
Abstract
Gelatin has been reported to induce generation of mesenchymal stem cells (MSCs) with enhanced potential of differentiation into neuronal lineage cells. However, the presence of various cell types besides MSCs in bone marrow has raised doubts about the effects of gelatin. In the following report, we determined whether gelatin can directly enhance neurogenic differentiation potential in MSCs without proliferation of neural progenitor cells (NPCs). MSCs comprised a high proportion of bone marrow-derived primary cells (BMPCs) and gelatin induced significant increases in MSC proliferation during primary culture, and the proportion of MSCs was maintained at more than 99% throughout the subculture. However, NPCs comprised a low percentage of BMPCs and a decrease in proliferation was detected despite gelatin treatment during the primary culture, and the proportion of subcultured NPCs gradually decreased. In a similar manner, MSCs exposed to gelatin during primary culture showed more enhanced neurogenic differentiation ability than those not exposed to gelatin. Together, these results demonstrate that gelatin directly enhances neurogenic differentiation in bone marrow-derived MSCs without stimulating NPC proliferation.
Introduction
M
To date, differentiation of MSCs into neuronal lineage cells has been induced using chemical compounds and/or growth factors, biomaterials, coculture system, and purified extracellular matrix (ECM) proteins. Tretinoin (ATRA) (Komori et al., 2013; Chu et al., 2015), dimethyl sulfoxide/butylated hydroxyanisole (DMSO/BHA) (Khanabdali et al., 2015; Mu et al., 2015), indomethacin/3-isobutyl-1-methylxanthine (Indomethacine/IBMX) (Wang et al., 2013; Mu et al., 2015), and 2-mercaptoethanl (BME) (Khanabdali et al., 2015; Mu et al., 2015) as chemical compounds, epidermal growth factor (EGF) (Huat et al., 2014; Mukai et al., 2016), basic fibroblast growth factor (bFGF) (Huat et al., 2014; Mukai et al., 2016), and brain-derived neurotrophic factor (BDNF) (Wang et al., 2013; Tsai et al., 2014) as growth factors, cerebrospinal fluid (Ye et al., 2011) and exosome (Takeda and Xu, 2015) as biomaterials, neural stem cells (Rong et al., 2015) and retinal pigmented epithelial cells (Yang et al., 2010) as coculture systems, and collagen (Mruthyunjaya et al., 2011), laminin (Lee et al., 2011), and tenascin (Tsai et al., 2014) as ECM proteins have been known as stimulators generating neuronal lineage cells from MSCs.
Recently, in a previous study (Han et al., 2013), the differentiation potential of MSCs into neuronal lineage cells was enhanced by exposure to gelatin during primary culture. However, improvement of neurogenic differentiation potential may not have resulted from the proliferation of MSCs transformed directly by gelatin, but rather by neural progenitor cells (NPCs) attached preferentially to gelatin. This possibility is consistent with studies reporting that primary cells isolated from bone marrow include diverse types of cells, such as Fukata et al. (2013); Ghosn et al. (2012) MSCs (Ben Azouna et al., 2012; Takemitsu et al., 2012; Li and Ikehara, 2013), NPCs (Ricciardi et al., 2012; Neirinckx et al., 2013), hematopoietic stem cells (Ghosn et al., 2012; Fukata et al., 2013), and lymphoid progenitor cells (Pang et al., 2011; Anthony and Link, 2014). To definitively establish whether gelatin enhances the neurogenic differentiation potential to MSCs without any proliferation of NPCs among primary cells derived from bone marrow, we characterized the percentages of MSCs and NPCs in primary cells isolated from bone marrow. These cells were cultured with gelatin during primary culture and then subcultured without gelatin, followed by comparison of their differentiation into neuronal lineage cells.
Materials and Methods
Animals
Three-week-old male Sprague-Dawley (SD) rats were purchased from DBL (Eumseong, Korea) and were used as bone marrow cell donors. All protocols for animal experiments, including housing and handling of the animals, were approved by the Institutional Animal Care and Use Committee (IACUC) of Kangwon National University (IACUC approval No. KW-121101-1) and performed according to the Animal Care and Use Guidelines of Kangwon National University.
