
Abstract
Osteosarcoma is an aggressive tumor and the most common malignancy of the skeleton. Due to pulmonary metastasis, the 5-year survival rate is still unsatisfactory. It has been reported that SPRY4 intronic transcript 1 (SPRY4-IT1) promotes cell growth, invasion, and inhibits apoptosis in several cancers. However, the role of SPRY4-IT1 in osteosarcoma remains unclear. In the present study, we investigated the role of SPRY4-IT1 in osteosarcoma cells. Loss- and gain-of-function assays demonstrated that SPRY4-IT1 promoted cell proliferation, migration, and invasion in osteosarcoma. Moreover, SPRY4-IT1 induced epithelial–mesenchymal transition phenotype in osteosarcoma cells. Subsequent investigations revealed that SPRY4-IT1 promoted migration and invasion through association with Snail1 and regulating its stability. Based on these findings, the SPRY4-IT1/Snail1/E-cadherin pathway may play a crucial role in promoting osteosarcoma metastasis. Thus, SPRY4-IT1 may be a potential target for new therapies of osteosarcoma.
Introduction
O
Epithelial–mesenchymal transition (EMT) is an essential phenotypic conversion during embryonic development, tissue remodeling and healing, and tumor metastasis (Thiery et al., 2009). During EMT process, epithelial cells acquire fibroblast-like properties, which reduce its intercellular adhesion and increases cell motility. When cancer cells disseminate to distant sites of other organs, then they can return back to an epithelial phenotype through mesenchymal–epithelial transition. The loss of E-cadherin expression is a hallmark of EMT process (Nieto, 2009). Several transcription factors, including Snail, Twist, and ZEB family, have been implicated in the transcriptional repression of E-cadherin and the induction of EMT (Villarejo et al., 2014; Diaz-Lopez et al., 2015; Wang et al., 2015a). Although E-cadherin expression is often downregulated in osteosarcoma and closely related with poor prognosis (Yin et al., 2012; Yang et al., 2014), the molecular mechanism of EMT has not been completely understood.
Advances in genome sequencing technologies have enabled a large scale of transcripts study. The genomes encode an abundance of different types of noncoding RNAs (ncRNAs), such as microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circle RNAs, which are expected to constitute a complicated regulatory network that modulates different gene expressions. The lncRNAs are defined as ncRNAs longer than 200 nucleotides (nt) (Mattick and Rinn, 2015). Even though thousands of lncRNAs have been identified, few of them have been assessed for their biological function. The lncRNAs exhibit tissue-specific regulated expression patterns, and are often dysregulated in human cancers, including osteosarcoma (Zhao and Lin, 2015). Recent studies have reported that lncRNAs play critical roles in tumor initiation, progression, and metastasis through modulating cancer-related pathways (Hu et al., 2014; Yuan et al., 2014, 2015). Interestingly, lncRNAs regulate target genes at specific genomic loci (Wang et al., 2015b), modulate the stability of its binding protein (Hu et al., 2014), recruit chromatin-modifying complexes to their downstream genes (Zhang et al., 2015a, 2015b), and serve as competing endogenous RNAs by sponging with miRNAs (Tay et al., 2014; Lu et al., 2015). The lncRNA SPRY4 intronic transcript 1 (SPRY4-IT1), derived from an intron within SPRY4, was previously reported to be upregulated in many kinds of human cancers, including prostate cancer (Mouraviev et al., 2015), melanoma (Khaitan et al., 2011), glioma (Liu et al., 2015), and non-small-cell lung cancer (Sun et al., 2014). SPRY4-IT1 promoted cell growth, invasion, and inhibited apoptosis (Khaitan et al., 2011). However, the role of SPRY4-IT1 in osteosarcoma remains unclear.
In the present study, we investigated the role of SPRY4-IT1 in osteosarcoma cells. We found that SPRY4-IT1 promoted proliferation, migration, and invasion of osteosarcoma cells. SPRY4-IT1 stimulated migration and invasion through association with EMT-related protein SNAIL1.
Materials and Methods
Cell culture
The human osteosarcoma lines MG63 were purchased from the Cell Resource Center of the Institute of Basic Medical Sciences at the Chinese Academy of Medical Sciences. Cells were maintained in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 μg/mL penicillin G, and 100 U/mL streptomycin (Invitrogen).
