
Abstract
Inflammation is thought to have a major role in the pathogenesis of disc degeneration. Studies have shown that nucleus pulposus cells (NPCs) respond to one or two specific cytokines by regulating cell proliferation or matrix synthesis. However, the effects of a cocktail of factors secreted by degenerated disc cells on transplanted exogenous healthy NPCs remain unknown. Concentrations of multiple cytokines in degenerated disc tissue-conditioned medium (dCM) were measured using enzyme-linked immunosorbent assay (ELISA). 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay and Ki67 immunofluorescence staining were used to evaluate the proliferation of cells in dCM. The function of exogenous NPCs cultured in dCM was evaluated by examining catabolic markers (ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13), anabolic markers (TIMP-1, TIMP-2, and TIMP-3), and the extracellular matrix protein—aggrecan (ACAN) and collagen II (COL2)—expression with real time polymerase chain reaction (RT-PCR). Mitogen-activated protein kinase (MAPK) and nuclear factor-kappa B (NF-κB) pathway activation was observed using Western blotting. Finally, we examined the role of transforming growth factor (TGF)-β1 in reducing dCM-mediated exogenous NPC dysfunction. Levels of tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-1α, IL-2, IL-4, IL-6, IL-8, IL-10, IL-17, interferon-γ (IFN-γ), and prostaglandin E2 (PGE2) were higher and TGF-β1 levels were lower in dCM compared with the control medium. Treatment with dCM increased the proliferation of healthy NPCs. NPCs exhibited significantly higher expression of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 and decreased TIMP-2, ACAN, and COL2 expression in the dCM group in a dose- and time-dependent manner. Treatment with dCM moderately increased TIMP-1 expression and had no effect on TIMP-3 mRNA levels. The MAPK and NF-κB pathways were implicated in dCM-mediated responses of healthy NPCs. TGF-β1 partially reversed the dCM-mediated NPC dysfunction. Increased levels of inflammatory factors and decreased TGF-β1 levels in dCM suggest an inflammatory environment in degenerated disc tissue. The catabolic effect of dCM on human healthy NPCs is mediated by MAPK and NF-κB pathways and can be reduced by TGF-β1.
Introduction
D
Secreted cytokines associated with IVDD include tumor necrosis factor (TNF)-α, interleukin (IL)-1, IL-2, IL-4, IL-6, IL-8, IL-10, IL-17, interferon-γ (IFN-γ), various chemokines, and prostaglandin E2 (PGE2) (Le Maitre et al., 2005, 2007; Seguin et al., 2005; Shamji et al., 2010; Cuellar et al., 2013). Most of these cytokines are implicated in disc degeneration by downregulating ECM protein transcription and upregulating ECM-degrading enzyme expression (Studer et al., 2008; Risbud and Shapiro, 2014). The aforementioned molecular changes within the nucleus pulposus (NP) and annulus fibrosus (AF) in the early stages of IVDD, followed by structural deficits in the NP and AF and the formation of tears and clefts in these tissues, could enable immune cell activation and infiltration. Furthermore, nucleus pulposus cells (NPCs) and infiltrating immune cells continue to secrete cytokines and neurogenic factors, which reinforce cytokine-mediated IVDD and neurovascular ingrowth into the IVD (Risbud and Shapiro, 2014).
Various biological therapies have been developed in attempts to prevent or reverse the disc degenerative process with growth factors, cell transplantation, and gene therapy (Cai et al., 2015; Sakai and Grad, 2015). Among these methods, increasing the number of functional NPCs can be instrumental to rebuilding IVD structure and function (Leung et al., 2006). Advances in regenerative medicine and cell-based therapies, particularly mesenchymal stem cell and NPC transplantation, have led to numerous reports and clinical trials on the utility of these biological therapies to treat degenerative spinal conditions (Oehme et al., 2015).
The most important consideration after ensuring that cells remain within the disc is their survival and function in the inflammatory environment. Many studies have assessed cell viability and survival within discs, with conflicting results (Oehme et al., 2015). It is likely that factors released from resident disc cells reduce NPC function through certain hostile inflammatory factors, especially in severely degenerated discs. However, it is not fully known how resident disc cells influence the expression of ECM-degrading enzymes and their inhibitors in transplanted seed cells within a paracrine framework.
The increased expression and synthesis of inflammatory factors and ECM-degrading enzymes, such as a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTSs) and matrix metallopeptidases (MMPs), are mediated by specific signal transduction, in particular the nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK)-mediated pathways. NF-κB and MAPK were identified as major regulators of inflammation and catabolism in several musculoskeletal disorders (e.g., osteoarthritis), and growing evidence supports the importance of these signaling pathways in degenerated disc disease. However, the function of NF-κB- and MAPK-mediated pathways on the survival and function of transplanted healthy NPCs in an environment driven by paracrine effects of resident degenerated NPCs is not well understood.
Transforming growth factor (TGF)-β1 is a multifunctional factor that regulates cell growth, adhesion, and differentiation in a wide variety of cell types (Massague, 2008). In addition, TGF-β1 is a strong immune suppressor (Kulkarni et al., 1993). Previous studies examined TGF-β1 content in normal and degenerated human IVDs using immunohistochemistry and revealed that TGF-β1 expression in normal disc tissue was significantly higher than in degenerated disc tissue (Tolonen et al., 2001; Burke et al., 2003; Peng et al., 2006). A recent study found that mesenchymal stem cells promoted the amelioration of disc degradation by inducing TGF-β1 expression in the NP (Yang et al., 2015a). These findings suggest that TGF-β1 may have a role in disc aging and degeneration; however, it is not well known whether TGF-β1 could promote the function of transplanted healthy NPCs in degenerated disc tissue.
Earlier studies have shown that NPCs respond to one or two specific cytokines by regulating cell proliferation or matrix synthesis (Mavrogonatou et al., 2014; Wang et al., 2014a, 2015); however, the effects of a cocktail of cytokines secreted by degenerated NPCs on transplanted healthy NPCs are unknown.
In this study, we investigated the survival and function of transplanted healthy NPCs in degenerated NP tissue-conditioned medium (dCM) representative of the paracrine function of degenerated NPCs. The most important function of transplanted NPCs is the synthesis of ECM, including ACAN and COL2, to promote disc regeneration. Therefore, we evaluated NPC function by examining the expression of ADAMTSs (ADAMTS-4 and ADAMTS-5), MMPs (MMP-1, MMP-3, and MMP-13), tissue inhibitor of metalloproteinases (TIMP-1, TIMP-2, and TIMP-3), and ECM proteins (ACAN and COL2). Furthermore, we examined MAPK and NF-κB signaling pathway activation in transplanted healthy NPCs to explore the underlying mechanisms. Finally, we examined whether treatment of transplanted healthy NPCs with TGF-β1 could protect them from an inflammatory environment. The structure of this article is shown in the following schematic (Fig. 1).

The structure of this study.
Methods
Patient samples
Degenerated human IVDs were obtained from patients receiving posterior lumbar fusion (Table 1). In six donors, disc degeneration was measured as grade V according to the Pfirrmann scale of IVDD (Pfirrmann et al., 2001). Another six patients with lumbosacral spondylolysis or acute spine damage, but without IVDD (grade I on the Pfirrmann scale), were included as healthy cases. IVDs were collected and divided into several segments containing only NP tissue and kept in sterile tubes. All of the tissues used in this study were obtained in compliance with Chinese law and the guidelines of the Helsinki Declaration. All experiments were conducted with the approval of the ethics committee of Zhongda Hospital, affiliated to Southeast University.
TLIF, transforaminal lumbar interbody fusion.
Isolation and culture of NPCs
NP and AF tissues were defined and identified during the surgical and laboratorial procedures. During surgery, a small hole was cut in the AF, and the white and tough AF tissue covering at the hole was removed clearly by a nucleus pulposus clamp. Material from inside the disc, most of which was soft NP tissue, was obtained and kept in sterile tubes. Other fragments of the disc after using a vertebral endplate clear reamer should not be collected due to a mixture of endplate tissue. NP tissue kept in sterile tubes was brought into a clean bench and the remaining AF tissue was picked out under a microscope. Following the above procedures, NP tissue was finely minced and digested using 0.2% collagenase II for 6 h before passing sequentially through a cell strainer with pore size of 70 μm (BD Biosciences, United Kingdom). Even if there was a little bit of mixed AF tissue, collagen I-rich AF tissue was left after incubation in 0.2% collagenase II for 6 h and separated from NPCs.
The isolated NPCs were harvested and grown in Dulbecco's modified Eagle's medium with low glucose (DMEM-LG; Gibco), supplemented with 10% (v/v) fetal bovine serum (FBS; Wisent, Inc., Canada), and 1% penicillin–streptomycin (Gibco) and incubated at 37°C in a humidified atmosphere with 5% CO2. The medium was replenished every 3 days. When cells reached 80–90% confluence, they were subcultured using 0.25% trypsin/0.02% ethylenediaminetetraacetate (EDTA; Gibco) and designated as passage 1 (P1). P1 cells were used in subsequent experiments. NPCs collected from donors 1–6 were pooled as healthy NPCs.