Retrieval of primary cells from bone marrow
Tibias and femurs, surgically obtained from the hind limbs of SD rats sacrificed by CO2 asphyxiation, were washed with Dulbecco's phosphate-buffered saline (DPBS; Welgene, Daegu, Korea) supplemented with 1% (v/v) antibiotic–antimycotic (Welgene). After removing muscle tissue from bones, the marrow cavities were exposed by cutting the spongious end of each bone. The bones were then flushed through the bone cavity with DPBS supplemented with 2% (v/v) heat-inactivated fetal bovine serum (FBS; Welgene) to collect primary cells. Elimination of red blood cells (RBCs) from primary cells retrieved from the bone marrow was performed using the RBC lysis buffer (Sigma-Aldrich, St. Louis, MO). The bone marrow-derived primary cells (BMPCs) were counted using a hemocytometer.
Primary culture of BMPCs
The primary cells collected from bone marrow were cultured for 2 days on plates coated with or without 1% (w/v) gelatin derived from bovine skin (Cat. no. G9391; Sigma-Aldrich) in low-glucose Dulbecco's modified Eagle's medium (LG-DMEM; Welgene) supplemented with 10% (v/v) heat-inactivated FBS and 1% (v/v) antibiotic–antimycotic at 37°C in humidified 95% air and 5% CO2. Subsequently, nonadherent cells were discarded and replaced with a fresh culture medium at 2-day intervals for 14 days. The confluent primary cells were dissociated using 0.25% trypsin-EDTA (Welgene) and then used for subculture and flow cytometric analyses.
Subculture of bone marrow-derived stem cells
Five times 105 bone marrow-derived stem cells (BMDSCs) from each group were cultured on gelatin-free culture plates in LG-DMEM supplemented with 10% (v/v) heat-inactivated FBS and 1% (v/v) antibiotic–antimycotic in 95% humidified air and 5% CO2. BMDSCs that were approximately 80% confluent were detached with 0.25% trypsin-EDTA, and the cells were replated continuously at the same cell density and cultured under the same conditions to the fifth passage.
Differentiation of subcultured BMDSCs into neuronal lineage cells
The subcultured BMDSCs were exposed to a preinduction medium, consisting of high-glucose DMEM (HG-DMEM; Welgene) supplemented with 0.1 mM 2-mercaptoethanol (Gibco Invitrogen, Grand Island, NY) and 2% (v/v) dimethylsulfoxide (Sigma-Aldrich). After 5 h, the cells were incubated in the neuronal induction medium, consisting of HG-DMEM supplemented with 10% (v/v) heat-inactivated FBS, 1% (v/v) antibiotic–antimycotic, 10 μg/L basic fibroblast growth factor (b-FGF; Peprotech, Rocky Hill, NJ), 10 μg/L human epidermal growth factor (hEGF; Peprotech), 1 mM dibutyryl cyclic AMP (Sigma-Aldrich), and 0.5 mM isobutylmethylxanthine (Sigma-Aldrich). After incubation for 7 days, the neuronal lineage differentiated cells were washed with DPBS, harvested using 0.25% trypsin-EDTA, and analyzed by flow cytometry.
