
Abstract
The proliferation and migration of vascular smooth muscle cells (VSMCs) play crucial roles in the pathogenesis of diabetes and its complications. Thymoquinone (TQ) is the primary bioactive component of Nigella sativa L. seed oil, which exhibits antihyperglycemic effect in diabetic rats, but its role in VSMC proliferation and migration has not been investigated. The results of MTT assay and flow cytometry assay indicated that TQ dose-dependently inhibited angiotensin II (Ang II)-induced VSMCs' cell cycle progression, as well as cyclin D1 expression, whereas p21 expression was altered conversely. TQ dose-dependently suppressed Ang II-induced VSMC migration accompanied by reduced MMP-9 expression. In addition, we observed the elevated reactive oxygen species (ROS) generation and NADPH oxidase activity and reduced superoxide dismutase activity in Ang II-treated VSMCs, which were dose-dependently reversed by TQ. Western blot analysis indicated that TQ dose-dependently restored Ang II-inhibited expression of p-AMPK, PPARγ, and peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) proteins. Furthermore, adenosine monophosphate-activated protein kinase (AMPK) inhibitor Compound C and PGC-1α siRNA transfection abrogated the activation of TQ on Ang II-inhibited AMPK/PPARγ/PGC-1α signaling, but abolished the inhibitory effects of TQ on Ang II-induced VSMC proliferation and migration, as well as ROS generation. Taken together, these results demonstrated that TQ inhibited Ang II-induced VSMC proliferation and migration through the AMPK/PPARγ/PGC-1α pathway.
Introduction
C
Thus, inhibition of VSMC proliferation and migration through drugs or other agents is considered of great benefit in the exploration of anti-AS drugs and antidiabetic drugs, as well as the prevention of DM and related cardiovascular complications.
Thymoquinone (TQ) is an active ingredient derived from the medicinal plant Nigella sativa L., which belongs to family Ranunculaceae, and different parts of plant can be used for medicinal purposes to cure various diseases (Tauseef Sultan et al., 2009). It has been investigated for its antioxidant, anti-inflammatory, and anticancer properties in both in vitro and in vivo models since its first extraction. Indeed, there is accumulating evidence that TQ has therapeutic effects in many diseases, including inflammation, cancer, sepsis, AS, and diabetes (Ragheb et al., 2011; Bamosa, 2015; Rajput et al., 2015). Notably, TQ exhibits antihyperglycemic effect in streptozocin (STZ)-induced diabetic rats (Sangi et al., 2015). However, the role of TQ in VSMC proliferation and migration has not yet been investigated.
A more comprehensive understanding of the precise molecular mechanism involved in VSMC proliferation and migration is essential to elucidate the pathological process of DM. Research has showed that DM is associated with enhanced production of free radicals, which further complicates the condition, leading to oxidative stress, cardiovascular abnormalities, renal failure, neurodegeneration, and immune dysfunction (Hung et al., 2012). Peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) is considered to be the main regulator of the expression of various mitochondrial proteins and energy metabolism, and most importantly, participates in the onset of diabetes through regulation of major metabolic tissues (Besseiche et al., 2015).
Interestingly, PGC-1α has been documented to be a broad and powerful regulator of reactive oxygen species (ROS) metabolism, and PGC-1α inhibition leads to elevated ROS levels paralleled with a decrease in antioxidant factors (Cao et al., 2015). Moreover, PGC-1α limits angiotensin II (Ang II)-induced rat VSMC proliferation through attenuating ROS generation (Zhao et al., 2015). Meanwhile, TQ has been found to exhibit anticancer activity through modulation of PPARγ activation and ROS generation (Woo et al., 2012) and ameliorate oxidative stress in experimental hyperglycemic rats (Sankaranarayanan and Pari, 2011). Therefore, TQ may be a promising candidate in inhibiting VSMC proliferation and migration through regulating oxidative stress and PGC-1α-related signaling in the pathological process of DM.
