
Abstract
Salicylic acid (SA) is a plant hormone, which influences several physiological processes, and is a critical modulator of multiple levels of immunity in plants. Several high-throughput screens, which were developed to identify SA-binding proteins through which SA mediates its many physiological effects in plants, uncovered several novel targets of aspirin and its primary metabolite, SA, in humans. These include glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and high mobility group box 1 (HMGB1), two proteins associated with some of the most prevalent and devastating human diseases. In addition, natural and synthetic SA derivatives were discovered, which are much more potent than SA at inhibiting the disease-associated activities of these targets.
Introduction
A
SA is a plant hormone involved in many physiological processes in plants including immunity, where it plays a central role (Vlot et al., 2009). For the past few decades, we sought to uncover SA's mechanisms of action by identifying an array of plant proteins that bind SA, and, as a result, have altered activity. Recently, we developed high-throughput screens to identify additional SA-binding proteins. These screens revealed dozens of novel SA targets in plants (Tian et al., 2012, 2015; Manohar et al., 2015).
Subsequently, we used these screens to investigate whether SA or aspirin exerts its pharmacological activities by targeting proteins besides the cyclooxygenases in humans. This strategy identified several proteins that bind SA, and as a result, exhibit altered activities. One such protein is glyceraldehyde 3-phosphate dehydrogenase (GAPDH), a cytosolic enzyme that plays a central role in the production of energy. In addition to this housekeeping function, other roles of this protein have come to light in the past few decades, including participation in neuronal cell death. Thus, GAPDH is a major suspect in neurodegenerative diseases such as Alzheimer's, Parkinson's, and Huntington's diseases (Chuang et al., 2005). Hara and Snyder (2006) discovered that GAPDH plays a key role in a novel cell death cascade, which involves GAPDH movement into the nucleus, where it enhances degradation of nuclear proteins. This results in neuronal cell death. The anti-Parkinson's disease drug deprenyl inhibits GAPDH movement into the nucleus and the resulting cell death.
SA and Its More Potent Derivatives Bind GAPDH and Inhibit Its Role in Cell Death
We discovered that human GAPDH, like its plant counterpart (Tian et al., 2015), binds SA, which suppresses both GAPDH nuclear translocation and the resulting cell death, much like deprenyl (Choi et al., 2015a). Importantly, we also identified several natural and synthetic derivatives of SA, which bind GAPDH more tightly than SA and thus inhibit its translocation to the nucleus and the resulting cell death more effectively than SA (Fig. 1). Interestingly, the natural derivative amorfrutin B1 is produced by a herb used for thousands of years in traditional Chinese medicine, Glycyrrhiza foetida, known commonly as licorice.
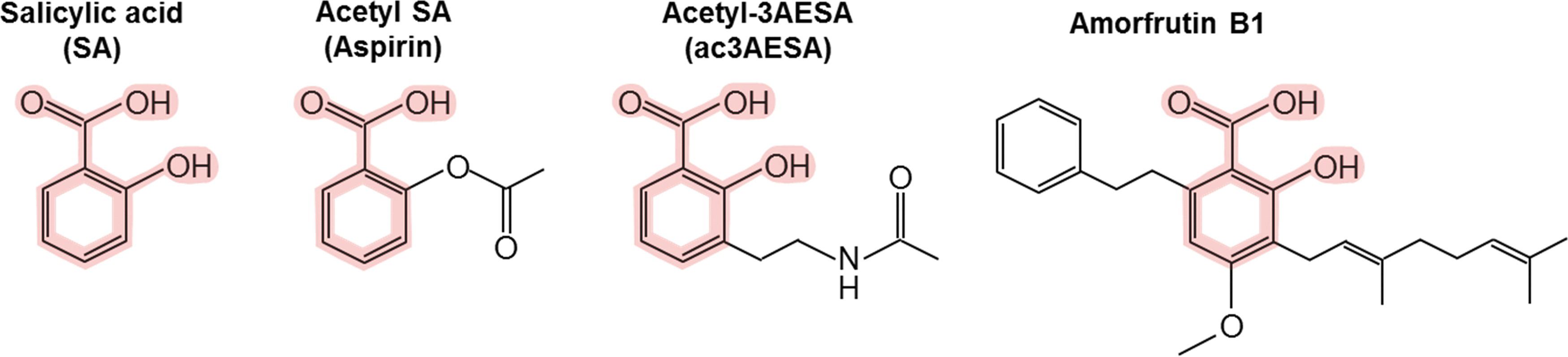
Chemical structures of SA and its synthetic and natural derivatives. The SA core is highlighted in pink. SA, salicylic acid.