Reagents
The following antibodies were used: rabbit anti-human Snail1 (Abcam), rabbit anti-human Snail2 (Abcam), rabbit anti-human ZEB1 (Abcam), rabbit anti-human ZEB2 (Abcam), rabbit anti-human Twist1 (Abcam), rabbit anti-human Twist2 (Abcam), mouse anti-human E-cadherin (Santa Cruz), mouse anti-human Vimentin (Santa Cruz), mouse anti-human N-cadherin (Santa Cruz), and mouse monoclonal antibody anti-β-actin (Proteintech).
RNA interference
shRNAs against SPRY4-IT1 and Snail1 were designed with MIT's siRNA designer (
SPRY4-IT1 overexpression
Full-length SPRY4-IT1 cDNA was cloned into pCDNA3.1 vector. Cells were transfected with the above plasmid. Stable clones were obtained by selection with G418.
Western blot
Cells were lysed in RIPA lysis buffer supplemented with cocktail protease inhibitor (Selleck). Total proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride (PVDF) membrane (Millipore). The PVDF membranes were incubated with primary antibodies and followed by incubation with secondary antibodies coupled to horseradish peroxidase (Jackson). Signals were visualized with a chemiluminescence system.
RNA extraction and quantitative real-time polymerase chain reaction assay
Total RNA was isolated using TRIzol (Invitrogen) according to the manufacturer's instructions. First-strand cDNA was generated using the reverse transcriptase (Transgene) and random primers. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed in the StepOne™ Real-Time PCR System using SYBR® Green (TaKaRa). Data were normalized to ACTB or to control samples. Primer sequences for the detected genes were as follows: CDH1-F: 5′-CTTCTGCTGATCCTGTCTGATG-3′, CDH1-R: 5′-TGCTGTGAAGGGAGATGTATTG-3′; CDH2-F: 5′-GGATGAAACGCCGGGATAAA-3′; CDH2-R: 5′-TCTTCTTCTCCTCCACCTTCTT-3′; VIM-F: 5′-CAGCTTTCAAGTGCCTTTCTG-3′, VIM-R: 5′-CTTGTAGGAGTGTCGGTTGTT-3′; Snail1-F: 5′-CCTTCGTCCTTCTCCTCTACTT-3′, Snail1-R: 5′-TTCGAGCCTGGAGATCCTT-3′; and ACTB-F: 5′-CACCAGGCACCCAGTTTAAT-3′, ACTB-R: 5′-AGTCTCTGCTCTCTCTTCCTATG-3′.
RNA immunoprecipitation
RNA immunoprecipitation (RIP) assays were performed by using the EZ-Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to the manufacturer's instructions. RIP products were analyzed by qRT-PCR. Five micrograms of Snail1 antibodies were used for per RIP reaction.
3-2,5-Diphenyl tetrazolium bromide assays
Cells were seeded at a density of 1 × 103 cells per well in 96-well plates, the viability of the cells was assessed by the 3-2,5-diphenyl tetrazolium bromide (MTT) assay (Dojindo) for 4 days.
RNA pull-down
RNA pull-down was performed as described (Yang et al., 2011). In vitro biotin-labeled RNAs (SPRY4-IT1, its antisense RNA) were transcribed with the biotin RNA Labeling Mix (Roche) and T7 RNA polymerase (Roche) treated with RNase-free DNase I (Promega) and purified with the RNeasy Mini Kit (QIAGEN). Biotinylated RNA was incubated with MG63 cell nuclear extracts, and the retrieved protein was detected by the standard western blotting.
Migration and invasion assays
Migration and invasion assays were performed as previously described (Tiwari et al., 2013). In brief, 105 cells were seeded in FBS-free DMEM in the upper chamber of a 24-well transwell migration or invasion insert (BD Bioscience). The lower chamber was filled with medium containing 10% FBS. After 24 h of incubation at 37°C, cells in the upper chamber were removed with a cotton swap, and the cells that had traversed the membrane were fixed in 4% paraformaldehyde and stained by crystal violet.
Statistical analysis
Data were analyzed with a two-tailed Student's t test using the SPSS 13.0 software and GraphPad Prism 6. p < 0.05 was considered statistically significant.
Results
SPRY4-IT1 promotes cell proliferation, migration, and invasion
To investigate the biological function of SPRY4-IT1 in osteosarcoma, we performed gain- and loss-of-function studies in osteosarcoma cells. We constructed stable SPRY4-IT1 overexpressed and knockdown MG63 cells (Fig. 1A). By performing MTT assay, we found that SPRY4-IT1 overexpression increased the proliferation of MG63 cells compared with control cells (Fig. 1B). In contrast, knockdown of SPRY4-IT1 significantly decreased the proliferative capacity of MG63 cells (Fig. 1C). To evaluate the role of SPRY4-IT1 in the metastasis of osteosarcoma, migration and invasion assays were performed. Overexpression of SPRY4-IT1 significantly promoted the migration and invasion in MG63 cells (Fig. 1D). In contrast, knockdown of SPRY4-IT1 dramatically reduced cell migration and invasion of MG63 cells (Fig. 1E). These data suggested that SPRY4-IT1 promoted cell proliferation, migration, and invasion in osteosarcoma cells.
