provides a larger window for observing subtle changes during process of liver injury and regeneration (Mangipudy et al., 1995; Chilakapati et al., 2005), TAA-induced liver injury model was used in the present study. Our results provide some evidence in support of the hypothesis that the two regulatory processes, ATP-dependent SWI/SNF-associated chromatin remodeling, and histone modifications operate together, but distinctively during liver injury and regeneration.
Materials and Methods
Animal handling
Animal housing and liver injury model: pathogen-free female Swiss albino mice (12–14 weeks old) housed under hygienic and uniform conditions (relative humidity 45–75%, temperature 22°C ± 3°C, and photoperiodicity of 14 h of artificial light), from in-house animal facility were used for all the experiments (CPCSEA Registration Number: 565/GO/AB/02/CPCSEA). Institutional animal ethics committee guidelines were followed in housing, handling, experimenting, and sacrificing the animals.
A group of 42 mice were divided into control and treatment groups. Single intraperitoneal injection of TAA (65 mg/kg of body weight of mice) was given to each mice (treatment group), whereas the mice that were kept as control were injected with saline. Out of the 42 mice, 36 acted as treatment group and 6 acted as control. Mice were sacrificed at designated time points at 0, 12, 36, 48, 60, 72, and 360 h postinjection for sample collection. The blood was collected and kept at 4°C overnight for clotting. Serum was collected from clotted blood by centrifugation at 3000 g for 10 min at 4°C and stored at −80°C until further use. Liver was excised out at designated time points from sacrificed mice. Any one lobe of the liver was collected and fixed in formalin for histological analysis. Approximately 200 mg of liver tissue was stored in 2 mL of RNAlater for RNA isolation and rest of the liver was stored at −80°C for biochemical analysis and histone isolation.
Liver injury and regeneration model was characterized by biochemical and immunological parameters. Established protocols were used to study serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels (in Supplementary Data; Supplementary Data are available online at
Materials
Antibody against proliferating cell nuclear antigen (PCNA) was purchased from Santa Cruz Biotechnology. Histone H3-modification antibodies against H3acK9, H3me3K9, and H3phS10 were purchased from Abcam. Antibodies against BRG-1 and BRM were also procured from Abcam. Goat anti-rabbit IgG-HRP-conjugated antibody and goat anti-mouse IgG-HRP-conjugated antibody for western blotting (WB) were from Bangalore Genei (now, Merck Limited). For immunofluorescence (IF), secondary antibody of Alexa Fluor 488 donkey anti-rabbit IgG (H+L) was obtained from Life Technology. ELISA Kit for TNF-α and IL-6 were purchased from R&D Systems. Polymerase chain reaction (PCR) Kit, Omniscript reverse transcriptase, RNase-free water, RNAlater, and RNase inhibitor were from Qiagen, Hilden, Germany. SYBR green was purchased from Applied Biosciences. Amersham Hyperfilm ECL high-performance chemiluminescence film and Amersham ECL Western Blotting Detection Reagent were from GE Healthcare Limited. Trans-Blot (nitrocellulose membrane) was from Bio-Rad Laboratories. All biochemical reagents, forward and reverse primers of genes, were purchased from Sigma-Aldrich Corporation.
Methods
Morphological evaluation of liver injury
Liver was harvested at the time of sacrifice and washed in saline. Photograph was taken to evaluate the extent of liver damage at different time points of injury and regeneration.
Hematoxylin and Eosin staining
Paraffin-fixed liver sections were processed with different grades of alcohol and stained with Hematoxylin and Eosin (HE) stain to evaluate necrosis, infiltration of immune cells, and cell division. Hepatocyte division was evaluated by counting cells in nonoverlapping fields on liver sections (Nostrant et al., 1978).
PCNA staining and immunohistochemistry
Paraffin-fixed liver sections were deparaffinized and rehydrated, followed by heat-mediated antigen retrieval, based on the established protocol (Shi et al., 1991) with some modifications. Slides were kept in 0.3% H2O2 for quenching and immunostained with anti-PCNA antibody followed by counterstaining with Hematoxylin (Yu et al., 1992). Cell proliferation score was calculated by counting PCNA-stained positive cells in nonoverlapping fields of liver section. Microscopic analyses were done on Nikon Eclipse 100 compound bright-field microscope with artificial light source and E400 Nikon Eclipse. NIS Elements Version 3.2 software was used for imaging and documentation, whereas ImageJ software was used for quantification.