Among various circulatory factors, Ang II is a main pathophysiological peptide for AS and hypertension through induction of VSMC proliferation and migration (Berk et al., 2000). Ang II has various actions on VSMCs, which can influence cell apoptosis, cause oxidative stress, inflammatory reactions, and extracellular matrix deposition, and stimulate production of other growth factors and vasoconstrictors. Accordingly, in this study, we investigated the effect of TQ on Ang II-induced cell proliferation and migration in rat aortic VSMCs, and further, explored the related potential mechanisms.
Materials and Methods
Cell isolaton and cell culture
Male Sprague-Dawley (SD) rats, aged 3–4 weeks, were supplied by the Experimental Animal Center of Xi'an Jiaotong University (Xi'an, China). All animal experimental procedures were approved by the Institutional Ethics Committee for animal experiment of Xi'an Jiaotong University. Rat aortic VSMCs were isolated from the thoracic aorta of SD rats as described previously (Gordon et al., 1986). Cells were maintained in DMEM (Gibco, Rockville, MD) supplemented with 10% FBS (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin (Life Technologies, Rockville, MD) at 37°C in a humidified atmosphere of 95% air and 5% CO2.
All experiments were conducted using VSMCs between passages 4 and 8 that were growth arrested at 70–80% confluence for 24 h with a medium containing 0.5% FBS. Then, the cells were subjected to a medium containing Ang II (0, 10−3, 10−2, 10−1, and 1 μM) or TQ (0, 5, 10, 15 μM) and incubated for the indicated times according to the experimental design. The adenosine monophosphate-activated protein kinase (AMPK) inhibitor, Compound C (Sigma, St Louis, MO), was added at a concentration of 5 μM.
Cell transfection
To knock down PGC-1α expression in VSMCs, double-stranded RNA interference oligonucleotides were designed and synthesized by Genesil (Wuhan, China), which targeted sequences within the rat PGC-1α coding sequence (5′-AAGACGGATTGCCCTCATTTG-3′). Nonspecific oligonucleotides (5′-GACTTCATAAGGCGCATGC-3′) were used as a negative control. For transfection experiment, the VSMCs were transfected with 50 nM PGC-1α siRNA or scrambled siRNA using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions.
MTT assay
Cultured cells (5 × 103 cells/well) after transfection or not were seeded in 96-well plates (Corning, Inc., Corning, NY) overnight and then incubated with DMEM containing 0.5% FBS for 24 h before use. Next, the cells were pretreated with TQ (0, 5, 10, 15 μM) for 1 h and then stimulated with or without Ang II (1 μM; both from Sigma) in the absence or presence of Compound C in DMEM containing 0.5% FBS according to the experiment requirements at 37°C in 5% of CO2. After 24 h, 20 μL of modified tetrazolium salt 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; 5 mg/mL; Sigma) were added to each well and samples were incubated at 37°C for 4 h. Then, the supernatant was carefully removed and 100 μL of dimethyl sulfoxide (Sigma) were added to lyse the cells.
After the dark-blue MTT crystals dissolved, the absorbance was measured at 490 nm using a Benchmark microplate reader (Bio-Rad, Hercules, CA).
Cell cycle analysis
Cultured cells (5 × 105 cells/well) were seeded in six-well plates (Corning, Inc.) and subjected to stimulation of TQ (0, 5, 10, 15 μM) and Ang II (0, 1 μM) as described previously. After 24 h, following the collection and fixation in 70% ethanol overnight at 4°C, cells were stained with propidium iodide (PI; 50 mg/mL; Sigma) and DNase-free RNase A (100 mg/mL) and then incubated at 37°C for 30 min in the dark before analysis. Subsequently, quantitative analysis of DNA content was performed in a FACSCalibur flow cytometer (Becton Dickinson Immunocytometry Systems, San Jose, CA), and the fraction of the cell population in each phase of the cell cycle was determined using the flow cytometry analysis.