Flow cytometry
Cells were fixed using 4% (v/v) paraformaldehyde (Junsei Chemical, Chuo-ku, Japan) and washed in ice-cold DPBS. In case of non- or cultured BMPCs and cultured BMDSCs, the fixed cells were stained for 45 min at 4°C with allophycocyanin (APC)-conjugated anti-rat CD90 (BioLegend, San Diego, CA), an MSC marker, antibody in DPBS, and fluorescein isothiocyanate (FITC)-conjugated anti-nestin (Santa Cruz Biotechnology, Santa Cruz, CA), a neural stem/progenitor cell marker, antibody in DPBS containing 0.01% (v/v) Triton-X-100 (Sigma-Aldrich), respectively. The fixed neurogenic differentiated BMDSCs were incubated for 45 min at 4°C with anti-neuron-specific protein (NeuN; Millipore, Bedford, MA), a neuron-specific marker, anti-neural/glial antigen 2 (NG2; Millipore), an oligodendrocyte precursor cell-specific marker, or anti-glial fibrillary acidic protein (GFAP; Abcam, Cambridge, MA), an astrocyte-specific marker, antibodies. All antibodies were diluted in DPBS containing 0.01% (v/v) Triton X-100. The primary antibodies were detected by incubating with Alexa Fluor® 488 chicken anti-rabbit IgG (Molecular Probes, Eugene, OR) for 45 min at 4°C. Details of the antibodies and their dilutions are shown in Table 1. After washing with DPBS, the stained cells were sorted using a FACSCalibur (Becton Dickinson, Franklin Lakes, NJ), and data analyses were performed using BD CellQuest Pro Software™ (Becton Dickinson).
Statistical analysis