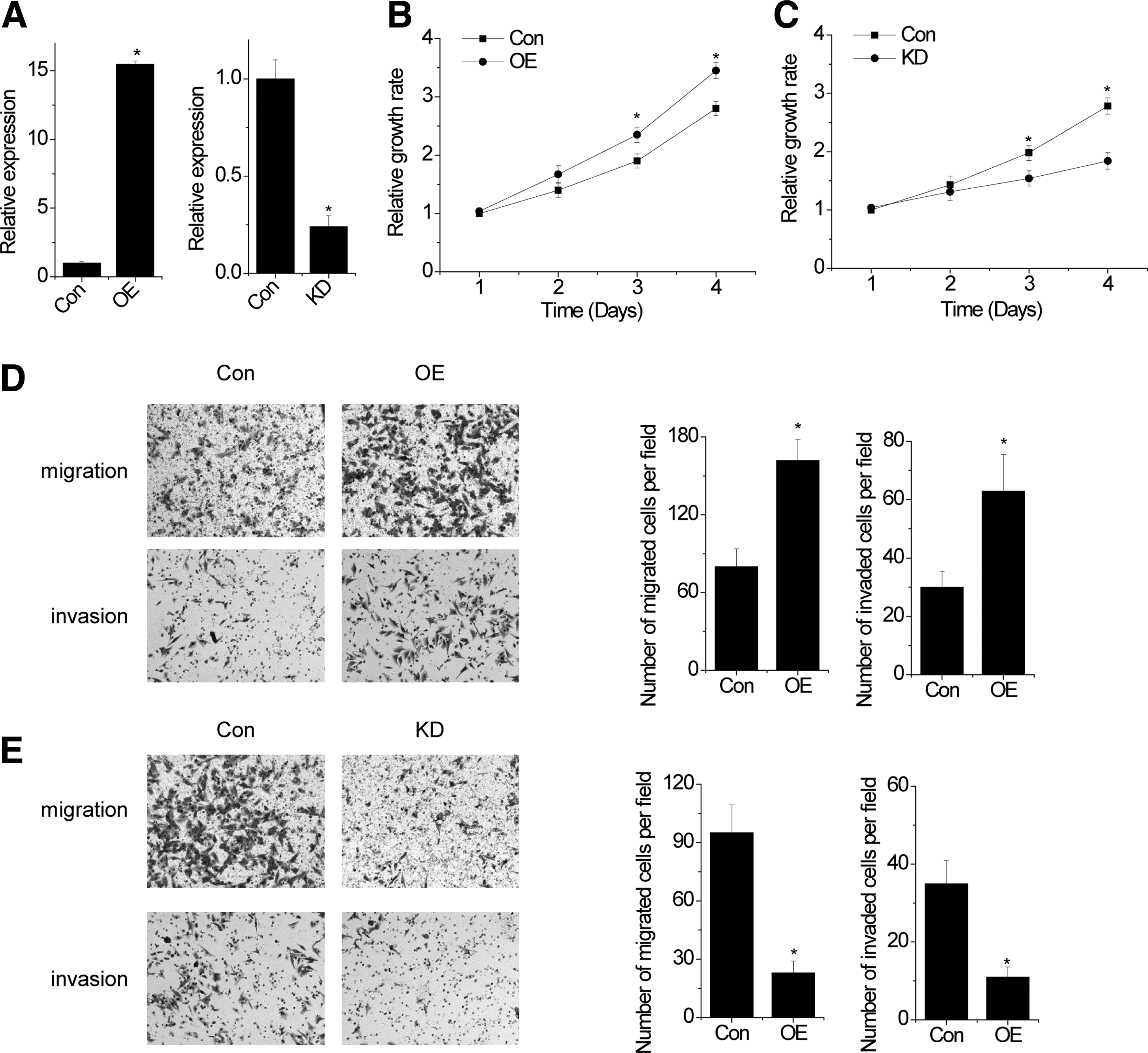
SPRY4-IT1 promotes cell proliferation, migration, and invasion.