For each sample, 5 × 104 P1 cells were characterized by means of flow cytometry analysis. Cells were incubated with appropriate phycoerythrin (PE)-conjugated antibodies to test the expression of hematopoietic and NPC markers: CD34 and CD24. After 30 min at 4°C, cells were washed once with phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde, and analyzed using a flow cytometer and cell quest software (BD Pharmingen). Immunofluorescence staining was used to test the specific markers of NPCs. P1 NPCs were washed with PBS and fixed in 4% paraformaldehyde for 25 min at room temperature. Cells were washed three times with PBS and permeabilized with 0.2% Triton X-100 in PBS for 15 min. Nonspecific binding was blocked by incubating cells in 5% bovine serum albumin (BSA)/PBS for 60 min. Then, cells were incubated overnight at 4°C with rabbit polyclonal anti-human CD24 and COL2 antibody (1:400; Abcam) in 1% BSA/PBS. The cells were incubated in the dark for 2 h with Alexa Fluor 594-conjugated donkey anti-rabbit IgG (1:800; Abcam) at room temperature. Nuclei were counterstained with 4′6-diamidino-2-phenylindole (DAPI) at 37°C for 2 min. Fluorescence was visualized using a laser scanning confocal microscope (FV2000; Olympus, Japan). Isotype-matched rabbit immunoglobulins were used as control.
Preparation of NP tissue-conditioned medium
NP tissue was cultured for 4 days in DMEM-LG with 1% penicillin–streptomycin (1 g tissue/30 mL) and incubated at 37°C in a humidified atmosphere with 5% CO2. After 4 days, the NP tissue was removed and the conditioned medium (CM) was filtered through a 70-μm cell strainer and the filtrate was centrifuged at 500 g (15 min, 4°C) to remove cells and debris. Then, the supernatant containing cell-free CM was concentrated by centrifuging at 4000 g (45 min, 4°C) in a centrifugal ultrafilter with a molecular weight cutoff of 3 kDa (Millipore, The Netherlands). Concentrated CM was used in the experiments after dilution with fresh culture medium at concentrations equivalent to 100%, 200%, and 300% of the initial protein content. NP tissue obtained from degenerated disc (Table 1, donors 1, 7–11) was cultured as an experimental group (degenerated CM [dCM]). Healthy NP tissue (Pfirrmann grade I) obtained from six donors (Table 1, donors 1–6) was cultured as a control (nondegenerated CM [nCM]).
Cell viability
The cultured NP tissue was examined for viability after culturing for 4 days in DMEM-LG using live/dead cell staining. After culturing for 4 days, the NP tissue samples were rinsed twice with warm PBS and incubated for 30 min in serum-free DMEM containing 2 mM calcein acetoxymethyl ester (calcein AM; Sigma-Aldrich) and 1 μg/mL propidium iodide (PI; Sigma-Aldrich). The fluorochrome-labeled samples were rinsed with PBS, cryosectioned, and then examined under a fluorescence microscope (Olympus) fitted with the appropriate excitation and emission filters to detect live (green fluorescent) and dead (red fluorescent) cells.
Enzyme-linked immunosorbent assays
Concentrations of TNF-α, IL-1β, IL-1α, IL-2, IL-4, IL-6, IL-8, IL-10, IL-17, IFN-γ, PGE2, and TGF-β1 in nCM and dCM were measured using commercially available enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Inc.) following the manufacturer's protocols.
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay
P1 cells were plated in 96-well cell culture plates at a density of 1000 cells/well and cultured in a humidified incubator. The 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay was used to measure the number of viable cells. Cells treated with 300% nCM and 300% dCM were divided into five groups assigned to different time points (days 1–5). At each time point, 20 μL of 5 mg/mL MTT solution (Sigma-Aldrich) was added to each well of the corresponding group and incubated at 37°C for 4 h. The medium containing MTT was aspirated and 150 μL of dimethyl sulfoxide (DMSO; Sigma) was added to each well and incubated at 37°C for 30 min. Finally, the number of viable cells was determined by measuring the optical density at 492 nm using a microplate reader (Thermo Scientific).
Ki67 immunofluorescence staining
After culturing in 300% nCM, 300% dCM, or in standard culture condition (DMEM with 10% FBS) for 3 days, immunofluorescence staining was used to test the expression of Ki67 in NPCs. The detailed procedure was described above. The rabbit polyclonal anti-human Ki67 antibody (1:400; Abcam) and Alexa Fluor 488-conjugated donkey anti-rabbit IgG (1:800; Abcam) were used in this experiment.
TGF-β1 treatment
Healthy NPCs were seeded into six-well plates (Corning Life Sciences, NY) at a concentration of 5000 cells/cm2. Cells were preincubated in culture medium with 5 ng/mL recombinant human TGF-β1 (Abcam) for 24 h and then cultured in 300% dCM, supplemented with 5 ng/mL TGF-β1 (Tekari et al., 2015).
Inhibition of NF-κB and MAPK pathways
For the inhibitor treatment experiments, cells were pretreated with inhibitors for NF-κB (pyrrolidinedithiocarbamate ammonium [PDTC], 100 μmol/L), p38 (SB203580 [SB], 10 μmol/L), ERK1/2 (U0126, 10 μmol/L), or JNK (SP600125 [SP], 10 μmol/L) for 30 min before the dCM treatment. In addition, the effect of inhibitors on untreated cells was observed. All of the inhibitors were obtained from Abcam.
Real-time RT-PCR
After culturing in the above different conditions for 3 days, NPCs were washed with ice-cold PBS and total RNA was isolated, measured, and used to immediately synthesize cDNA. Real-time quantitative PCR using gene-specific primers (Table 2) was performed using the SYBR Green method, as described previously. Reactions were conducted using a 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA) in triplicate in 96-well plates with final volumes of 20 μL under standard conditions. The reaction mixture contained 10 μL of 2× SYBR Green Master Mix (TaKaRa, Japan), 0.4 μL of 10 μM forward primer, 0.4 μL of 10 μM reverse primer, 0.4 μL of 50× ROX reference dye, 6.8 μL of water, and 2 μL of 5 ng/μL cDNA. The 2−ΔCt and 2−ΔΔCt methods were used to calculate the relative expression levels of target genes. The housekeeping gene GAPDH was used as a reference to normalize PCR data, expressed as the ratio of the gene of interest to GAPDH.
Western blot analysis
After culturing in 300% nCM and 300% dCM for 0, 0.5, 1, and 6 h, NPCs were rinsed with ice-cold PBS and lysed in lysis buffer containing a mixture of protease inhibitors. Total protein was extracted from six samples and the protein content was quantified using a bicinchoninic acid quantification kit (Thermo Scientific). Proteins were resolved using sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred onto polyvinylidene fluoride membranes (Millipore), and blocked for 1 h with 5% (w/v) BSA in Tris-buffered saline with 0.1% Tween. Then, membranes were incubated with rabbit anti-human ERK1/2 (1:1000), phospho-ERK1/2 (Thr202/Tyr204) (1:2000), p38 MAPK (1:1000), phospho-p38 MAPK (Thr180/Tyr182) (1:1000), SAPK/JNK (1:1000), phospho-SAPK/JNK (Thr183/Tyr185) (1:1000), NF-κB p65 (1:1000), phospho-NF-κB p65 (Ser536) (1:1000), and α-tubulin (1:3000) overnight at 4°C with gentle shaking. All of the antibodies were obtained from Cell Signaling Technology. Next, the membranes were incubated with secondary antibodies coupled to horseradish peroxidase (goat anti-rabbit IgG, 1:2500; KeyGEN, China) for 1 h at room temperature. Protein was observed using the enhanced chemiluminescence method following the manufacturer's instructions (Millipore). Immunoblot band density was quantified using a Bio-Rad calibrated densitometer.
Statistical analyses
Statistical analyses were performed using SPSS ver. 20.0. All values are expressed as mean ± standard deviation. Groups were compared using the one-way analysis of variance and unpaired Student's t-test, and multiple comparison tests were conducted using Dunnett's test. p < 0.05 was considered statistically significant.
Results
Primary cultured human NPC morphology
Healthy NPCs were spindle shaped or polygonal, with multiple cytoplasmic processes and granular cytoplasms, similar to chondrocytes. NPCs isolated from degenerated discs exhibited a stellate morphology, with many cells extending several branching cell processes (Fig. 2). Flow cytometry analysis showed that the healthy and degenerated NPCs expressed high percentage of positivity for CD24, which was consistent with the previous study (Navone et al., 2012) (Fig. 3A), while the isolated cells expressed a lower positivity fraction for CD34, which was a hematopoietic marker. In addition, immunofluorescence staining showed that the isolated cells in both healthy and degenerated groups expressed characteristic marker of NPCs, CD24, and COL2 (Fig. 3B).