Histone extraction from liver tissue and WB
Nuclei and histones were isolated following standard procedures (Song et al., 2011) with minor modifications. Isolated histones were analyzed by resolving on SDS-18% PAGE (Laemmli, 1970), electrotransferred to nitrocellulose membrane followed by WB using histone modification specific primary antibodies H3phS10 (1:500), H3acK9 (1:6000), and H3me3K9 (1:1000). Goat anti-mouse IgG-HRP-conjugated secondary antibodies were used and signals were detected by enhanced chemiluminescence (ECL). For the quantification of histone bands on gel/nitrocellulose membrane/x-ray film, Quantity One 1-D Analysis Software from Bio-Rad was used.
Nuclear extract preparation and WB for BRG-1 and BRM
Nuclear extract was prepared from liver tissue following standard protocol (Gorski et al., 1986) with minor modifications. Briefly, the nuclear pellet prepared as described earlier was suspended in lysis buffer (15 mM Tris-HCl, pH 7.5, 0.5 M KCl, 2 mM MgCl2, 10% glycerol, Tween 0.1%) to a final concentration of 1 mg/mL of DNA and allowed to stand on ice for 1 h. It was subsequently centrifuged at 22,000 g for 25 min. The clear supernatant was analyzed by resolving on SDS-7% PAGE, electrotransferred to nitrocellulose membrane and probed with anti-BRG-1 and anti-BRM antibodies (1:200). Goat anti-mouse IgG-HRP acted as secondary antibodies. The signals were detected by ECL.
Immunofluorescence
Established protocol was used to study immunofluorescence (Wang et al., 2005) with some modifications. Serial sections taken on adjacent slides were used for HE staining and IF study. Liver sections for IF study were taken on poly-lysine-coated slides and processed with different grades of alcohol followed by heat-mediated antigen retrieval. Semi-dried liver sections were incubated overnight with primary antibodies (1:50) in phosphate-buffered saline (PBS), such as H3phS10, H3acK9, or H3me3K9. The slides were washed in PBS Tween thrice, subsequently incubated for 1 h in secondary antibody (1:200), and counterstained with DAPI. Microscopy was performed on E400 Nikon Eclipse. NIS Elements Version 3.2 software was used for imaging and documentation purpose.
RNA isolation
Total RNA was isolated from liver tissue using TRIzol reagent according to the manufacturer's protocol (Sigma-Aldrich). RNA was reverse transcribed using Omniscript Reverse Transcriptase Kit obtained from Qiagen.
Quantitative PCR
Quantitative analysis of gene expression by comparative C(T) method was performed according to standard protocols (Schmittgen and Livak, 2008). PCR cycle of denaturation at 95°C for 5 min, 40 cycles of 20 s at 94°C, 20 s at 61°C for primer annealing and extension followed by a dissociation cycle (melting curve) at 65°C for 0.5 s and 95°C for 0.5 s, were used. The forward and reverse primers were used for amplifications of Afp, Hnf-4α, Lrg-21, Brg-1, and Brm genes are listed in Table 1. Glyceraldehyde-3-phoshpate dehydrogenase (GAPDH) was used as internal control to measure relative increase in expression in terms of fold change normalized to GAPDH. BIO-RAD 1001 Real-time PCR system was used for PCR run and data analysis.
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) was performed following standard procedures (Wagschal et al., 2007a, 2007b). Briefly, nuclei were isolated from treated and untreated mice liver tissues at different time points and were subsequently digested with 10 U of micrococcal nuclease (MNase). The digested nuclei were centrifuged and the supernatants (S1) were collected. The pellets were resuspended in 1 mL of dialysis buffer (50 mM Tris-HCl, pH 7.5; 5 mM EDTA, pH 8.0; 0.1 mM PMSF, and 5 mM sodium butyrate) and dialyzed overnight at 4°C. The dialysates were subsequently centrifuged and supernatant (S2) and pellet (P) were collected. Ten micrograms each of S1 and S2 were mixed together and the volume was made to 1 mL with ChIP incubation buffer (l50 mM NaCl; 50 mM Tris-HCl, pH 7.5; 5 mM EDTA, pH 8.0; 0.1 mM PMSF, and 5 mM sodium butyrate). To this, 8 μg of the specific antibody was added and rotated for 12–16 h at 4°C at 20–30 rpm. Subsequently, 0.0125 g of protein-A agarose beads was added to samples and further incubated for 4 h at 4°C and centrifuged to collect supernatant as unbound fraction. Beads were washed and resuspended in 500 μL of elution buffer (50 mM NaCl; 50 mM Tris-HCl, pH 7.5; 5 mM EDTA, pH 8.0; SDS 1%, 0.1 mM PMSF, and 5 mM sodium butyrate) and DNA sample was eluted from the antibody/protein A beads. Furthermore, the immunoprecipitated DNA samples were used for PCR amplification.