SPRY4-IT1 induces EMT
EMT plays an important role in metastasis. Interestingly, we found that overexpression of SPRY4-IT1 in MG63 cells resulted in the reduced expression of epithelial markers (E-cadherin) and increased expression of mesenchymal markers (N-cadherin and vimentin), as evidenced by western blot and qRT-PCR assays (Fig. 2A, B). Conversely, epithelial markers were markedly increased and mesenchymal markers were significantly decreased in SPRY4-IT1 knockdown cells (Fig. 2C, D). These findings suggested that SPRY4-IT1 induced EMT in osteosarcoma cells.
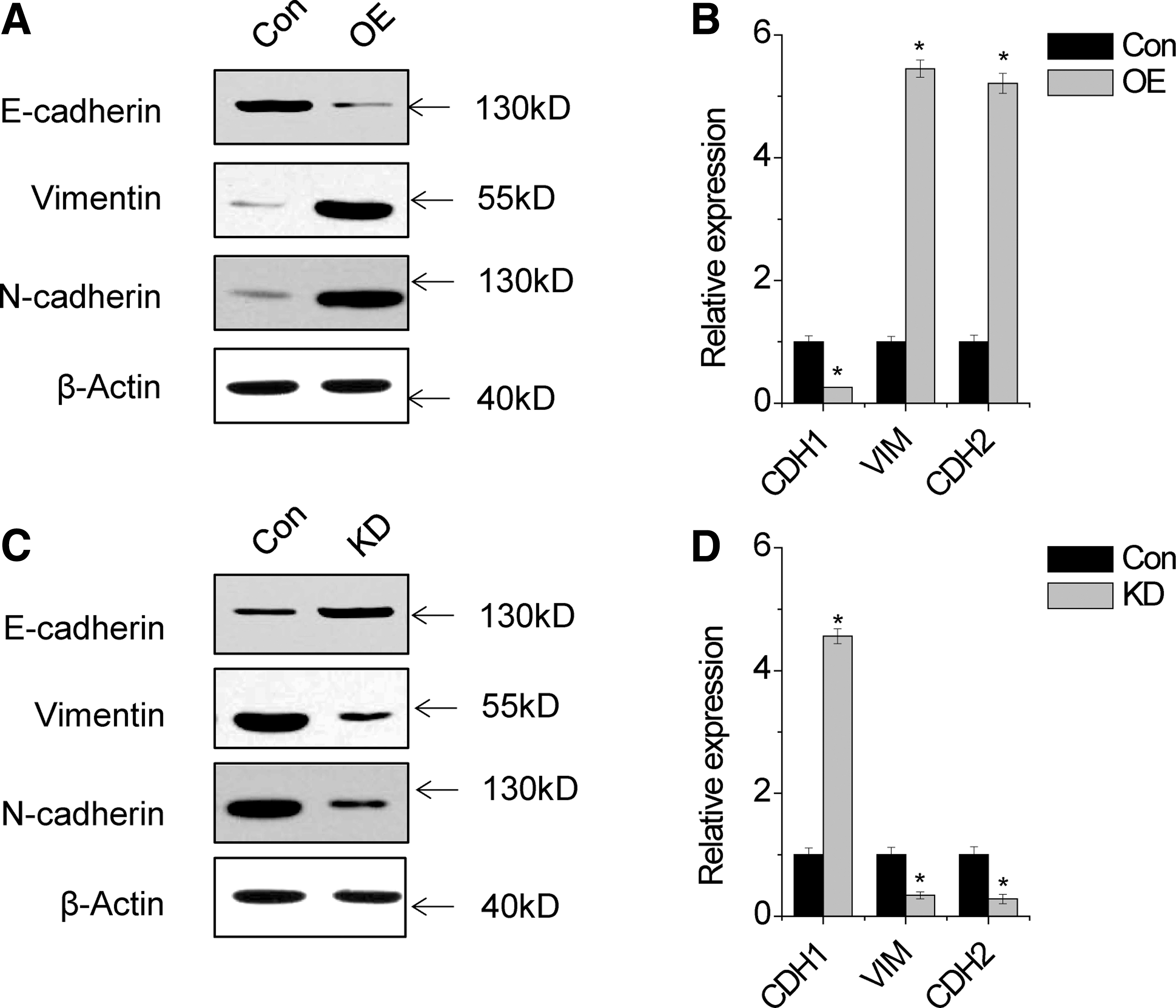
SPRY4-IT1 induces EMT.