Cell migration assay
The cell migration assay was performed using Transwell chambers with fibronectin-coated 8-μm-pore-size polycarbonate membrane (BD Biosciences, Bedford, MA). Then, 600 μL of DMEM containing 0.5% FBS were added to the lower compartment and 100 μL of cell suspension (4 × 104 cells/well) after transfection or not were added to the upper compartment. Then, TQ (0, 5, 10, 15 μM), Ang II (0, 1 μM), or 5 μM Compound C were added to both the upper and the lower chambers as indicated. After 24 h of incubation at 37°C, the nonmigrated cells on the upper membrane were removed by a cotton swab. The migrated cells were fixed with 4% paraformaldehyde and stained with 0.1% Crystal Violet (both from Sigma).
The stained migratory cells were quantified by manual counting under an inverted light microscope (Olympus Corporation, Tokyo, Japan) in at least five randomly selected areas of view and analyzed statistically.
Western blot analysis
Cultured cells (5 × 105 cells/well) after transfection or not were seeded in six-well plates and subjected to stimulation of TQ (0, 5, 10, 15 μM) and Ang II (0, 10−3, 10−2, 10−1, 1 μM) in the absence or presence of Compound C as described previously. After 24 h, the cells were extracted using RIPA lysis buffer (Beyotime, Shanghai, China). The protein concentrations were determined using the Pierce bicinchoninic acid assay (BCA) Protein Assay Kit (Thermo Scientific, Rockford, IL). Equal amounts of protein were processed for Western blot analysis following the standard protocols.
The primary antibodies used were rabbit polyclonal anti-PPARγ antibody, rabbit polyclonal anti-p-AMPK antibody, goat polyclonal anti-PGC-1α antibody, rabbit polyclonal anti-cyclin D1 antibody, rabbit polyclonal anti-β-actin antibody, and rabbit polyclonal anti-p21 antibody (Santa Cruz Biotechnology, Santa Cruz, CA). After incubation with proper secondary antibody, including horseradish peroxidase-conjugated anti-rabbit or anti-goat antibodies (Cell Signaling Technology, Beverly, MA), the resultant protein bands were visualized using an Enhanced Chemiluminescence (ECL) Kit (Beyotime). Quantification analysis was performed using the Gel-Pro Analyzer version 4.0 software (Media Cybernetics, Inc., Silver Spring, MD).
Measurement of ROS generation
ROS generation was measured by the ROS Assay Kit (Beyotime) according to the manufacturer's instructions (Li et al., 2010). Cultured cells after transfection or not were seeded in six-well plates and subjected to stimulation of TQ (0, 5, 10, 15 μM) and Ang II (0, 1 μM) in the absence or presence of Compound C as described previously. After incubation with 2′,7′-dichlorofluorescein diacetate (DCFH-DA), the cells were collected and detected by fluorescence spectrophotometry (Hitachi, Tokyo, Japan) at 488 nm excitation and 525 nm emission wavelengths. The mean densities of the fluorescent images were quantified with Image-Pro Plus 6.0 software (Media Cybernetics, Inc.) from five random fields per well.
Determination of superoxide dismutase and NADPH oxidase activities
Cultured cells were seeded in six-well plates and subjected to stimulation of TQ (0, 5, 10, 15 μM) and Ang II (0, 1 μM) as described previously. The superoxide dismutase (SOD) activity in cell lysates was determined using the SOD Detection Kit (Jiancheng Biological Engineering, Nanjing, China) according to the manufacturer's instructions. For detection of NADPH oxidase activity, lucigenin ECL assay was used as described previously (Chen et al., 2012). Briefly, the total cell lysates were incubated with 5 μM lucigenin (Sigma) for 10 min at 37°C in the dark. After adding NADPH, luminescence was recorded for 180 s and NADPH oxidase activity was calculated as relative light units per 1 mg protein for each minute.