The sequence of BRG-1 primers for promoter-specific region was designed using Eukaryotic Promoter Database and Primer-BLAST (
Results
Distinct phases of liver injury and regeneration in TAA-treated mice
Pilot experiments were performed to establish mice liver injury and regeneration model. Single injections of different TAA doses were administered to find out a dose that induced hepatocyte necrosis near centrilobular region and where survival was up to 80% (Supplementary Fig. S1A). Based on the above observation, the TAA dose of 65 mg/kg body weight was chosen for the present study. Following treatment, mice were sacrificed at designated time points and liver were promptly harvested as described in Materials and Methods. Subsequently, they were analyzed by photography, HE staining, and PCNA staining. The TAA-induced liver injury was evaluated by looking into mice liver morphology and histopathology, which indicated significant damage, swelling, and necrosis of hepatocytes with infiltration of immune cells at initial time points, when compared with control (Fig. 1A). Infiltration of mononuclear cells was clearly evident at 12 h, which showed gradual increase up to 36 h posttreatment (Fig. 1A-panel 2). This observation was further supported by biochemical and immunological parameters. Evaluation of serum AST, ALT, tissue SOD, catalase, and lipid peroxidation levels, and serum TNF-α and IL-6 levels also indicated that the injury peaked at 36 h (Supplementary Figs. S2A–D and S3A–C). The regeneration phase that followed subsequently was marked by an increase in hepatocyte division. Dividing hepatocytes were clearly found at 60 and 72 h posttreatment (Fig. 1A). The total number of mitotic figures were counted in random fields in liver section and plotted against time, which showed peak at 60 h (Fig. 1B). It was also supported by cell proliferation assay. The cell proliferation was confirmed by PCNA expression and showed maximum number of hepatocytes in the proliferating stage at 60 and 72 h posttreatment (Fig. 1A-panel 3, C). The mitotic index and the PCNA level also followed similar patterns and showed peak at 60 h. Therefore, from the above observations, it can be hypothesized that the change in treated liver can be divided into two distinct phases: (1) Injury phase lasting up to 36 h post-TAA injection, and (2) Regeneration phase that starts post 36 h.
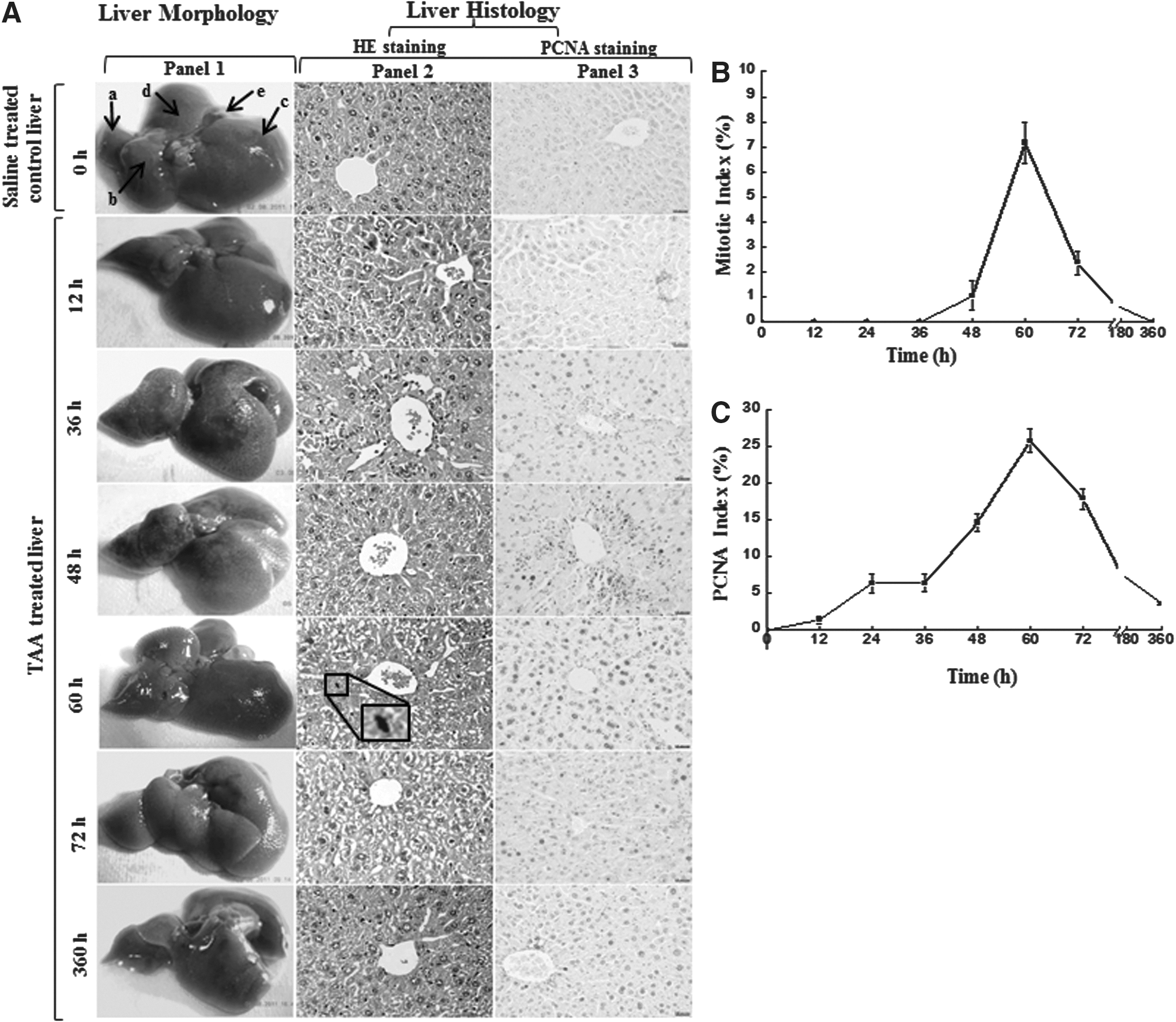
The morphological and histological assessment of thioacetamide-induced liver injury and regeneration.