SPRY4-IT1 associates with Snail1 and regulates its stability
Functional loss of E-cadherin, induced by Snail, ZEB, and Twist family, is considered a hallmark of EMT process (Loeffler and Wolf, 2015). We determined whether SPRY4-IT1 is associated with these repressors. RIP analysis showed that SPRY4-IT1 was significantly pulled down by Snail1 antibody compared with the nonspecific immunoglobulin G control antibody, Snail2, ZEB1, ZEB2, Twist1, or Twist2 antibody (Fig. 3A). Furthermore, we performed RNA pull-down assay to validate the association between SPRY4-IT1 and Snail1. Consistent with the RIP assay, only Snail1 could be pulled down by biotinylated SPRY4-IT1 RNA (Fig. 3B). Next, we performed deletion-mapping experiments to determine whether Snail1 would associate with a specific region of SPRY4-IT1 (Fig. 3B). These analyses identified a 266-nt region at the 5′ end of SPRY4-IT1 required for the association with Snail1. Taken together, we demonstrated a specific association between Snail1 and SPRY4-IT1.
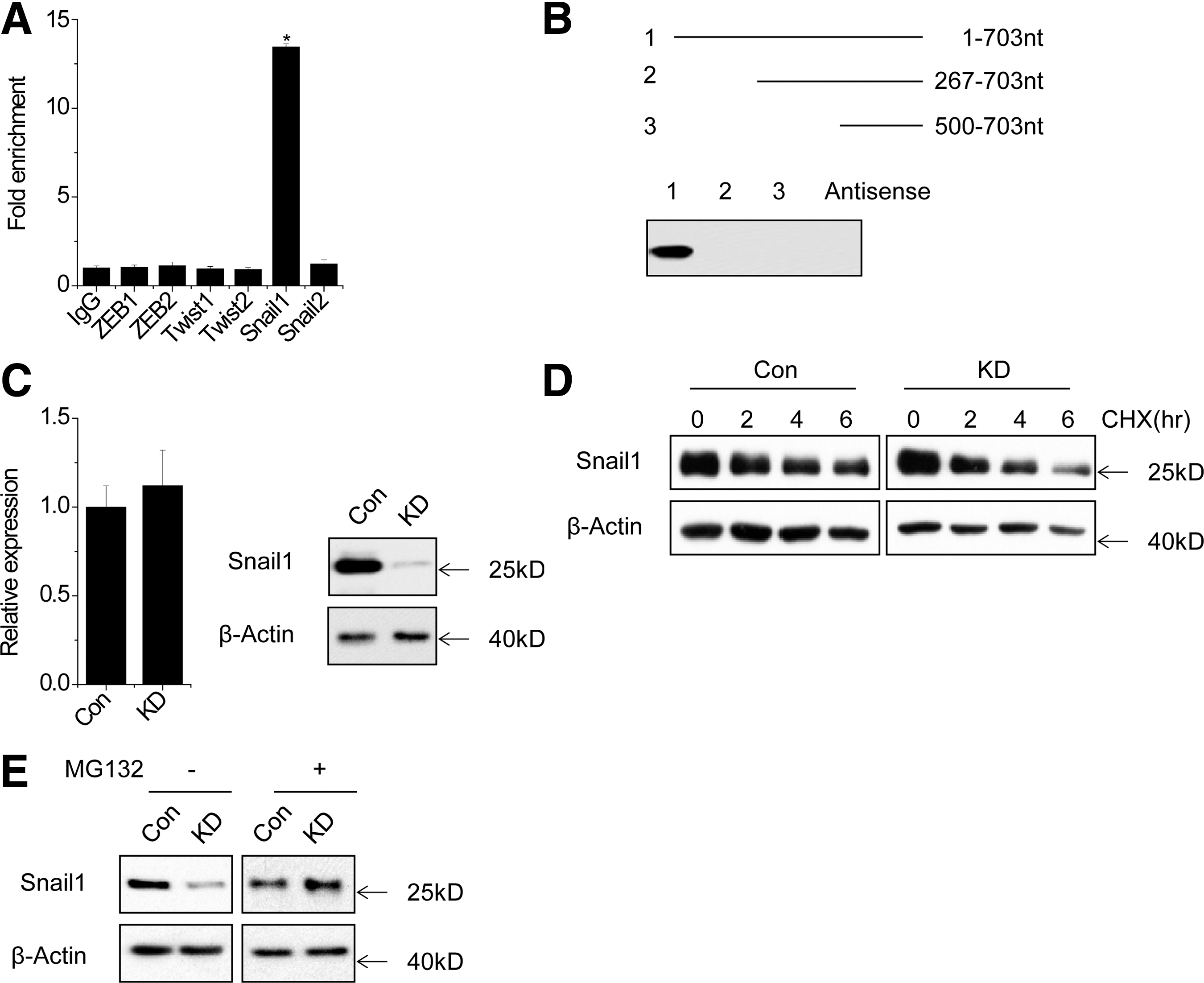
SPRY4-IT1 associates with Snail1 and regulates its stability.