SA and Its Derivatives Also Bind HMGB1, Thereby Inhibiting Its Pro-Inflammatory Activities
These screens also uncovered another target that is associated with some of the most prevalent and devastating diseases. It is high mobility group box 1 (HMGB1; Choi et al., 2015b). HMGB1 is the most abundant nuclear protein besides the histones and plays a central role in condensing DNA. Upon release into the extracellular milieu from damaged or necrotic cells, or active secretion by certain immune or cancer cells, HMGB1 becomes highly pro-inflammatory, recruiting immune cells and inducing them to produce pro-inflammatory cytokines (Andersson and Tracey, 2011). The resulting inflammation protects damaged tissue against infection and promotes healing. Unfortunately, sometimes the inflammation is not properly controlled or persists (unresolved), which leads to or is part of the pathogenesis of many inflammation-associated diseases such as arthritis, atherosclerosis, lupus, sepsis, inflammatory bowel disorders, and certain cancers. We discovered that SA binds to HMGB1, thereby blocking its pro-inflammatory activities. It does so at concentrations that are far lower than those required to suppress the enzymatic activity of COX1 and COX2. Moreover, we have discovered that HMGB1 also induces the expression of the gene encoding COX2, and these low levels of SA suppress this induction. Thus, SA does act, in part, through its effect on cyclooxygenase, but it does so by inhibiting the production rather than the activity of this enzyme. Notably, amorfrutin B1 and the synthetic derivative acetyl 3-aminoethyl SA are 40–70 times more potent than SA at blocking HMGB1's pro-inflammatory activities.
Discussion
The discovery that HMGB1's various pro-inflammatory activities are inhibited by low levels of SA provides one likely explanation for the protective effects of low-dose aspirin usage. The more potent SA derivatives also provide proof-of-concept that better SA-based drugs are achievable. It should be noted that a few other aspirin/SA targets have been identified over the past few decades. However, the relevance of these interactions has been called into question because of the very high, nonphysiological levels of aspirin/SA required to alter their activities.
What are the implications of these results for neurodegenerative diseases like Alzheimer's and Parkinson's? Although much more research is clearly needed, our findings hold great promise. Neuroinflammation is a well-established feature of Alzheimer's disease pathogenesis. Although presently its cause is not well characterized, our studies have uncovered three likely culprits and promising means to suppress their deleterious activities. They are HMGB1 and a newly discovered target, both of which have pro-inflammatory activities when released into the extracellular milieu, and GAPDH, whose cell death-mediating activity could facilitate release of these two pro-inflammatory proteins from neuronal cells. Furthermore, since SA and its derivatives also block HMGB1's induction of Cox2 gene expression, another major source of inflammation can be neutralized. These anti-inflammatory activities, together with suppression of GAPDH's role in neuronal cell death by SA-based drugs, suggest that they might be an effective and low-cost treatment, and perhaps will even find utility in prevention of these neurodegenerative diseases. Note that while assessing whether an SA-based drug can help prevent Alzheimer's will require long-term studies, a 2002 study already found that 3,000 individuals, who exhibited early signs of the disease or were likely candidates, showed a 45% risk reduction with 2 or more years of regular use of anti-inflammatory drugs (Zandi et al., 2002). Should SA-based drugs prove effective in either treatment or prevention, the impact would be immense. The number of people worldwide suffering from Alzheimer's and related dementia is estimated to be 25–35 million and growing rapidly.
GAPDH and HMGB1 are only the first two of several novel targets of aspirin/SA in humans that we have uncovered. Another target is a key regulator of energy metabolism, and therefore is an important player in obesity and diabetes. Several additional likely targets require further study, including a protein that, like HMGB1, plays a role in inducing inflammation, as already noted. Further research will likely reveal additional targets of aspirin/SA and will clarify which are responsible for SA's/aspirin's beneficial therapeutic activity, as well as their negative side effects. Given the potentially large number of targets for aspirin/SA, the multiple pharmacological effects of these compounds, and the widespread use of aspirin and/or natural SA derivatives, which our studies suggest are the basis for at least some traditional medicines, it behooves the biomedical community to invest in studies that clarify exactly how aspirin/SA works—that is, elucidating their mechanisms of action. I predict that aspirin/SA will exert its effects in humans through multiple mechanisms of action. In light of our discovery of more than two dozen proteins in plants, through which SA mediates its multiple effects on immunity and several other plant processes, and that most animals eat plants, all of which contain SA as an important hormone, a large number of targets in humans perhaps should not be surprising. Interestingly, many of these SA targets are shared by plants and humans/animals, including the GAPDHs and HMGBs (Fig. 2). For example, both human HMGB1 and its plant counterpart, HMGB3, when released from damaged cells into the extracellular milieu, induce innate immune responses to protect the damaged tissue against infection (Choi et al., 2015b; 2016). SA binds to both and inhibits their immune/inflammatory activities. In summary, SA and its targets appear to be at the crossroads of plant and human health.