Global histone H3 modification profile during liver injury and regeneration
Of all the four core histones, the histone H3 has the longest N-terminal tail, where maximum number of regulatory modifications has been reported (Kouzarides, 2007). Global pattern of histone H3 modifications, such as H3phS10, H3acK9, and H3me3K9, were studied by WB and IF, where anti-H3 signal was taken as control. Western blot analysis showed absence of H3phS10 signal in the injury phase, however, its signal showed subsequent increase between 60 and 72 h. Since H3phS10 is a maker of M-phase of cell cycle, it indicated that maximum number of cells were present in M-phase of the cell cycle at 60 and 72 h, TAA posttreatment (Fig. 2A). It correlated well with HE staining and PCNA staining data. The above observation was also supported by results obtained from IF using anti-H3phS10 antibody. It showed strong signals at 60 and 72 h posttreatment (Fig. 3 & Supplementary Fig. S4A). It clearly indicates that the histone H3phS10 increased during cell proliferation and regeneration phase.

Global histone modification profiling in different phases of liver injury and regeneration.

Immunofluorescence-mediated examination of histone modification in liver sections. Serial sections taken on adjacent slides were used for HE staining and IF study. Liver sections obtained from treated and untreated mice were deparaffinized and immunostained with antibodies against H3phS10, H3acK9, and H3me3K9 (Green). These were then counterstained with DAPI to visualize nuclei and only merged images are shown. The images shown are representative of data from three different sets of experiments. Magnification: 400×, Scale bar: 50 μm. IF, immunofluorescence.
The H3acK9 signal showed exponential increase up to 36 h with subsequent decline till 60 h, where it reached at the initial basal level. This premitotic increase in its signal after 12 h of TAA treatment (Fig. 2A) clearly suggests that some acetylation events might be responsible for activation of stress-related genes. This observation was further supported by the immunostaining data. The H3acK9 signal also increased up to 60 h, which decreased subsequently (Fig. 3 & Supplementary Fig. S4B). It clearly indicates that the initial increase in H3acK9 is well correlated with the initial injury phase.
H3me3K9 signal was found to increase up to 12 h then maintained up to 60 h followed by a gradual decline. It showed a quasi-biphasic pattern. Its peak around 48 h coincided with the initiation of phosphorylation of H3 at serine 10, and increase in the mitotic index (Fig. 2A). Similar biphasic pattern was demonstrated by our IF data. It possibly indicates repressed chromatin and downregulation of some genes associated with stress and metabolic processes (Fig. 3 & Supplementary Fig. S4C).
Differential expression of Afp, Hnf-4α, Lrg-21, and genes of chromatin remodeling subunits Brg-1 and Brm during liver injury and regeneration
These genes were selected based on their functional association with regeneration, maintenance, as well as liver injury, and cell cycle regulation. Quantitative PCR analysis was carried out to evaluate the expression level of these genes. Based on the results of semiquantitative PCR (data not shown), selective time points were chosen for further quantitative analysis. The Afp expression evaluated through real-time PCR showed 8.5-fold increase at 36 h posttreatment. It reached maximum at 60 h with 17.4-fold increase, indicating Afp induction and reactivation during liver injury and regeneration (Fig. 4A). Furthermore, our observation was supported by biochemical analysis, HE staining, and PCNA staining data, which showed peak of injury at 36 and peak of cell proliferation at 60 h (Fig. 1B, C). We also quantified expression profile of Hnf-4 using real-time PCR, which showed upregulation with a maximum of 4.1-fold increase at 60 h (Fig. 4A). Furthermore, the expression of stress-induced gene Lrg-21 was examined and showed a 2.6-fold increase at 36 h posttreatment, which subsequently declined (Fig. 4A), indicating its association with liver damage and stress.