Next, we explored the functional association of Snail1–SPRY4-IT1interaction. SPRY4-IT1 shRNAs significantly reduced the protein level, but not the mRNA level of Snail1 (Fig. 3C). This result suggests that the Snail1–SPRY4-IT1 association may help stabilize Snail1 protein. To further explore the mechanism of SPRY4-IT1-mediated Snail1 regulation, we treated control and SPRY4-IT1 knockdown MG63 cells with cycloheximide and analyzed the stability of Snail1. The half-life of Snail1 was much shorter in SPRY4-IT1 knockdown MG63 cells than in controls (Fig. 3D). Consistent with this result, when MG132 was used to inhibit proteasome degradation, the Snail1 protein expression in SPRY4-IT1 knockdown cells was markedly increased and reached a level that was comparable to that in control cells (Fig. 3E). In aggregate, these data suggested that SPRY4-IT1 is important for regulating Snail1 protein stability.
SPRY4-IT1 promotes migration and invasion through association with Snail1
Finally, we determined whether SPRY4-IT1 promotes migration and invasion through regulation with Snail1. SPRY4-IT1 overexpression promoted migration and invasion and decreased E-cadherin expression in MG63 cells, whereas the inhibition of Snail1 expression significantly attenuated this phenomenon induced by SPRY4-IT1 (Fig. 4A, B). Thus, Snail1 is critical for SPRY4-IT1-induced metastasis.

SPRY4-IT1 promotes migration and invasion through regulation Snail1.
Discussion
Recently, several lncRNAs have been characterized and many studies show that they play a central role in cancer occurrence and progression, which provide a new insight into the biology of human cancers. However, lncRNAs involved in osteosarcoma are still an emerging field, with only a handful of lncRNAs known to be functional in osteosarcoma development, such as HULC, MALAT1, and EWSAT1 (Marques Howarth et al., 2014; Dong et al., 2015; Sun et al., 2015). In the present study, for the first time, we found that the overexpression of SPRY4-IT1 significantly promoted cell proliferation, migration, and invasion. Conversely, knockdown of SPRY4-IT1 inhibited cell proliferation, migration, and invasion. These findings suggest that SPRY4-IT1 plays a direct role in the regulation of metastasis and progression of osteosarcoma, and SPRY4-IT1 may be a potential novel marker for prognosis and progression of osteosarcoma.
Recurrence and metastasis is the common lethal cause after surgery in osteosarcoma (Berner et al., 2015). Thus, it is essential to investigate the mechanisms underlying osteosarcoma metastasis. EMT plays an important role in migration and invasion. The EMT promotes dissemination of tumor cells, which is controlled by several transcription factors, including Twist, Snail, and ZEB family (Sanchez-Tillo et al., 2012). E-cadherin is directly repressed by Snail1, which in turn induces EMT in epithelial tumor cells (Dong et al., 2012). Several reports support a close association between SPRY4-IT1 expression and tumor metastasis (Liu et al., 2015; Peng et al., 2015). SPRY4-IT1 also induces EMT (Liu et al., 2015). However, the mechanism by which SPRY4-IT1 promotes metastasis through EMT in osteosarcoma remains unclear. This study was the first to demonstrate that SPRY4-IT1 is associated with Snail1 and regulated its stability through posttranslational modification. Inhibition of Snail1 expression significantly reduced SPRY4-IT1-induced migration and invasion. Thus, experimental evidences indicate that the SPRY4-IT1/Snail1/E-cadherin pathway may play a crucial role in promoting osteosarcoma metastasis.
In conclusion, our study demonstrates that SPRY4-IT1 promotes osteosarcoma proliferation, migration, and invasion. SPRY4-IT1 induces EMT process through association with Snail1 and regulates its stability. Thus, SPRY4-IT1 may be a potential target for new therapies of osteosarcoma.
Footnotes
Acknowledgment
This study was financially supported by the Hubei Provincial Department of Education and Natural Science Foundation (B20121304).
Disclosure Statement
No competing financial interests exist.