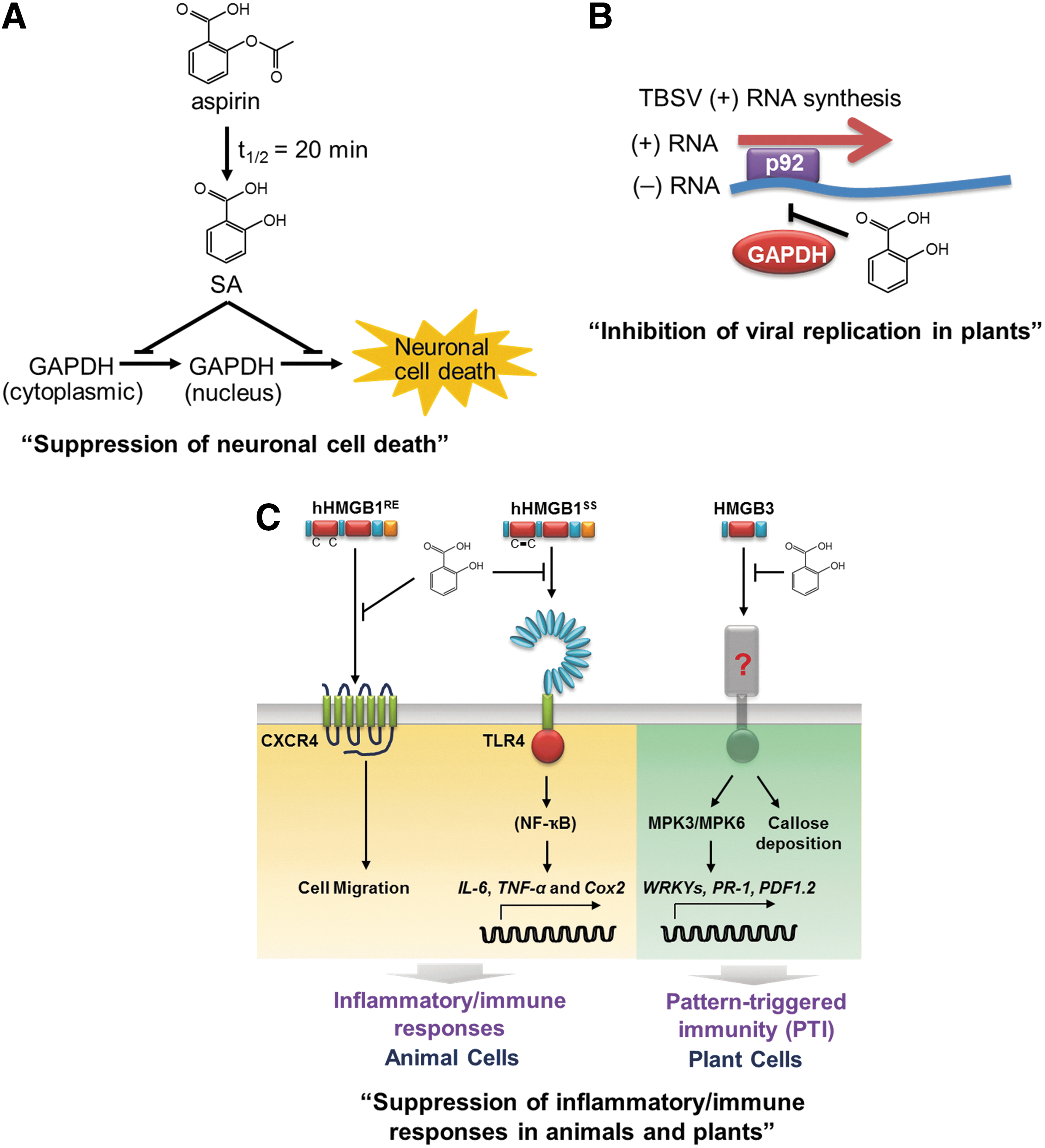
Common targets of SA in plants and animals. In animals/humans, aspirin is rapidly converted to SA.
Footnotes
Acknowledgment
That portion of the work summarized above, which was carried out by the author's group, was funded by the U.S. National Science Foundation Grant Number IOS-0820405.
Disclosure Statement
No competing financial interests exist.