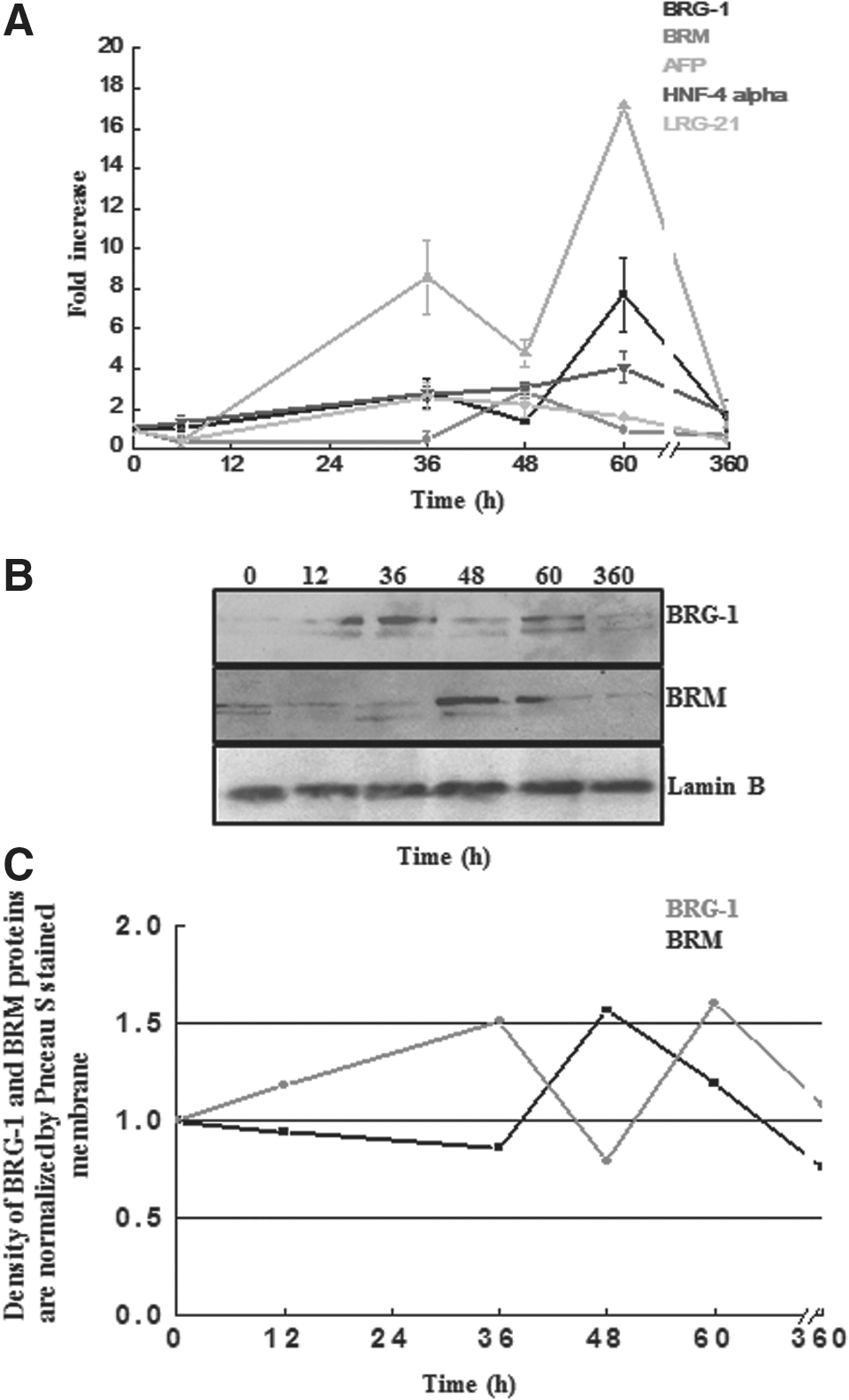
Expression profile of AFP, HNF-4α, LRG-21, BRM, and BRG-1 genes at transcript level as well as expression of BRG-1 and BRM at protein level during the process of liver injury and regeneration.