Statistical analyses
All data were expressed as mean ± standard deviation of results derived from three independent experiments performed in triplicate. All analyses were conducted using SPSS software (SPSS, Inc., Chicago, IL). Statistical analysis was performed using Student's t-test and analysis of variance. p < 0.05 was considered statistically significant compared to the respective control.
Results
TQ inhibited Ang II-induced VSMC proliferation and migration
Using the MTT assay, we observed that Ang II incubation elevated the cell proliferation rate at 24 h in VSMCs, which were dose-dependently restored by TQ (5, 10, 15 μM) pretreatment (Fig. 1A). Then, we carried out fluorescence-activated cell sorting analysis of DNA content, the results of which revealed that Ang II led to a significant reduction of cells in the G0/G1 phase of the cell cycle compared to the control group; conversely, TQ pretreatment promoted an accumulation of cells at the G0/G1 phase (Fig. 1C). Western blot analysis showed that Ang II distinctly resulted in an increased expression of cyclin D1 and MMP-9 proteins, as well as a decreased p21 expression, which were reversed by TQ pretreatment dose-dependently (Fig. 1B). Furthermore, TQ dose-dependently reduced the number of migrated VSMCs enhanced by Ang II (Fig. 1D).

Effects of TQ and Ang II on VSMC proliferation and migration. VSMCs were processed as described in Materials and Methods section for 24 h and then processed for respective detection.