Data analysis was performed with the Statistical Analysis System software (SAS Institute, Cary, NC). Comparisons among treatment groups were performed using the least squares difference or Duncan's method. Statistically significant differences among treatments were evaluated by analysis of variance (ANOVA) using the SAS software. A value of p < 0.05 was regarded as a statistically significant difference.
Results
The proliferative effects of gelatin during primary culture of MSCs and NPCs from BMPCs
To identify the existence of NPCs in cell populations derived from bone marrow, we first characterized the proportion of MSCs and NPCs in BMPCs. Figure 1 shows that 64.15% ± 13.13% of the total cells were MSCs, and NPCs comprised a small percentage (2.44% ± 2.12%) of the total cells. The percentages of MSCs and NPCs in BMPCs before or after culturing on gelatin were then determined. Compared with those derived directly from bone marrow without culturing on gelatin, BMPCs cultured on gelatin showed a significant increase in the percentage of CD90-positive (a MSC marker) cells (Fig. 2A, C) and did not show any significant increase of nestin (an NPC marker) (Fig. 2B, D). In addition, there was a preferential decrease in nestin-positive cells in BMPCs cultured on gelatin (Fig. 2B, D). In contrast to MSCs, the results show that NPCs existing in a heterogeneous cell population derived from bone marrow did not proliferate in the presence of gelatin.
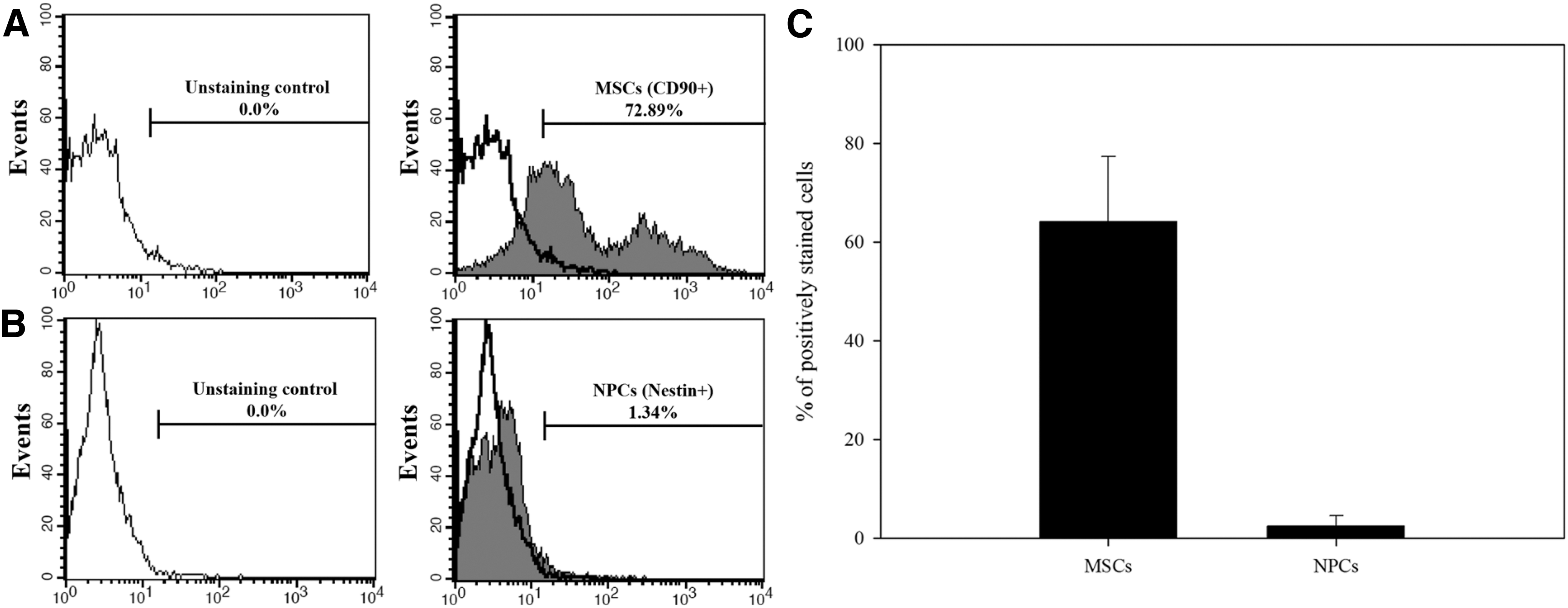
The proportion of MSCs and NPCs in the primary cell population retrieved from bone marrow. Primary cells derived from bone marrow were stained with APC-conjugated anti-CD90 (an MSC marker) and FITC-conjugated anti-nestin (an NPC marker) antibodies. Subsequently, the percentage of cells expressing CD90 and nestin was determined using flow cytometry