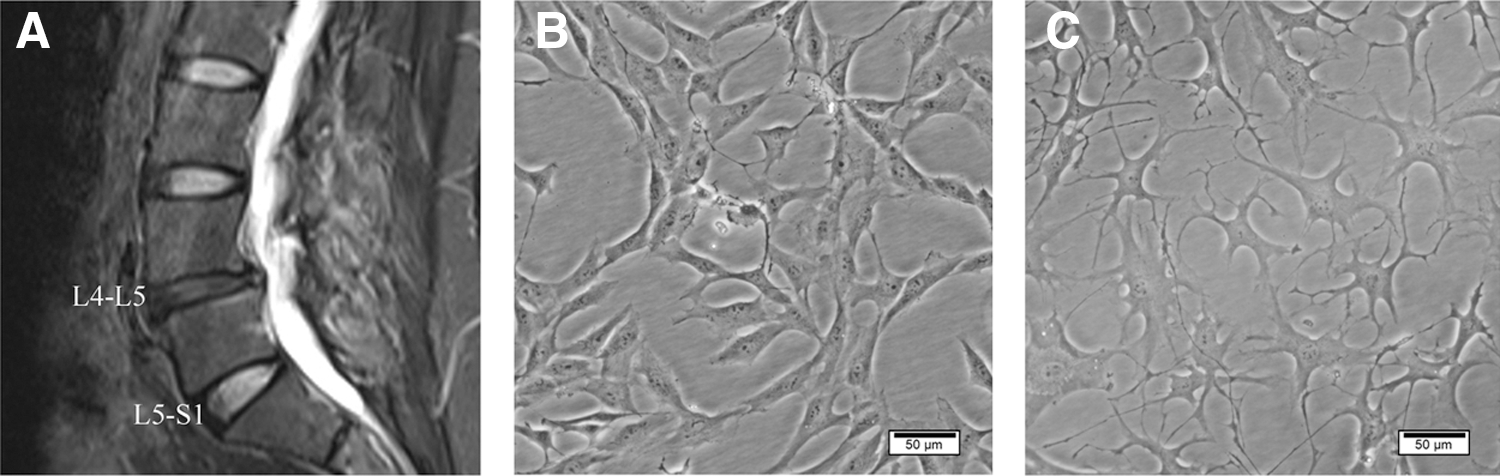
Morphology of primary cultured human NPCs.
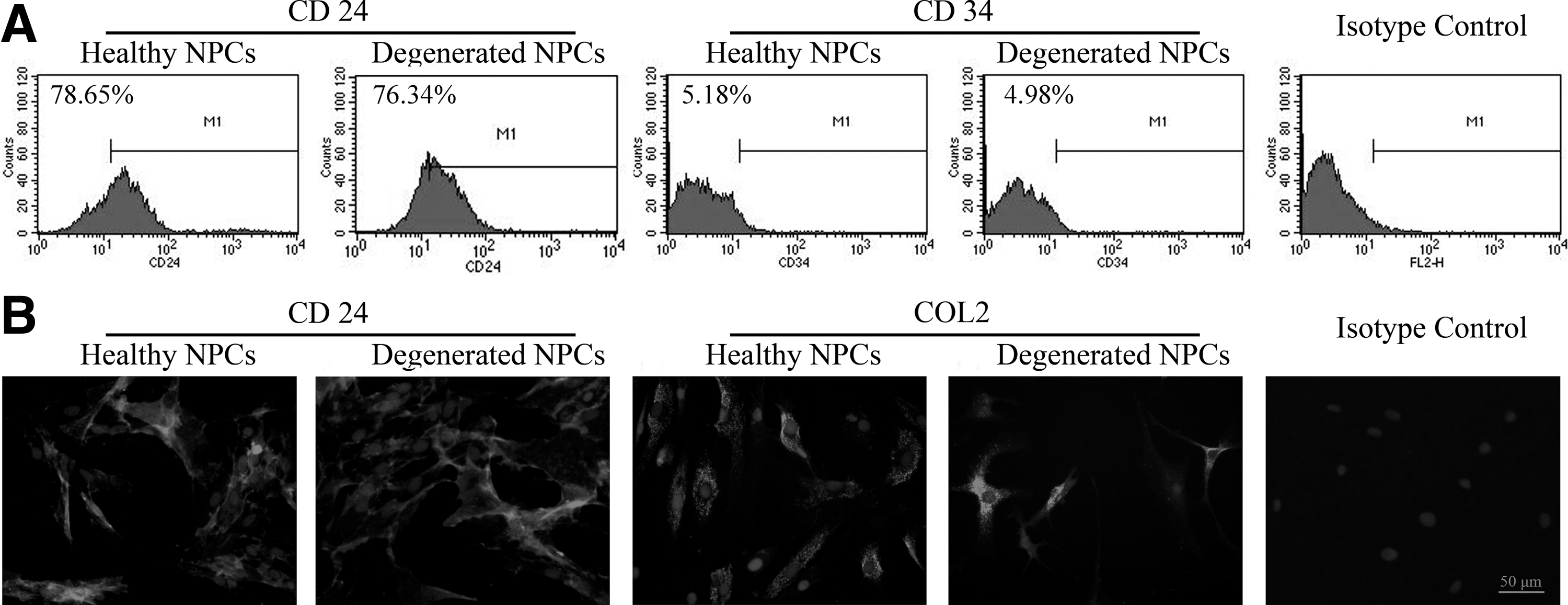
Identification of primary cultured human NPCs.
Live/dead assay
The live/dead assay allowed for direct observation of the proportion of living and dead cells (Fig. 4). The results showed that cell survival was maintained with >95% viable cells in the NP throughout the culture period (culturing for 4 days, 97.46% ± 1.6%, n = 6).
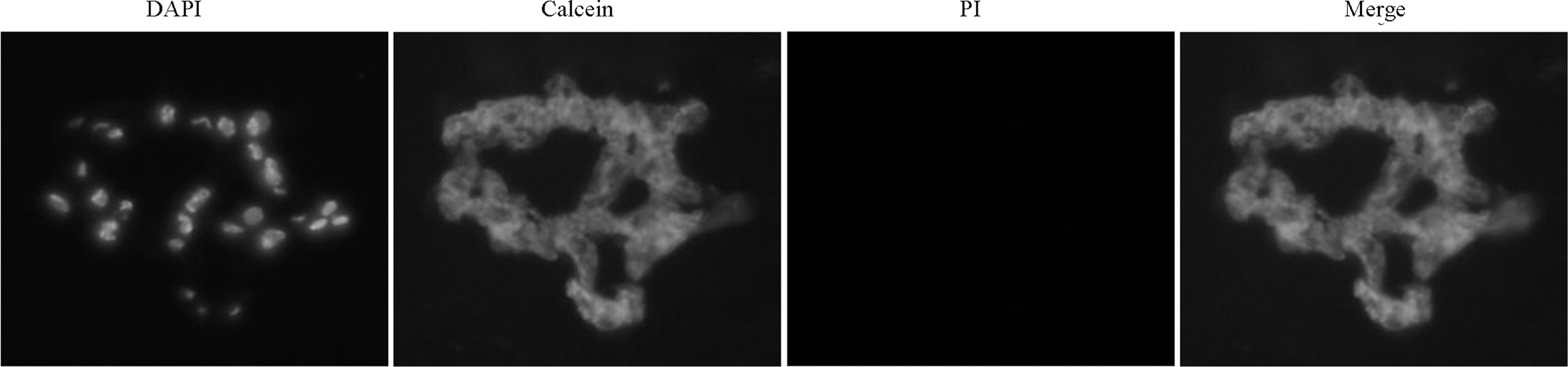
Live/dead assay. Cell viability of NP tissue cultures was examined after 4 days of culture in DMEM-LG using live/dead cell staining. DAPI, nucleus; calcein, live cells; PI, dead cells. Based on the results, cells survived over the 4 days of NP tissue culture in DMEM-LG. DAPI, 4′6-diamidino-2-phenylindole; DMEM-LG, Dulbecco's modified Eagle's medium with low glucose; PI, propidium iodide.
Quantization of inflammatory mediators in dCM
Table 3 summarizes the statistical comparison between dCM and nCM. dCM had increased levels of TNF-α, IL-1β, IL-1α, IL-2, IL-4, IL-6, IL-8, IL-10, IL-17, IFN-γ, and PGE2 and decreased levels of TGF-β1 compared with the nCM group. IL-4, IL-6, IL-8, IFN-γ, and PGE2 in nCM and TGF-β1 in dCM were below the detection limit of the system, suggesting that they were either expressed at extremely low levels or not expressed at all.
dCM, degenerated conditioned medium; IFN-γ, interferon-γ; IL, interleukin; nCM, nondegenerated conditioned medium; PGE2, prostaglandin E2; SD, standard deviation; TGF, transforming growth factor.
The effect of dCM on NPC proliferation
The effect of dCM on the proliferation of healthy NPCs was evaluated using the MTT assay. Healthy NPC proliferation was significantly stimulated by 300% dCM in a time-dependent manner over 5 days (Fig. 5A). To further validate the role of dCM in healthy NPC proliferation, we conducted Ki67 immunofluorescence staining. Compared with the 300% nCM group, dCM treatment increased the percentage of Ki67-positive NPCs (Fig. 5B, C), indicating that dCM had a positive effect on healthy NPC proliferation.