ATP-dependent chromatin remodeler SWI/SNF is known to have either BRG-1 or BRM as its ATPase subunit (Workman and Kingston, 1998). To find out which type of chromatin remodeling subunits were expressed during liver injury and regeneration, expression pattern of Brg-1 and Brm genes were examined during liver injury and regeneration using real-time PCR. Our results demonstrated significant increase in the level of Brg-1 gene expression at 36 h, which decreased at 48 h and again peaked at 60 h (Fig. 4A). Brg-1 showed biphasic curve with 2.7-fold increase at 36 h and 7.7-fold at 60 h (Fig. 4A). The first smaller peak coincided with the injury phase, whereas the second peak coincided with the regeneration phase. The expression pattern of Brm gene was also evaluated similarly. The expression of Brm pattern was distinctively different from that of Brg-1 (Fig. 4A). The Brm expression peaked at 48 h, which coincided with the decline in injury phase and preceded the regeneration phase.
Similarly, the expression level of BRG-1 and BRM proteins were also analyzed by WB. The BRG-1 protein expression showed a significant increase at 36 and 60 h during injury and regeneration phase, respectively (Fig. 4B, C), which was also consistent to their corresponding transcription level expression. Likewise, the BRM protein expression showed significant increase at 48 h followed by gradual decrease in the expression (Fig. 4B, C), correlated to the corresponding Brm mRNA expression.
From the above findings of RNA and protein level expression profiles of BRG-1 and BRM at different phases of injury and regeneration in mice liver, it is hypothesized that the chromatin remodeler SWI/SNF might have different ATPase subunits during different phases of liver injury and regeneration. We also intended to investigate the transcriptional state of Brg-1 promoter during different phases of injury and regeneration of mice liver, to validate our hypothesis that the differential expression of Brg-1 at injury and regeneration is mediated by a change in histone H3 modification marks of the chromatin in the promoter region of Brg-1. Hence, a ChIP assay was performed during different phases of injury and regeneration using site-specific histone modification antibodies H3acK9 and H3me3K27 for binding on the soluble chromatin. Subsequently, the bound DNA was amplified using specific region of Brg-1 (−689 to −461) promoters. The ChIP-PCR result for Brg-1 promoter analysis indicated increased H3acK9 signals (a mark of transcriptionally active chromatin) at 36 and 60 h similar to the expression of BRG-1, both at RNA and protein levels. On the contrary, there was reduced signals for H3me3K27 (a mark of repressed state of chromatin) at the corresponding 36 and 60 h and the signal intensity increased at 360 h in late phase of regeneration/recovery (Fig. 5). Data from the ChIP analysis further supported that the Brg-1 gene promoter becomes transcriptionally permissive during injury and regeneration phase with increase in the expression of BRG-1 at mRNA and protein levels.
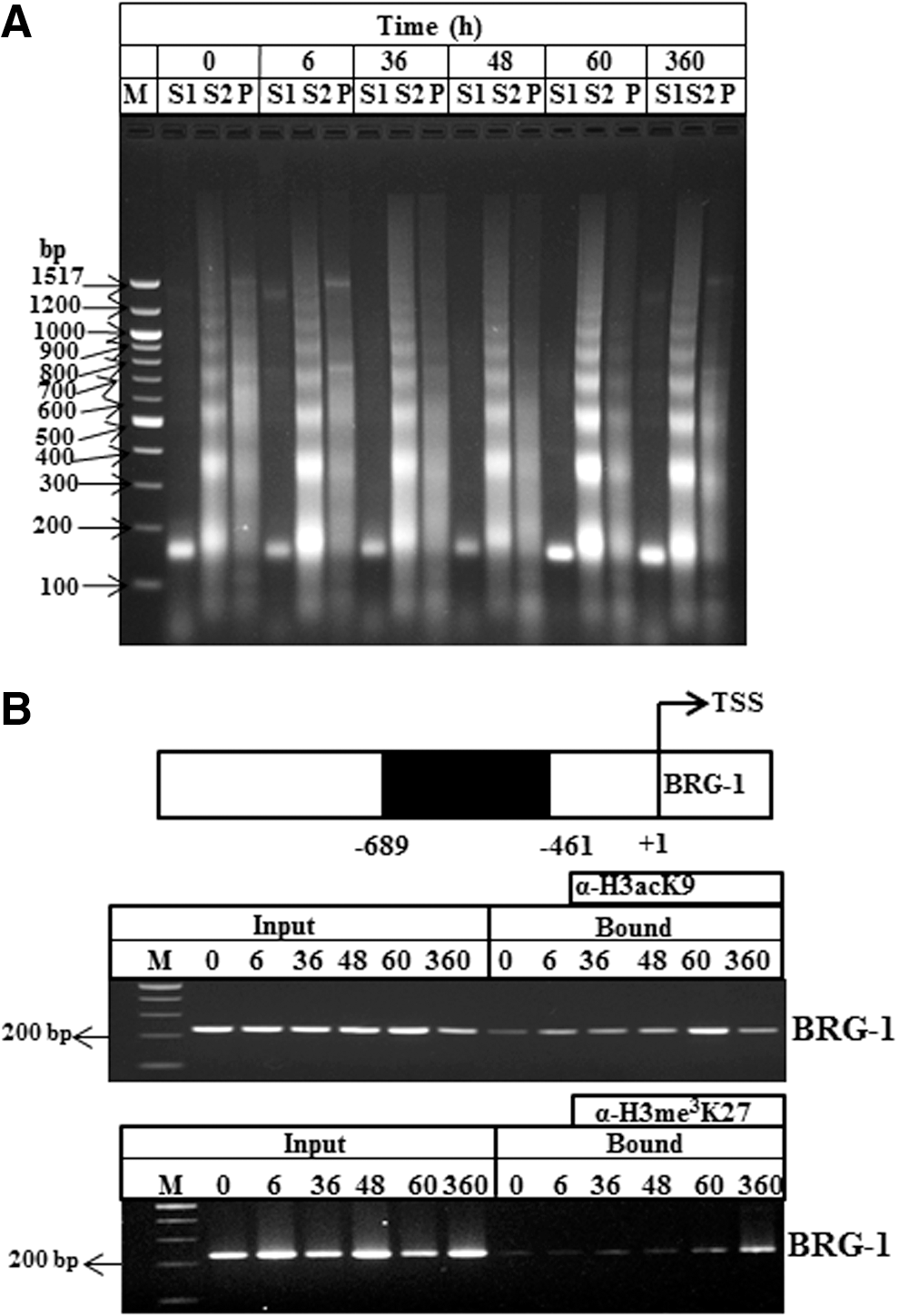
ChIP assay.
Discussion