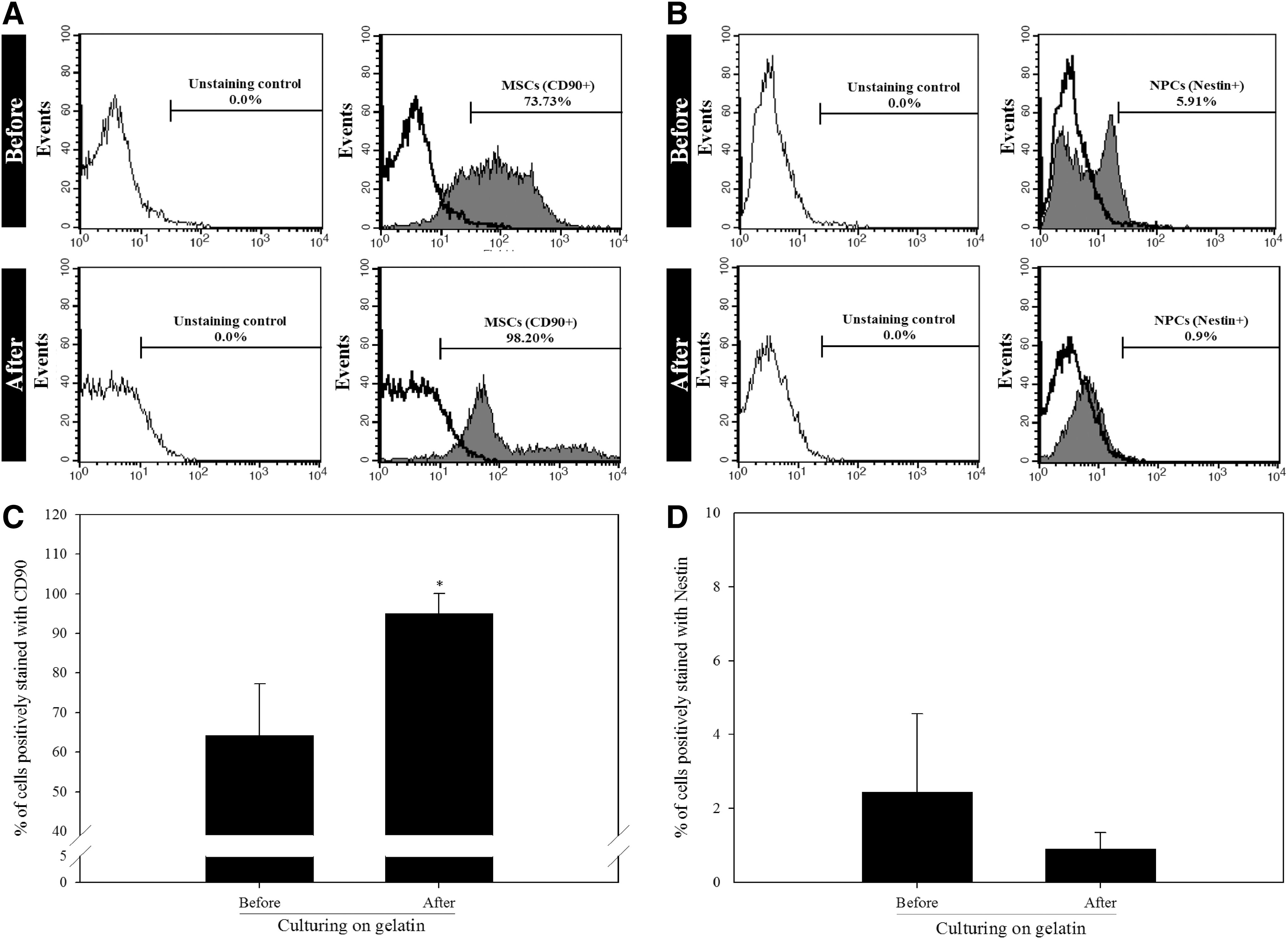
The effects of gelatin during primary culture on the percentage of MSCs and NPCs among BMPCs. Primary cells collected from bone marrow were cultured on plates coated with 1% (w/v) gelatin. After 14 days of culture, flow cytometry was conducted after staining the primary non- or cultured BMPCs with fluorescently conjugated primary antibodies against CD90 (an MSC marker)
The proliferative competence of subcultured MSC and NPC postprimary cultures in the presence of gelatin
To identify whether NPCs could proliferate during subculturing, the percentages of NPCs and MSCs in each passage were determined in BMDSCs subcultured in a postprimary MSC culture system in the presence of gelatin. Figure 3 shows that approximately 99% of the cells were identified as MSCs expressing CD90 in all passages, whereas the percentage of NPCs expressing nestin gradually decreased with increasing passage number, most notably in the fifth passage where only 0.73% ± 0.67% NPCs were detected. These results show that NPCs do not proliferate during subculture in a general MSC culture system.
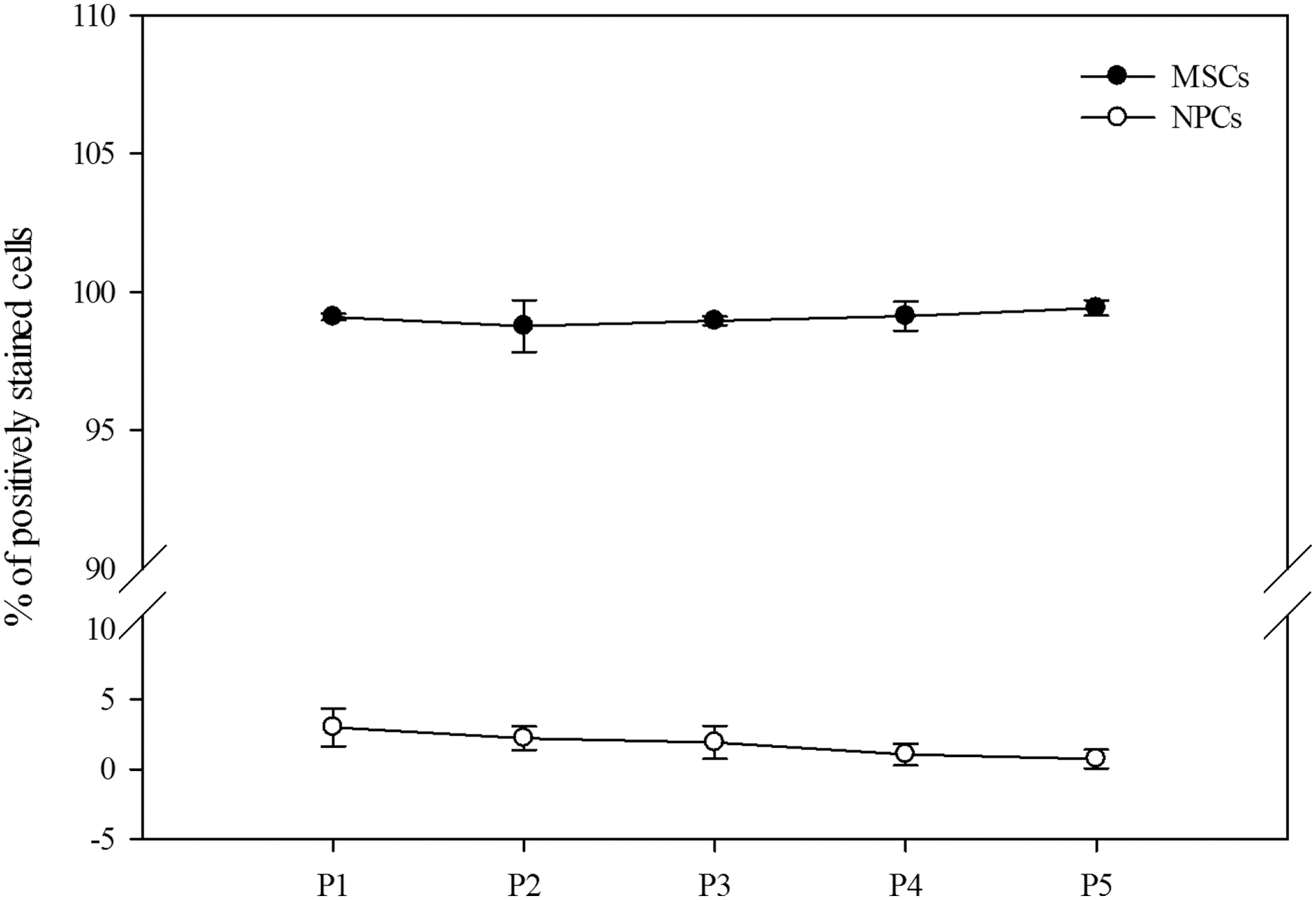
The percentage of MSCs and NPCs among BMDSCs subcultured postprimary culture in the presence of gelatin. The culturing of BMPCs on plates coated with 1% (w/v) gelatin was conducted for 14 days. Subsequently, the cultured BMDSCs continued to be cultured on