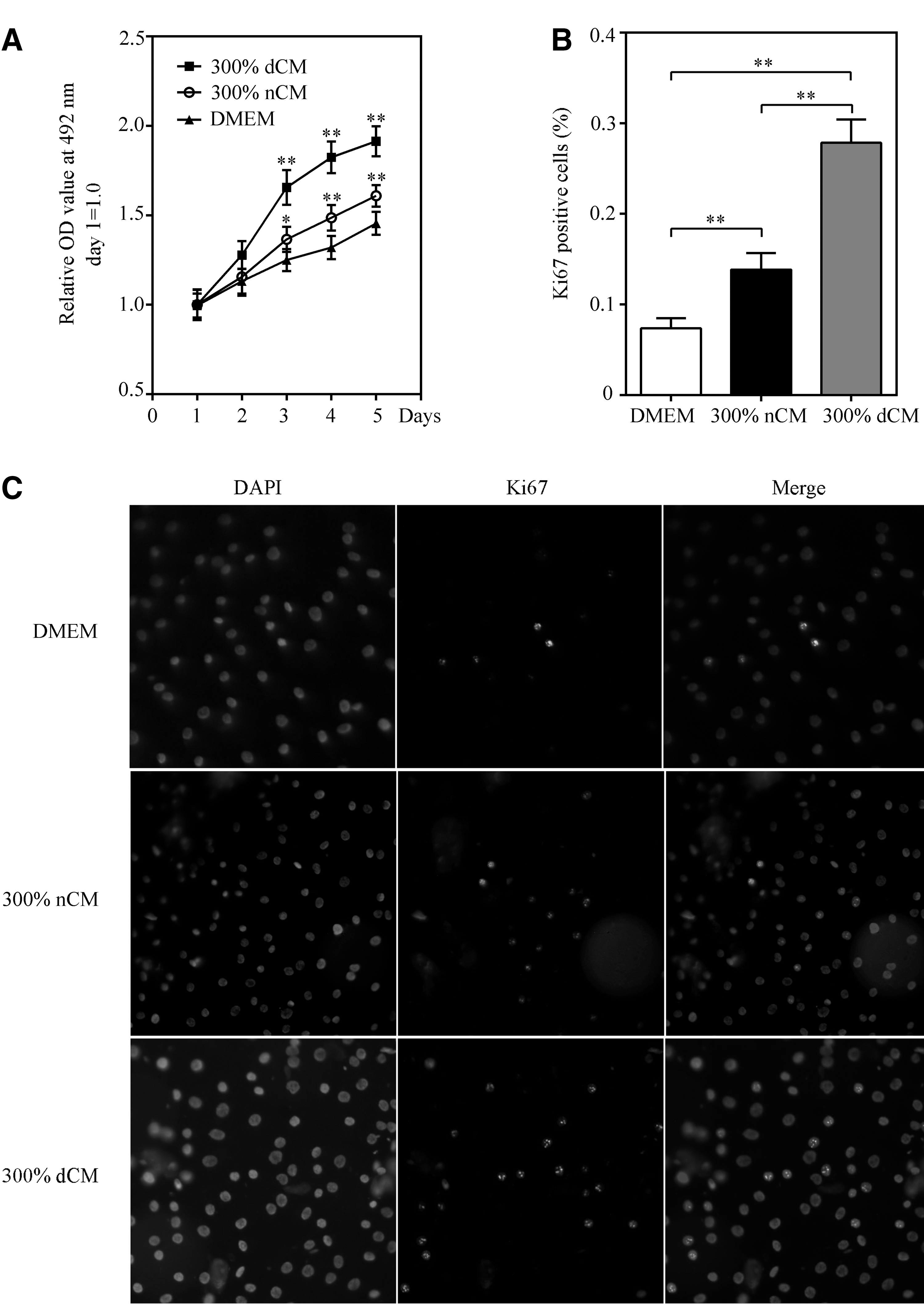
The effect of dCM on NPC proliferation. The proliferation of healthy NPCs was significantly stimulated by dCM in
Impact of dCM on ADAMTS, MMP, TIMP, and ECM gene expression in healthy NPCs
Healthy NPCs exhibited significantly increased expression of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 in the dCM group (Fig. 6). By contrast, TIMP-2, ACAN, and COL2 gene expression in healthy NPCs was significantly decreased in the dCM group.
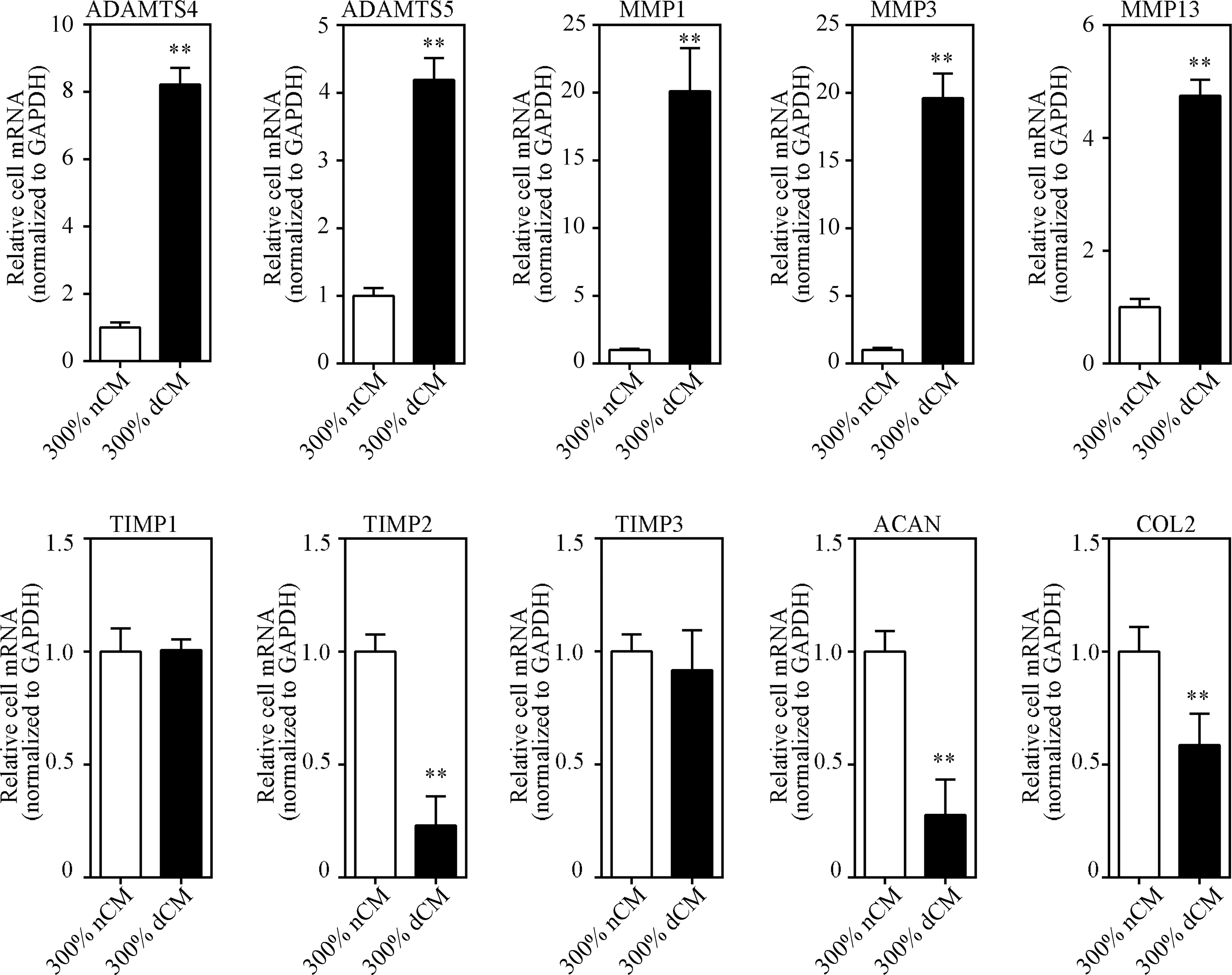
Impact of dCM on ADAMTS, MMP, TIMP, and ECM gene expression in healthy NPCs. Gene expression was measured using real-time RT-PCR, and mRNA levels of target genes in the nCM group (control) were normalized to 1. Healthy NPCs exhibited significantly increased expression of the catabolic factors, ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13, in the dCM group. By contrast, TIMP-2, ACAN, and COL2 gene expression in healthy NPCs was significantly decreased in the dCM group. dCM treatment modestly increased TIMP-1 expression and had no effect on TIMP-3 mRNA levels; n = 6 donors, donors 1–6 in nCM group (control), donors 1 and 7–11 in dCM group. **Indicates p < 0.01. ECM, extracellular matrix; MMP, matrix metallopeptidase.