Taken together, these findings indicated that TQ markedly inhibited Ang II-induced VSMC proliferation and migration, accompanied by changes in protein levels of cyclin D1, p21, and MMP-9.
TQ attenuated Ang II-induced oxidative stress in VSMCs
Compared with the control group, Ang II treatment remarkably increased ROS generation; whereas, TQ (5, 10, 15 μM) pretreatment dose-dependently attenuated the increased ROS generation induced by Ang II (Fig. 2A). In addition, NADPH oxidase and SOD are the major regulators of intracellular ROS. The SOD activity was reduced and NADPH oxidase activity was enhanced in Ang II-treated VSMCs. However, TQ pretreatment partially reversed both changes in a dose-dependent manner (Fig. 2B, C). These results implied that TQ could significantly attenuate Ang II-induced elevation in oxidative stress in VSMCs.
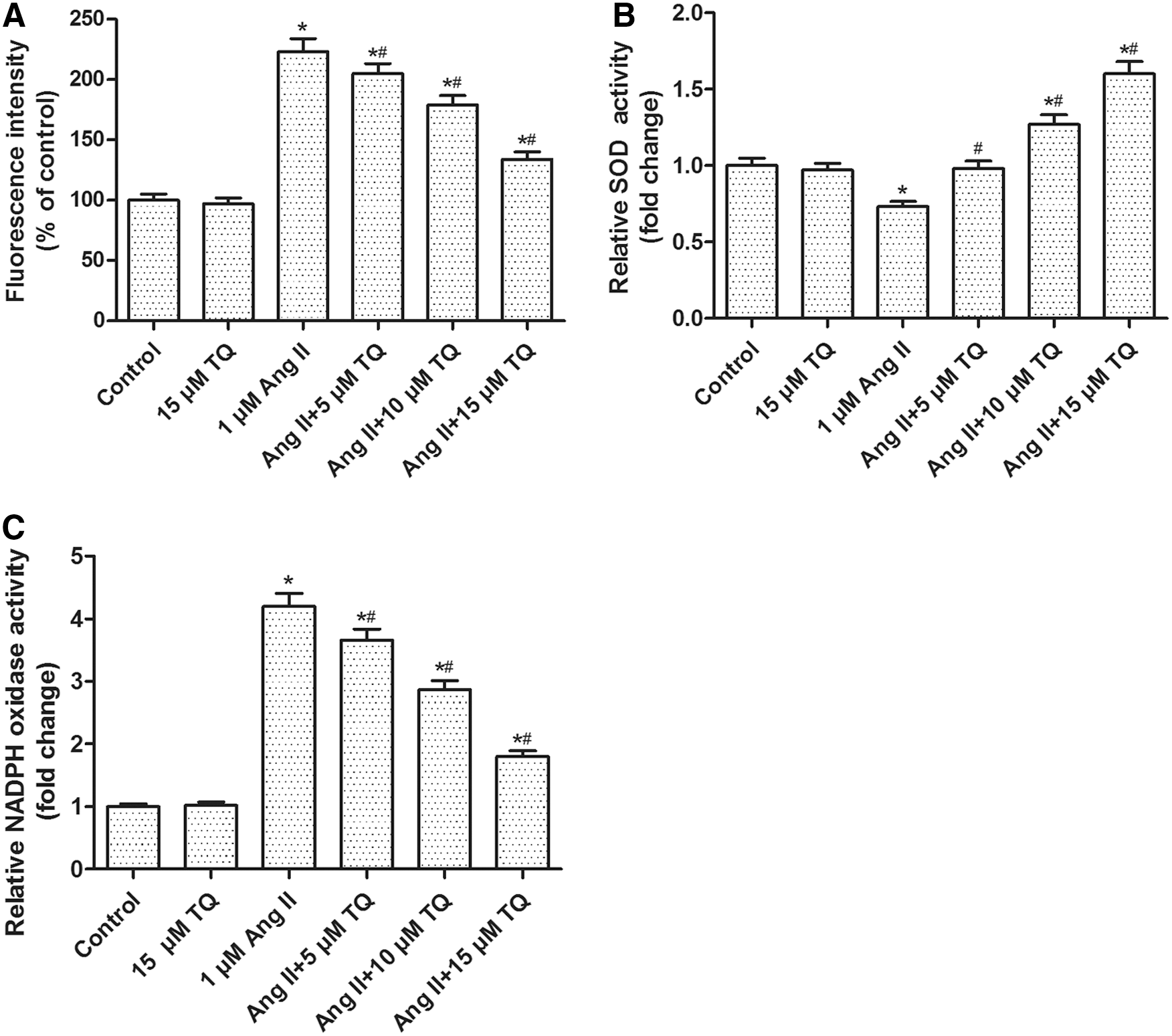
Effects of TQ and Ang II on ROS generation, SOD, and NADPH oxidase activities in VSMCs. VSMCs were processed as described in Materials and Methods section for 24 h and then processed for respective detection.
TQ enhanced Ang II-inhibited AMPK/PPARγ/PGC-1α signaling in VSMCs
Western blot analysis indicated that Ang II (10−3, 10−2, 10−1, 1 μM) inhibited the expression of p-AMPK, PPARγ, and PGC-1α proteins in a dose-dependent manner in VSMCs (Fig. 3A). However, TQ (5, 10, 15 μM) pretreatment dose-dependently enhanced the expression of p-AMPK, PPARγ, and PGC-1α proteins inhibited by Ang II (Fig. 3B). Thus, we speculated that AMPK/PPARγ/PGC-1α signaling might be involved in the process of VSMC proliferation and migration.