gelatin-free culture plates until passage five. Flow cytometry was conducted after staining the confluent BMDSCs at each passage with fluorescently conjugated primary antibodies against CD90 (an MSC marker) and nestin (an NPC marker). Extremely low yields (from 3.00% ± 1.34% to 0.73% ± 0.67%) of NPCs (open circle), but high yields (from 99.10% ± 0.12% to 99.42% ± 0.28%) of MSCs (black circle) were detected at each passage, and no significant differences in the percentages of MSCs and NPCs were found with increasing passage number. The data are expressed as mean ± SD of three independent experiments. BMDSC, bone marrow-derived stem cell.
The identification of enhanced neurogenic differentiation potential in postprimary MSCs cultured in the presence of gelatin
Whether gelatin can induce enhanced neurogenic differentiation of MSCs was confirmed by evaluating the differentiation ratios of neuronal lineage cells of MSCs subcultured in postprimary culture in the presence of gelatin at passage five. Although there was no significant difference among groups, there was a higher ratio of differentiation into neurons expressing NeuN, oligodendrocytes expressing NG2, and astrocytes expressing GFAP when compared with MSCs not exposed to gelatin during the primary culture (Fig. 4). These results show that the neurogenic differentiation potential of MSCs can be enhanced by gelatin during primary culture. At the same time, we confirmed that the enhancement of MSC neurogenic differentiation potential by gelatin during primary culture did not result from NPC contamination.