Relative gene expression levels were compared between the dCM and nCM groups. MMP-1 and MMP-3 were expressed at higher levels in the dCM group compared with all other catabolic factors tested. Relative MMP-1 expression increased 20.1-fold, and MMP-3 expression increased 19.6-fold in 300% dCM. ADAMTS-4, ADAMTS-5, and MMP-13 showed moderately increased expression levels in dCM. ADAMTS-4 expression increased 8.2-fold, ADAMTS-5 expression increased 4.2-fold, and MMP-13 expression increased 4.7-fold in 300% dCM, respectively. In addition, TIMP-2 and ACAN expression decreased significantly by 0.2- and 0.3-fold in 300% dCM, respectively. COL2 expression decreased 0.6-fold in all three dCM concentrations. Finally, dCM treatment had no effect on TIMP-1 and TIMP-3 mRNA levels.
MAPK and NF-κB pathways are implicated in the dCM-mediated response of healthy NPCs
To determine the molecular pathways implicated in the dCM-mediated response of healthy NPCs, we investigated the phosphorylation of major signaling pathways associated with NPC proliferation and ECM metabolism (Fig. 7). We found that MAPK (ERK1/2, p38, and JNK) and NF-κB (p65) were phosphorylated after 0.5 h of dCM treatment, while phosphorylation returned to basal levels after exposure for 6 h. dCM treatment had no effect on total cellular levels of MAPK (ERK1/2, p38, and JNK) and NF-κB (p65).
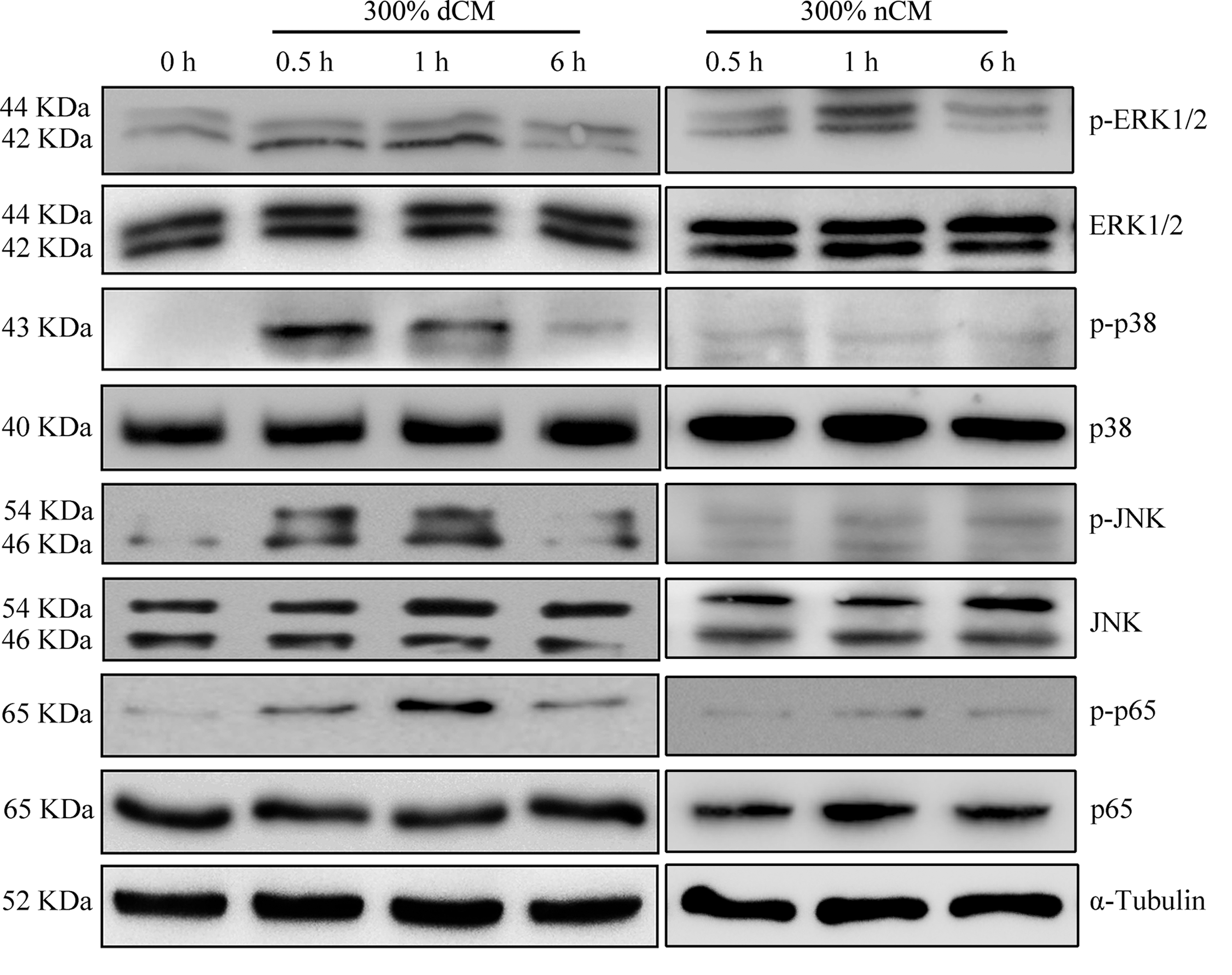
Western blot analysis of MAPK and NF-κB signaling proteins after treatment of NPCs with 300% dCM. Western blot analysis showed that MAPK (ERK1/2, p38, and JNK) and NF-κB (p65) were phosphorylated after 0.5 h of dCM treatment, while phosphorylation returned to basal levels after exposure for 6 h. dCM treatment had no effect on total cellular levels of MAPK (ERK1/2, p38, and JNK) or NF-κB (p65). MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor-kappa B.
Inhibition of MAPK and NF-κB pathways can weaken the effect of dCM
Inhibition of the ERK1/2 pathway significantly suppressed the dCM-stimulated NPC proliferation (Fig. 8). No significant changes of cell proliferation were observed when blocking the p38, JNK, and NF-κB pathways.

Involvement of MAPK and NF-κB pathways in the regulation of cell proliferation. Inhibition of the ERK1/2 pathway significantly suppressed dCM-stimulated NPC proliferation. No significant changes of cell proliferation were observed when blocking the p38, JNK, and NF-κB pathways. **Indicates p < 0.01.
To investigate the involvement of MAPK and NF-κB pathways in the regulation of ADAMTSs, MMPs, TIMPs, and ECM mRNA levels following dCM exposure, NPCs were treated with pharmacological inhibitors selective for the ERK1/2, p38, JNK, and NF-κB pathways (Fig. 9). The effect of inhibitors on control (without dCM treatment) cells is presented in Figure 9. The inhibition of the ERK1/2 pathway resulted in decreased gene expression of MMP-13, TIMP-1, TIMP-3, ACAN, and COL2 in control cells. No significant changes of ADAMTS, MMP, TIMP, and ECM gene expression in control cells were observed when blocking the p38, JNK, and NF-κB pathways.