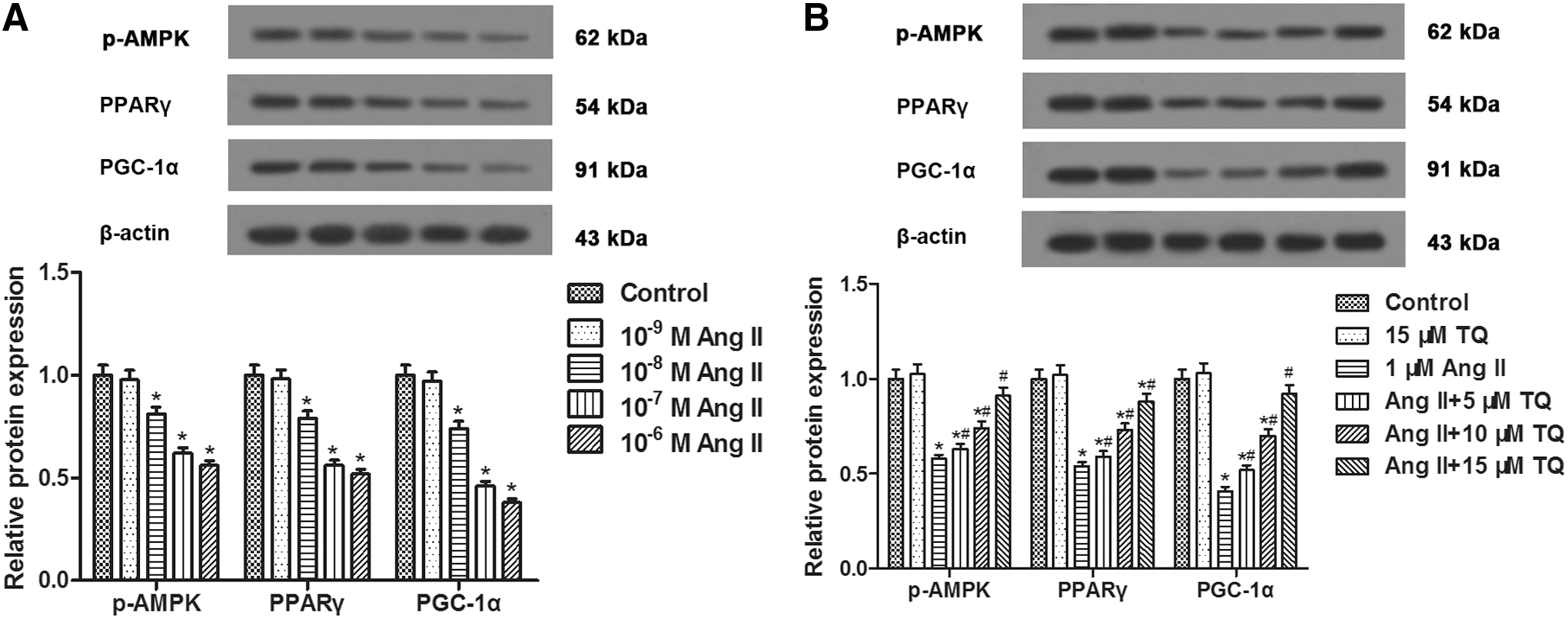
Effects of TQ and Ang II on the expression of p-AMPK, PPARγ, and PGC-1α in VSMCs. VSMCs were processed as described in Materials and Methods section for 24 h and then processed for respective detection.
TQ inhibited Ang II-induced VSMC proliferation and migration through the AMPK/PPARγ/PGC-1α pathway
In the present study, Ang II incubation elevated the cell proliferation and migration, as well as oxidative stress in VSMCs, which were weakened by TQ pretreatment. Inhibition of AMPK with Compound C and inhibition of PGC-1α with siRNA transfection abrogated the activation of TQ on Ang II-inhibited AMPK/PPARγ/PGC-1α signaling (Fig. 4A). In addition, both AMPK inhibitor Compound C and PGC-1α siRNA transfection abolished the effects of TQ on VSMC proliferation and migration (Fig. 4B, C). Likewise, TQ reversed the elevation of ROS generation induced by Ang II, which was abolished by AMPK inhibitor and PGC-1α siRNA transfection (Fig. 4D). Hence, we concluded that TQ inhibited Ang II-induced VSMC proliferation and migration through the AMPK/PPARγ/PGC-1α pathway.