Drug-induced liver injury and regeneration models have been widely used to understand the mechanism of liver-associated disorders such as cirrhosis, fibrosis, hepatitis, and hepatocellular carcinoma (Sherlock, 1968, 1984; Popper, 1972; Ferrell, 2000; Waris and Siddiqui, 2003; Bataller and Brenner, 2005; Schuppan and Afdhal, 2008). Extensive work has been done to characterize the mechanism of these systems using biochemical, histological, and immunological parameters (Michalopoulos and DeFrances, 1997; Sturgill and Lambert, 1997; Fausto et al., 2006; Michalopoulos, 2007). However, there are very few reports available on epigenetic regulation of liver disorders such as alcoholic liver disease and hepatocellular carcinoma (Kondo et al., 2007; Mandrekar, 2011; Page, 2015). In the present study, involvement of two epigenetic mechanisms that is, histone modifications and ATP-dependent chromatin remodeling, together in liver regeneration process using in vivo system is reported for the first time. Our findings provide certain evidence which depict that the two regulatory processes, ATP-dependent SWI/SNF-associated chromatin remodeling and histone modifications, might operate together, but distinctively during liver injury and regeneration.
Histone posttranslational modifications such as acetylation and methylation are known to regulate the expression of genes. Each site-specific modification on a particular histone type of a nucleosome codes for either activation or repression of the gene within that nucleosome. For example, H3acK9 leads to transcriptional activation, whereas H3me3K9 marks for transcriptional repression (He and Lehming, 2003). Similarly, BRG-1 and BRM-mediated SWI/SNF chromatin remodeling also regulates gene expression and are correlated to histone modification (Tie, 2012; Wu, 2014).
Our data reveal biphasic expression pattern of Brg-1 during liver injury and regeneration. In the present study, the first peak of BRG-1 was observed during injury phase that is, at 36 h. The levels of proinflammatory cytokines, such as TNF-α and IL-6, were also observed to be maximum at these time points, which indicated toward involvement of BRG-1 in the transcriptional activation of proinflammatory genes. Our results are supported by recent observations that BRG-1 is recruited to Tnf-α and Mcp-1 gene promoters in case of acute kidney injury (Naito et al., 2009). Furthermore, BRG-1 recruitment increases the transcription of proinflammatory mediators, such as Tnf-α, Mcp-1, and RNA polymerase II (Pol II) levels, indicating the process to be transcriptionally mediated. Also, BRG-1 in association with Nrf2 is reported to selectively mediate induction of heme oxygenase-1 (HO-1), a cytoprotective gene in response to oxidative stress (Zhang et al., 2006). Thus, probably the increased inflammatory and stress-related genes during the injury phase in our system, contains BRG-1 in SWI/SNF complex at their promoters. In terms of global histone modification, H3acK9 is also observed to peak at the same time points that is, during injury phase. Reports are available in support of this observation that suggests histone acetylation and prebound BRG-1 is characteristic of early inflammatory genes such as Tnf-α (Sullivan et al., 2007).