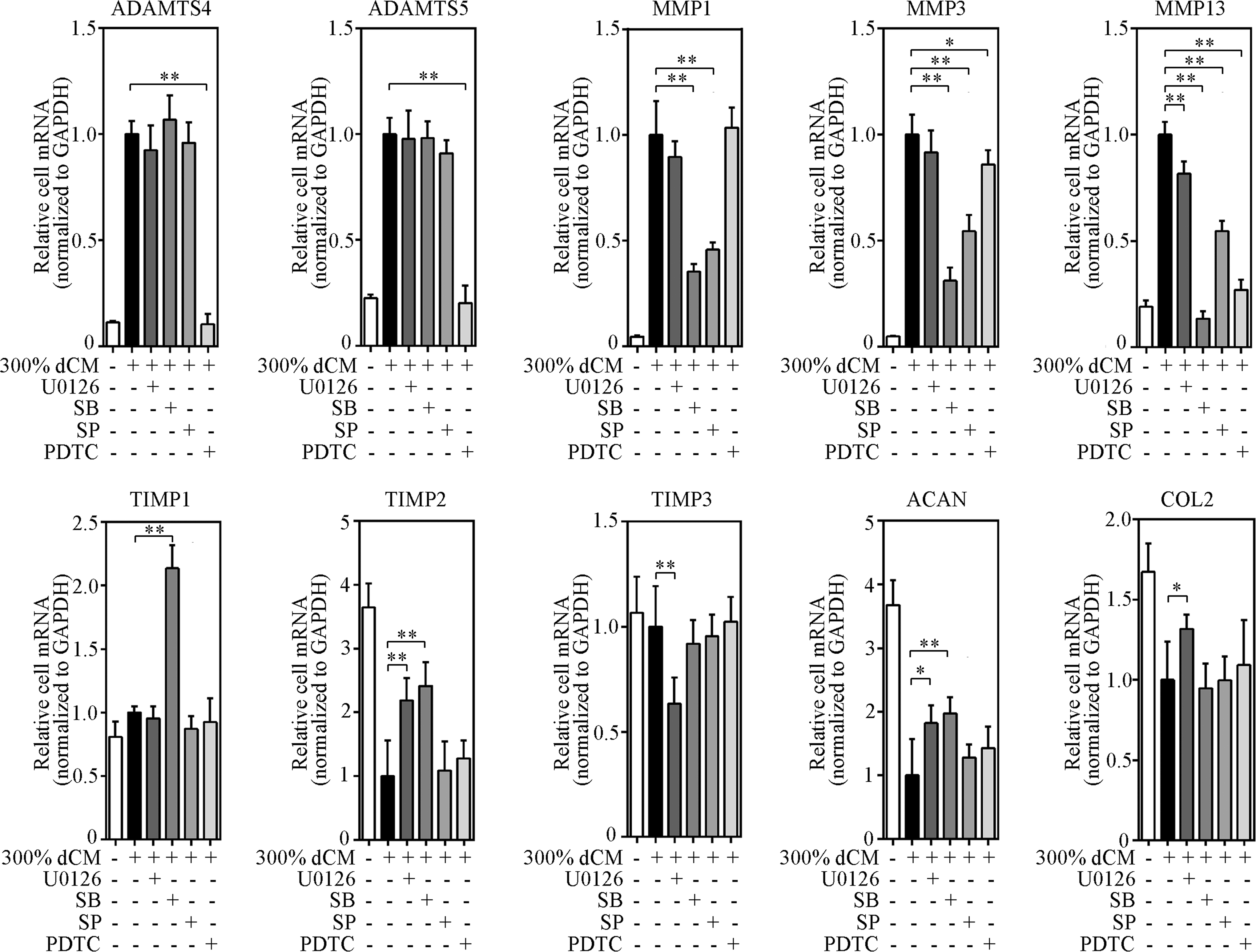
Involvement of MAPK and NF-κB pathways in the regulation of ADAMTS, MMP, TIMP, and ECM protein mRNA levels following exposure to 300% dCM. Specific pathway inhibitors were used and gene expression was measured using real-time RT-PCR; n = 6 donors, donors 1 and 7–11 in dCM group. *Indicates p < 0.05, **indicates p < 0.01. Inhibition of the NF-κB pathway significantly suppressed the dCM-stimulated upregulation of ADAMTS-4 and ADAMTS-5 mRNA by 90% and 80%, respectively, while the upregulation of ADAMTS-4 and ADAMTS-5 mRNA appeared to be independent of all three MAPK pathways. Inhibition of the p38 or JNK pathway significantly suppressed the dCM-stimulated upregulation of MMP-1 mRNA by 65% and 54%, respectively. These effects were similar to the dCM-stimulated upregulation of MMP-3 mRNA, which was suppressed by 69% after selective p38 inhibition and 46% after selective JNK inhibition. MMP-13 mRNA upregulation by dCM was attenuated by inhibition of the ERK1/2 (18%), p38 (86%), JNK (45%), and NF-κB (73%) pathways, suggesting that all four pathways regulated MMP-13 expression. dCM modestly increased TIMP-1 expression and when p38 was inhibited with SB, a twofold increase in TIMP-1 mRNA was observed. Although independent of the JNK and NF-κB pathways, dCM-induced TIMP-2 mRNA downregulation was prevented through selective inhibition of the ERK1/2 (2.2-fold) and p38 (2.4-fold) pathways. These effects were similar to dCM-induced ACAN mRNA downregulation, which increased by 1.8- and 2-fold by selective ERK1/2 and p38 inhibition, respectively. TIMP-3 mRNA expression was significantly reduced after ERK1/2 inhibition (>37% suppression with 10 μmol/L U0126) and appeared to be independent of the other three pathways. ERK1/2 pathway inhibition prevented dCM-induced downregulation of COL2 mRNA levels (1.3-fold). Treatment with the other three pathway inhibitors did not significantly alter the dCM-induced downregulation of COL2 mRNA levels.
Inhibition of the NF-κB pathway significantly suppressed the dCM-stimulated upregulation of ADAMTS-4 and ADAMTS-5 mRNA by 90% and 80%, respectively, while this upregulation appeared to be independent of all three MAPK pathways. Inhibition of the p38 and JNK pathways significantly suppressed dCM-stimulated MMP-1 mRNA upregulation by 65% and 54%, respectively. These effects were similar to dCM-stimulated MMP-3 mRNA upregulation, which was suppressed by 69% after p38 inhibition and 46% after JNK inhibition. dCM-stimulated MMP-13 mRNA upregulation was suppressed by inhibition of ERK1/2 (18%), p38 (86%), JNK (45%), and NF-κB (73%) pathways, suggesting that all four pathways regulated MMP-13 expression.
dCM modestly increased TIMP-1 expression, and inhibition of p38 with SB yielded a twofold increase in TIMP-1 mRNA (Fig. 9). Although independent of the JNK and NF-κB pathways, dCM-induced TIMP-2 mRNA downregulation was prevented through inhibition of ERK1/2 (2.2-fold in the 300% dCM group) or p38 (2.4-fold in the 300% dCM group) pathways. These effects were similar to the dCM-induced downregulation of ACAN mRNA, which increased by 1.8- and 2-fold after ERK1/2 and p38 inhibition, respectively. TIMP-3 mRNA expression was significantly reduced by ERK1/2 inhibition (>37% suppression with 10 μmol/L U0126), but appeared to be independent of the other three pathways. ERK1/2 pathway inhibition prevented dCM-induced downregulation of COL2 mRNA by 1.3-fold. None of the other pathway inhibitors significantly altered dCM-induced downregulation of COL2 mRNA levels.
The effect of MAPK and NF-κB pathway inhibitors on control (without dCM treatment) cells
The inhibition of the ERK1/2 pathway resulted in decreased gene expression of MMP-13, TIMP-1, TIMP-3, ACAN, and COL2 in control cells. No significant changes of ADAMTS, MMP, TIMP, and ECM gene expression in control cells were observed when blocking the p38, JNK, and NF-κB pathways (Fig. 10).

The effect of MAPK and NF-κB pathway inhibitors on control (without dCM treatment) cells. The inhibition of the ERK1/2 pathway resulted in decreased gene expression of MMP-13, TIMP-1, TIMP-3, ACAN, and COL2 in control cells. No significant changes of ADAMTS, MMP, TIMP, and ECM gene expression in control cells were observed when blocking the p38, JNK, and NF-κB pathways; n = 6 donors, donors 1–6. *Indicates p < 0.05, **indicates p < 0.01.
Treatment of NPCs with TGF-β1 altered the effect of dCM
The MTT assay was used to determine the effect of TGF-β1 on NPC proliferation. Human NPC proliferation was significantly stimulated in the TGF-β1 and TGF-β1 + 300% dCM groups (Fig. 11). Treatment of healthy NPCs with TGF-β1 following exposure to dCM elicited an additional proliferative response.

The effect of 5 ng/mL TGF-β1 on NPC proliferation. The MTT assay was used to determine the effect of TGF-β1 on NPC proliferation. Human NPC proliferation was significantly stimulated in the TGF-β1 and TGF-β1 + 300% dCM groups; n = 6 donors, donors 1 and 7–11. *Indicates p < 0.05. MTT, 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide; TGF-β1, transforming growth factor-β1.
Real time polymerase chain reaction (RT-PCR) showed that dCM-induced upregulation of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 mRNA was prevented following treatment with 5 ng/mL TGF-β1. By contrast, TIMP-1, TIMP-2, TIMP-3, ACAN, and COL2 gene expression significantly increased in the TGF-β1-treated group (Fig. 12).