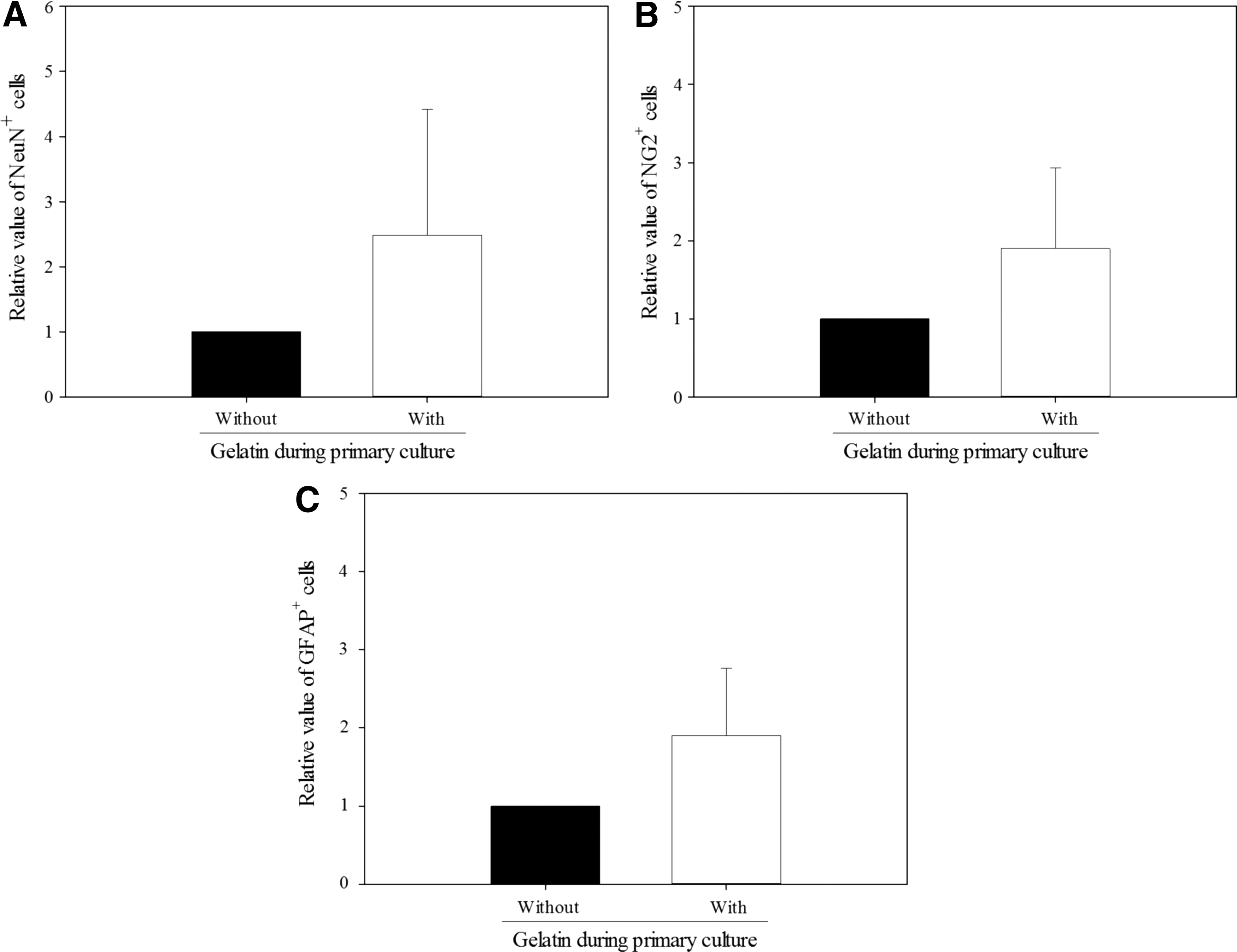
Neurogenic differentiation potential of MSCs derived from BMPCs exposed to gelatin during primary culture. BMPCs were cultured on plates coated without (“Without”) or with (“With”) 1% (w/v) gelatin for 14 days. MSCs were then derived by culturing BMDSCs from each group in gelatin-free culture conditions until passage five. MSCs differentiated into neuronal lineage cells after incubating cells for 7 days in differentiation medium. They were then stained with fluorescence-conjugated primary antibodies against neuron-specific protein (NeuN), a neuron-specific marker, neural/glial antigen 2 (NG2), an oligodendrocyte precursor cell-specific marker, and GFAP, an astrocyte-specific marker, followed by quantitation of fluorescence using flow cytometry. MSCs derived from BMPCs exposed to gelatin during primary culture showed higher percentages of NeuN
Discussion
NPCs present in cells derived from bone marrow do not proliferate in an MSC culture system. In primary cultures, the MSC culture system combined with gelatin did not induce proliferation of the small number of NPCs in the BMPC population. Moreover, the percentage of NPCs subcultured in the gelatin-free MSC culture system gradually diminished in postprimary culture and was rarely detected in the fifth passage. Simultaneously, the MSCs from BMPCs cultured in the gelatin-combined MSC culture system during primary culture showed more enhanced neurogenic differentiation ability than those in the gelatin-free MSC culture system. These results demonstrate that gelatin directly enhances neurogenic differentiation of bone marrow-derived MSCs without stimulation of NPC proliferation.
As shown in Figures 2 and 3, NPCs did not proliferate in the MSC culture system, which may have resulted from an inappropriate culture system for NPC proliferation. When culturing NPCs, EGF and bFGF play important roles in stimulating NPC proliferation (Fiorelli et al., 2013; Mothe and Tator, 2013) and neurosphere formation (Mothe and Tator, 2013; Walker and Kempermann, 2014). The absence of these mitotic growth factors in the MSC culture system significantly contributes to inducing inhibition of NPC proliferation, suggesting that screening of specific cells in the mixed cell population can be effectively conducted by removing growth factors that activate the target cells.