The second peak of BRG-1 was observed in the regeneration phase that is, at 60 h. This is the time point where H3phS10 and mitosis were also at the peak. Conversely, the expression of BRM peaked at 48 h and coincided with peak of H3me3K9 at 48 h, both of which gradually decreased. Thus, histone acetylation and BRG-1 peak during the initial injury phase was followed by histone methylation and BRM expression, which was further replaced by histone phosphorylation and BRG-1 expression during regeneration phase in the present study. It is known that H3me3K9 recruits HP1 and brings about transcriptional repression (Sims et al., 2003; Schotta et al., 2004). On the other hand, H3phS10 by Aurora B disrupts this HP1-H3me3K9 interaction and initiates M-phase of the cell cycle (Fischle et al., 2005). Our observations are also in conformity with these reports that when H3phS10 increases during proliferative phase, there is a decline in the level of H3me3K9.
Studies on cell cycle events in HeLa cells have revealed that several subunits of SWI/SNF complex, including BRG-1 and BRM, are phosphorylated at G2/M transition (Muchardt et al., 1996; Reyes et al., 1997; Stukenberg et al., 1997; Sif et al., 1998). Concomitant with phosphorylation, BRM is targeted for degradation and is only resynthesized upon exit from M-phase (Muchardt and Yaniv, 1999b), whereas, the BRG-1 does not undergo such degradation. These reports also justify the decline in the expression of BRM, both at the transcript and protein levels, observed before proliferative phase in the present study. BRG-1expression, both at the transcript and protein levels suggest that BRG-1-containing SWI/SNF complex are involved in stress and inflammatory gene regulation during injury phase and cell proliferation during regeneration phase (Dunaief et al., 1994). On the other hand, BRM-containing SWI/SNF complex is associated with cell differentiation and cell cycle arrest (Coisy-Quivy et al., 2006) and probably it is involved in the regulation of genes that carry out the transition from injury to regeneration phase.
Initially, it is evident from the study that the hepatocytes were the major population of liver cells participating in the regeneration process; however, further examination based on specific cell type analysis of liver cells involved in injury and regeneration process is required. Looking into the overall expression pattern of BRG-1 and BRM, it is evident that the SWI/SNF remodeler might contain BRG-1 as its ATPase subunit during injury phase between 12 and 36 h. Whereas, BRM-associated SWI/SNF remodeler was probably predominant during decline of the injury phase and at the initiation of the regeneration phase.
Furthermore, during regeneration phase, BRG-1-containing remodeler again predominates. Besides this, the catalytic ATPase subunit modulates chromatin structure that often regulates gene expression. There are reports suggesting that upregulation of BRG-1 is associated with senescence. On the contrary, downregulation of BRG-1 induces process of heterochromatin formation, which is associated with activation of retinoblastoma-like protein, P53- related pathway, and cell cycle control (Alessio et al., 2010; Squillaro et al., 2015). The recent findings suggest that alteration in BRG-1 level affect the expression of different chromatin proteins, including histones and nonhistone proteins (Alessio et al., 2010). BRG-1 is also proposed to act as a global regulatory element and regulate histone code both at the translational and posttranslational levels and thereby affect nuclear architecture (Squillaro et al., 2015). The present study was focused on liver regeneration, and our ChIP data analysis indicates that Brg-1 promoter is transcriptionally active during injury and regeneration phases, as H3acK9 is a marker for active transcription. Hence, our result also supports recent findings, suggesting BRG-1 is required for activation of gene promoter. Conversely, BRM-specific complexes are reported to be present only on the repressed promoters (Flowers et al., 2009; Squillaro et al., 2015). Hence, based on the present data and earlier reports, it can be hypothesized that BRG-1 might act as a global and important regulatory element for gene transcription during liver regeneration model.
Considering all these observations, we propose a model depicting interplay between chromatin interacting machineries in different phases of thioacetamide-induced liver injury and regeneration (Fig. 6). It would be interesting to investigate the above proposition further by performing promoter-specific immunoprecipitation (ChIP, ChIP-Seq analysis), to demonstrate differential occupancy of BRG-1- and BRM-containing SWI/SNF remodeling complexes.

Model proposed to depict the interplay of chromatin interacting machineries during different phases of thioacetamide-induced liver injury and regeneration.
Footnotes
Acknowledgments
Financial support from the Department of Science and Technology, Government of India PURSE grants, and Research & Development grant from the University of Delhi to Prof. M.M.C., for the present study, are highly acknowledged. Research fellowships from the University Grants Commission (UGC) and Council of Scientific and Industrial Research (CSIR) to S.S. and S.V., respectively are acknowledged with thanks. The authors highly acknowledged Dr. J.S. Purohit, Cluster Innovation Center (CIC), University of Delhi, for critical reviewing of the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.