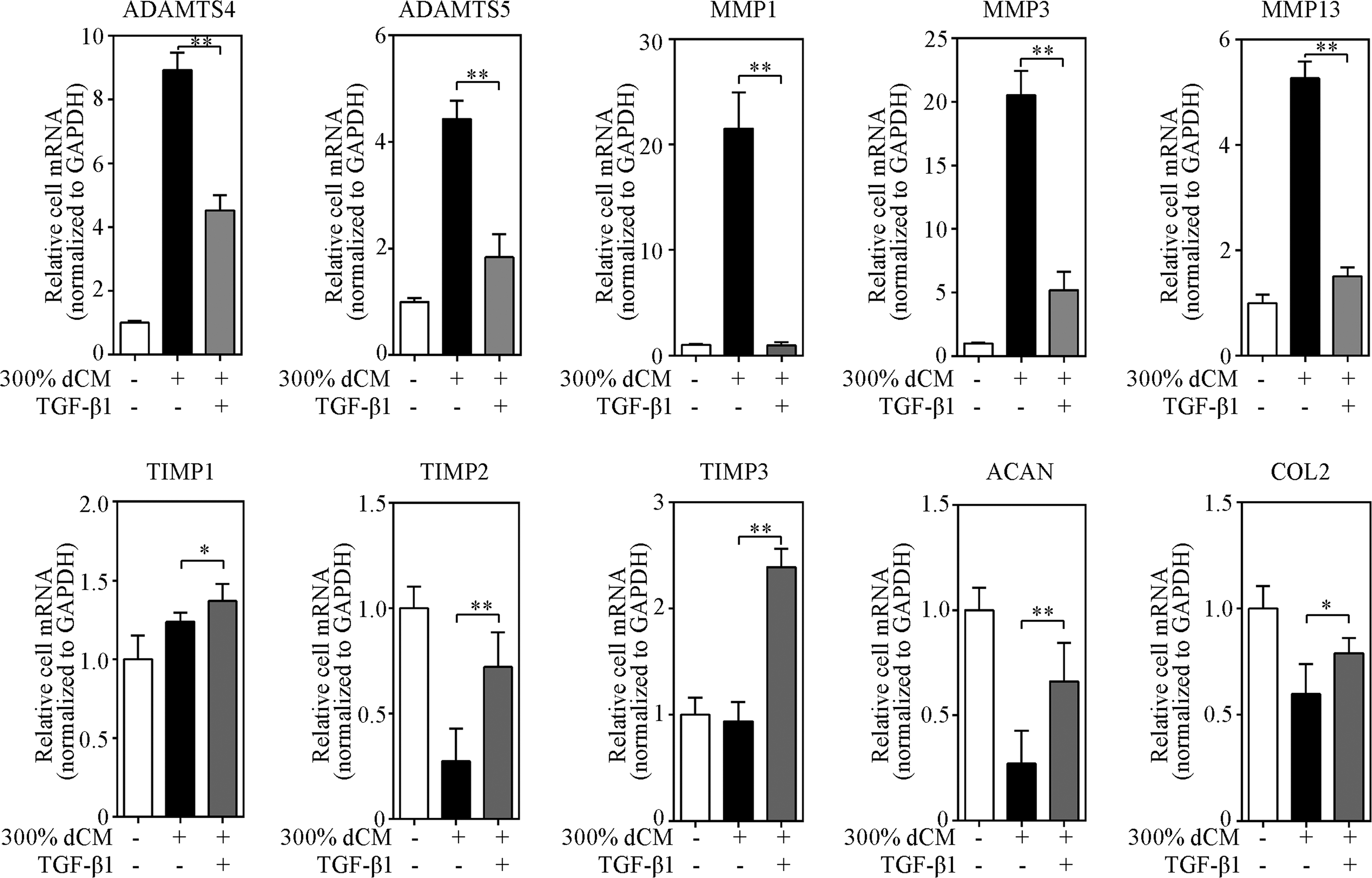
Treatment of NPCs with 5 ng/mL TGF-β1 altered the effect of dCM. RT-PCR showed that dCM-induced upregulation of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 mRNA was prevented following treatment with 5 ng/mL TGF-β1. By contrast, the gene expression of TIMP-1, TIMP-2, TIMP-3, ACAN, and COL2 appeared to significantly increase in the TGF-β1-treated group; n = 6 donors, donors 1 and 7–11. *Indicates p < 0.05, **indicates p < 0.01.
Discussion
Recent studies have demonstrated new therapeutic strategy using transplantation of seed cells to preserve IVD structure and functions. It is important to understand whether and how the exogenous cells survive and thrive in the hostile microenvironment of the degenerated IVD.
During the course of IVD degeneration, changes, including increased cell senescence, less production of ACAN and COL2, increased proteinases and cytokines, and acidic pH, have been observed (Huang et al., 2013). These events will present an unfavorable microenvironment for the endogenous cells and cells to be introduced into the disc in cell-based therapy. Inflammatory cytokines play a major role in the pathogenesis of disc degeneration and the hostile microenvironment (Risbud and Shapiro, 2014). Mediated through cytokines (such as TNF-α, IL-6, and IL-8), degenerated NP is a biologically active tissue capable of responding to proinflammatory factors and produces cytokines (Huang et al., 2013). Apart from the effects on promoting matrix breakdown, cytokines in IVD also regulated the behavior of transplanted seed cells (Krock et al., 2015). Thus, caution must be taken when opting to use cell-based therapies. So, we investigated the survival and function of transplanted healthy NPCs in mimical degenerated NP microenvironment ex vivo.
On the other hand, the microenvironment of the degenerate disc could be modified with other therapeutics, making it more accepting to the implanted stem cells. For example, stem cells may be transplanted in the presence of either anti-inflammatory agents or inhibitors of toll-like receptor signaling to neutralize negative effects on cell differentiation, tissue matrix repair, and increased inflammatory factor production (Krock et al., 2015). In addition, there is substantial evidence that mesenchymal stem cells (MSCs) are able to suppress inflammatory reactions in the degenerated NP tissue (Benneker et al., 2014). However, there are serious concerns associated with MSC transplantation, including poor cell survival, cell leakage through injection site (Vadala et al., 2012), unintended differentiation toward osteogenesis, and the potential of tumorigenesis (Benneker et al., 2014). Autologous or allogeneic chondrocytes or NPCs appear safe and effective in initial clinical trials (Benneker et al., 2014; Sakai and Andersson, 2015). Thus, in this study, we examined whether treatment of transplanted healthy NPCs with TGF-β1 could protect them from an inflammatory environment due to the anti-inflammatory property of TGF-β1 (Seifarth et al., 2009).
Combined with the previous and present studies, it is easy to conclude that cell therapy for repair of degenerate discs holds great therapeutic promise; however, several aspects of their use must be considered before placement into the degenerated microenvironment. The present study found that the repair ability of implanted cells could be negatively influenced by inflammatory cytokines, which suggested that severely degenerated discs with remarkable inflammation are likely unsuitable candidates for cell therapy. The present results and opinions lead researchers to focus on the effects of disc microenvironment on seed cell survival and function before transferring cell transplantation from bench to bedside successfully.
Multiple studies have suggested that inflammatory factors are present in degenerating disc tissue, and isolated IVD cells can actively secrete these factors in culture (Le Maitre et al., 2005, 2007; Seguin et al., 2005; Shamji et al., 2010; Cuellar et al., 2013). A previous study found that the gene expression of numerous cytokines and chemokines was increased in degenerated samples compared with nondegenerated samples (Phillips et al., 2013). The above study also indicates that NPCs are a source of cytokines and chemokines within the IVD and that these expression patterns are altered in IVD pathology (Phillips et al., 2013). In addition, other studies found that the inflammatory cytokine concentration increased in the blood of patients with LBP and may be a predictor of response to treatment of LBP with epidural steroid injection (Weber et al., 2015; Wang et al., 2016).
Thus, increased level of inflammatory factors in disc tissue is associated with disc degeneration, but whether degenerating discs release sufficient concentrations of factors that induce the physiological dysfunction of transplanted healthy NPCs when conducting a cell transplantation treatment for IVDD remains unclear. In this study, degenerated NP tissue (Pfirrmann grade V) and normal NP tissue (Pfirrmann grade I) from human were cultured ex vivo, and inflammatory factors released by NP tissue into culture media were quantified. The expression of TNF-α, IL-1β, IL-1α, IL-2, IL-4, IL-6, IL-8, IL-10, IL-17, IFN-γ, and PGE2 increased, while TGF-β1 expression decreased in dCM compared with the nCM. The results suggest that degenerated NP exhibits an inflammatory environment. The present study also indicates that degenerated NP tissue releases sufficient concentrations of cytokines that induce the physiological dysfunction of transplanted healthy NPCs when conducting a cell transplantation treatment for IVDD.
Usually, experimental conditions mimicking degenerative discs and inflammatory environments in IVDD are created by adding one or two cytokines to cultured NPCs. However, a cocktail of cytokines secreted by NPCs together promote the progress of pathological changes in degenerated discs (Risbud and Shapiro, 2014). Although previous research has revealed the impact of these factors individually on disc cells, their combined effect has rarely been studied. The combined effect on cell proliferation and function was not consistent with that of any one specific factor. This could be due to the presence of other regulatory factors or even specific inhibitors. Another explanation could be that various factors in dCM form complexes with binding proteins and are released into the pericellular environment.
Therefore, the present study uses an NP tissue culture approach to compare the profile of factors released from degenerated discs with healthy discs to establish a more direct link to in vivo disc degeneration. We investigated the survival and function of transplanted healthy NPCs grown in dCM to represent the paracrine effects of degenerated NPCs. In addition, proteins in dCM were concentrated using centrifugal ultrafilters to enhance their biological effects. In a similar study, degenerating IVDs from low back pain patients and healthy painless IVDs from human organ donors were cultured ex vivo, and inflammatory and nociceptive factors released by IVDs into culture media were quantified (Krock et al., 2014). However, the purpose of the above study was to investigate whether painful IVDs release sufficient concentrations of nociceptive factors that induce neuronal plasticity and neoinnervation, which is different from ours. To our knowledge, the present study is the first to explore the paracrine effects of degenerated NPCs on healthy human NPCs through dCM exposure.
Normal IVDs are characterized by a very low cell proliferation rate, which intensifies during aging and degeneration, especially in cell clusters (Johnson et al., 2001). This proliferation probably represents attempted tissue repair and can be triggered by exogenous or endogenous cytokines. Some cytokines secreted by degenerated NPCs stimulate mitosis (Pratsinis et al., 2012), while others suppress cell proliferation (Zhang et al., 2014).