Gelatin, a soluble protein produced by hydrolysis of collagen, has been well known to promote cell adhesion, proliferation, and stem cell differentiation by the arginine–glycine–aspartic acid (RGD) sequence that existed in gelatin (Gautam et al., 2013; Nair et al., 2015). However, the mechanism on enhancement of MSC neurogenic differentiation by gelatin is still not clearly understood. As a possibility, we can consider softness of gelatin with the mechanics of below 1 kPa (Even-Ram et al., 2006; Du et al., 2011; Nava et al., 2012), supported by the previous reports that soft ECM substrates with the mechanics of below 1 kPa can stimulate neurogenic differentiation of MSCs by blocking the BMP/Smad signaling pathway (Engler et al., 2006; Du et al., 2011), and the β1 integrin internalization enhanced by soft matrix led to the expression of neuronal genes such as microtubule-associated protein 2 (MAP2), neurofilament light chain (NF-L), and nestin (Du et al., 2011; Lv et al., 2015).
In our preliminary study (date not shown), the presence of MSCs coexpressing CD90 and nestin was identified in BMPCs, and the alterations of CD90+/Nestin+ cell proportion were observed in BMPCs cultured in vitro. Accordingly, we can speculate that enhancement of neurogenic differentiation potential of MSCs by gelatin during primary culture may have resulted from an increase of CD90+/Nestin+ cell proportion in BMPCs cultured on gelatin.
Conclusion
We confirmed that the enhancement of neurogenic differentiation in MSCs did not result from contamination by proliferated NPCs, but from gelatin during primary culture. Together, these results will more definitively establish the mechanism involved in the transformation of MSCs derived from bone marrow by extracellular matrix proteins.
Footnotes
Acknowledgment
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries (IPET) through the Agri-Bioindustry Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) (IPET112015-4).
Disclosure Statement
No competing financial interests exist.