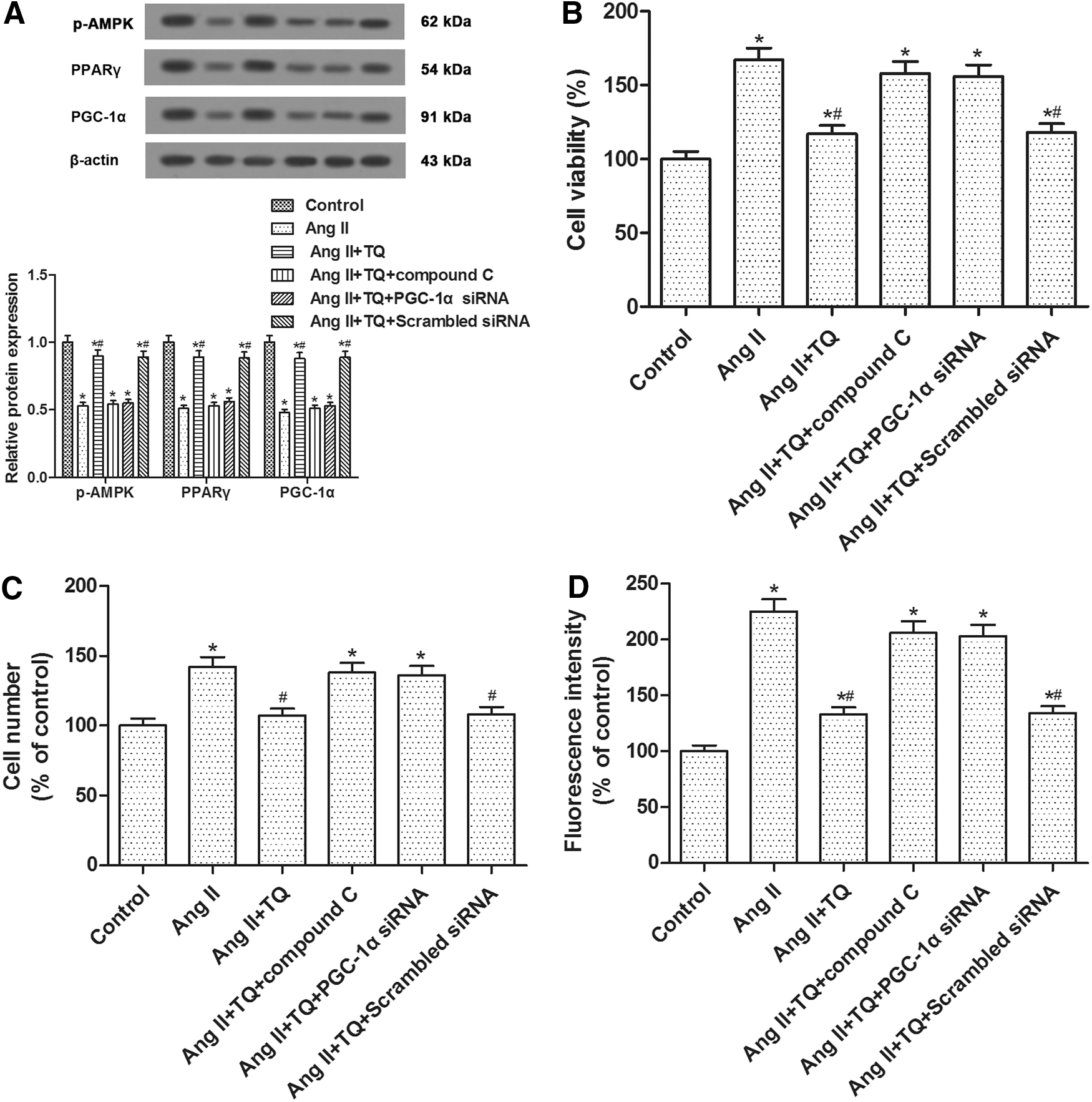
The inhibitory effects of TQ on Ang II-stimulated VSMC proliferation and migration, as well as oxidative stress, were reduced by AMPK inhibition and PGC-1α inhibition. VSMCs were processed with Ang II (1 μM), TQ (15 μM), or Compound C (5 μM) as described in Materials and Methods section for 24 h and then processed for respective detection.