In this study, we found that dCM promoted healthy NPC proliferation in vitro by activating the ERK1/2 signaling pathway. These results were similar to those of a previous study that observed the effects of autocrine factors on human IVD cell proliferation (Pratsinis et al., 2012). Furthermore, we found that TGF-β1 modestly enhanced the effects of dCM on cell proliferation. The effects of TGF-β1 on chondrocyte and disc cell proliferation have been widely studied, but with conflicting results dependent on the culture conditions, culture type (e.g., monolayer, three-dimensional, suspension), and physiopathological origin of the sample. The results obtained in this study depended on the conditions mimicking the inflammatory environment of IVDD, Pfirrmann grades of the donors, and the cell passage level.
Increasing the number of functional NPCs can be instrumental to rebuilding the structure and function of IVDs. Although dCM promoted healthy NPC proliferation in vitro, the anabolic–catabolic balance of the ECM was altered under these conditions. This could explain the existence of cell clusters and increase in cell density with reduced ECM in degenerated disc tissue.
Degradation and loss of ECM proteins (e.g., ACAN and COL2) is a hallmark of IVDD. It is believed that ACAN and COL2 destruction in IVDD is associated with the upregulation of proteinases, such as ADAMTSs and MMPs, and downregulation of TIMPs, which might be regulated by inflammatory cytokines (e.g., IL-1 and TNF-α) (Purmessur et al., 2013b; Risbud and Shapiro, 2014). Seguin et al. found a dose-dependent increase in MMP and ADAMTS gene expression and decrease in TIMP gene expression in NPCs obtained from bovine distal caudal spines exposed to TNF-α (Seguin et al., 2005).
In this study, we found that dCM induced the upregulation of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 in healthy NPCs. Conversely, treatment with dCM resulted in a decreased expression of TIMP-2, an endogenous inhibitor of MMPs. This suggested that the cocktail of cytokines secreted by degenerated NPCs induced the expression of the degenerated phenotype of NPCs in a paracrine manner. The inflammatory environment of degenerated discs is not only severe for resident disc cells but also challenges the functionality of transplanted seed cells. Although previous studies have found that NPC transplantation partially improves ECM metabolism in discs, the results from this study suggest that the environment of degenerated discs leads to dysfunction and degeneration of transplanted healthy NPCs over the short term. This obstacle to cell transplantation therapy cannot be neglected when exploring strategies for disc regeneration.
In the past few years, extensive therapeutic studies in the field of rheumatoid arthritis and osteoarthritis have highlighted the need to identify the underlying signaling pathways, prompting many IVD researchers to explore the molecular mechanisms involved in IVD homeostasis. Specific signal transduction pathways mediate the increased expression and synthesis of matrix metalloproteinases and inflammatory factors, in particular the MAPK and NF-κB pathways. MAPK and NF-κB have been identified as the major regulators of inflammation and catabolism in several musculoskeletal disorders (e.g., osteoarthritis), and growing evidence supports the importance of these signaling pathways in disc disease (Wuertz et al., 2012). However, it is still unclear which physiological stressors trigger NF-κB or MAPK activation and how disc matrix homeostasis is affected as a result of such activation, although some specific cytokines (e.g., TNF-α and IL-1β) are known to control ADAMTS-4 transcription in an MAPK- and NF-κB-dependent manner (Tian et al., 2013; Wang et al., 2014a).
In this study, we found that dCM induced NF-κB or MAPK activation, which led to catabolic enzyme upregulation and ECM downregulation. Furthermore, dCM treatment led to rapid and transient phosphorylation of MAPK and NF-κB. Inhibition of MAPK and NF-κB pathways significantly suppressed the effect of dCM on healthy NPCs. Suppression of MAPK and NF-κB pathways has become a major research target for treating diseases, including disc degeneration (Wuertz et al., 2012). The results from this study suggest that MAPK and NF-κB pathway inhibitors might have the therapeutic potential to protect transplanted healthy NPCs from dysfunction in the inflammatory environment of discs. This could provide a new target for improving the therapeutic effect of cell transplantation strategies.
Despite the promising potential of MAPK and NF-κB inhibitors, their risk-to-benefit ratio remains controversial due to their importance in numerous physiological processes. Inhibition of these signal pathways, especially inhibition of MAPK, could have detrimental side effects. For example, the ERK1/2-mediated pathway regulates growth factor signaling and the JNK-mediated pathway modulates cell proliferation, differentiation, and apoptosis. Inhibition of both pathways could lead to general cytotoxicity. In addition, the p38 MAPK pathway is involved in chondrocyte differentiation. Therefore, future research should examine MAPK and NF-κB inhibitors in blocking inflammation-mediated NPC degeneration.
In this study, we found that dCM containing a cocktail of cytokines activated NF-κB and MAPK and led to catabolic enzyme upregulation and ECM downregulation. This underlines the potential of anticytokine agents to ameliorate structural damage and ECM degeneration in IVDs, which could reduce unanticipated deleterious side effects compared with MAPK and NF-κB pathway inhibition.
Many researchers examined the inhibitory effect of TGF-β1 on inflammation and demonstrated that TGF-β1 may suppress inflammation in articular cartilage or other tissue (Rubtsov and Rudensky, 2007; Seifarth et al., 2009; Pinar Karapinar et al., 2016; Serban et al., 2016; Zhang et al., 2016). We found that TGF-β1 not only markedly suppressed ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 but also stimulated TIMP-2, TIMP-3, ACAN, and COL2, which could reverse the effects of dCM. This suggests that TGF-β1 could help promote the function of transplanted healthy NPCs in degenerated disc environments. However, the mechanism behind this scenario is yet to be defined. TGF-β1, as an immune suppressor, has been shown to inhibit NF-κB-dependent inflammatory responses in multiple cell types (Azuma et al., 1999; Cavin et al., 2003; Rautava et al., 2012). However, whether TGF-β1 could inhibit NF-κB-dependent inflammatory responses in NPCs is not clear up to now, and further research should be done in this field.
Research indicates that growth factors can antagonize the upregulation of ADAMTSs and MMPs by inflammatory cytokines in disc cells (Cho et al., 2013; Wang et al., 2014b; Yang et al., 2015b). For example, Wang et al. (2014b) demonstrated that the increase of ADAMTS-4 and ADAMTS-5 in NPCs in response to TNF-α was significantly decreased following bone morphogenetic protein-7 (BMP-7) treatment, which led to decreased matrix macromolecule degradation. Similarly, Cho et al. (2013) showed that BMP-2 and TGF-β1 antagonized the TNF-α and IL-1β-induced upregulation of MMP-1 in AF cells. The results of this study suggest that TGF-β1 could abolish the catabolic effects of inflammatory environments on transplanted healthy NPCs. These findings could provide a deeper understanding of the pathophysiology of disc degeneration and substantially facilitate the development of cell transplant therapy combined with TGF-β1 as a treatment strategy.
There were some limitations in this study. For example, we selected donors with an average age of 39.67 in the nondegenerated group and an average age of 59.5 in the degenerated group. Unlike animal experiments, it is hard to select donors with the same age in the two groups. We have tried to minimize the difference between ages when selecting donors, but it is hard to obtain nondegenerated IVDs (Pfirrmann grade I) from donors at age 60. In addition, we think that it may not be necessary to match the age in the two groups. To date, promoters of IVD degeneration have extended from aging to excessive mechanical loading (Xing et al., 2010; Purmessur et al., 2013a), disc injury (Park et al., 2014; Cai et al., 2015), oxidative stress (Nasto et al., 2013; Hou et al., 2014), diabetes (Won et al., 2009; Illien-Junger et al., 2013), chronic tobacco smoking (Wang et al., 2012; Nasto et al., 2014), obesity (Samartzis et al., 2012; Takatalo et al., 2013), and genetic inheritance. Thus, age is one of the factors that contribute to degenerated condition. The degenerated disc microenvironment was discussed in this research, including age-related degeneration, stress-related degeneration, and so on.
In summary, we demonstrated that activation of both the MAPK and NF-κB signaling pathways is involved in dCM-mediated upregulation of ADAMTS-4, ADAMTS-5, MMP-1, MMP-3, and MMP-13 and downregulation of TIMP-2, ACAN, and COL2 in human healthy NPCs. Furthermore, TGF-β1 could antagonize the dCM-induced dysfunction of transplanted seed cells. Therefore, the paracrine effects of degenerated disc cells on human healthy NPCs are mediated by the MAPK and NF-κB pathways and can be reduced by TGF-β1.
Footnotes
Acknowledgments
The work was supported by the National Natural Science Foundation of China (81272035; 81572190; 81572170; 81572109; 81201423), the 51st China Postdoctoral Science Foundation 2012 (2012M511800), the Fundamental Research Funds for the Central Universities (KYLX_0202), and the Jiangsu Provincial Special Program of Medical Science (BL20122004). The English in this document has been checked by at least two professional editors, both native speakers of English.
Disclosure Statement
No competing financial interests exist.