Discussion
With the increasing incidence of type 2 diabetes, cardiovascular risk in individuals with diabetes is increased up to 10-fold. The proliferation and migration of VSMCs have been proved to perform a crucial role in the pathogenesis of diabetic vascular disease. TQ, a known herbal antioxidant, has been demonstrated to contain strong antioxidant properties. Considering this, the present study mainly focused on the antioxidative effect of TQ on the Ang II-induced VSMC proliferation and migration and the possible mechanisms involved.
The proliferation and migration of VSMCs in blood vessels play vital roles in the pathogenesis of vascular disorders such as AS. Increasing evidence suggests that Ang II is the most potent stimulator of VSMC proliferation. Previous studies have indicated that TQ exhibits inhibitory effects on multiple cancers. TQ inhibits proliferation and migration of human non-small cell lung cancer cells (Yang et al., 2015) and reduces migration and invasion of human glioblastoma cells associated with MMP-9 downregulation (Kolli-Bouhafs et al., 2012). Treatment with TQ arrests the cell cycle in G1 by upregulating p21 and downregulating cyclin D1 expression in hepatocellular carcinoma (Ke et al., 2015). Indeed, our results indicated that TQ inhibited Ang II-induced VSMC proliferation and migration, accompanied by changes in protein levels of cyclin D1, MMP-9, and p21.
Some studies demonstrated that Ang II induced VSMC proliferation and oxidative stress, accompanied by upregulation of NADPH oxidase activity and ROS production and downregulation of SOD activity (Chen et al., 2014). NADPH oxidase is a principal factor in Ang II-induced ROS generation, which is an essential contributor to VSMC proliferation and AS (Lusis, 2000). NADPH oxidase and SOD are the major regulators of intracellular ROS (Xu et al., 2013).
TQ acts as a free radical and superoxide radical scavenger and preserves the activity of various antioxidant enzymes. TQ inhibits inflammation-induced ROS generation in osteoblasts (Thummuri et al., 2015) and impairs the phosphorylation of a cytosolic subunit of the NADPH oxidase (Boudiaf et al., 2016). Simultaneous treatment with TQ inhibits the augmented ROS production and suppressed SOD levels (Chandra et al., 2009). Both TQ pretreatment and posttreatment can attenuate ROS generation and increase SOD activity in rats during initiation and promotion of colon carcinogenesis (Jrah-Harzallah et al., 2013). Likewise, our data implied that TQ could attenuate Ang II-induced elevation in oxidative stress in VSMCs.
AMPK is expressed in many tissues, including the heart, skeletal muscle, and brain, and further activated by the increase in the AMP/ATP ratio. AMPK is considered an important regulator of diverse cellular pathways and also a switch for cellular energy levels, the activation of which can be induced by glucose deprivation. AMPK activation might attenuate Ang II-induced VSMC proliferation through the inhibition of cell cycle progression (Kim and Choi, 2010).
In addition, PPARγ belongs to the nuclear hormone receptor superfamily of the ligand-activated transcription factors, and the antiproliferative effect of PPARγ has been reported in multiple cancers. Both cell cycle arrest in human hepatocarcinoma cells (Wang et al., 2007) and abolishment of proliferation of human aortic smooth muscle cells (Heo et al., 2007) can be induced by PPARγ activation through elevating p21 and reducing cyclin D1 expression. Accumulating evidences suggest that AMPK may function as a sensor in response to oxidative stress (Wu et al., 2012), and AMPK activation can enhance PGC-1α expression (Hu et al., 2014). PGC-1α attaches to PPARγ and accelerates the interaction with multiple transcription factors. PPARγ/PGC-1α complex involves in different metabolic activities through regulating various gene expressions.
Meanwhile, PGC-1α is a key regulator of ROS (Zhao et al., 2015) and PGC-1α activation may be associated with reduced oxidative stress (Fernandes et al., 2015). Protective role of PGC-1α in diabetic nephropathy is associated with the inhibition of ROS production (Guo et al., 2015). PGC-1α overexpression inhibited Ang II-induced proliferation and migration, ROS generation, and NADPH oxidase activity in VSMCs (Zhao et al., 2015). PPARγ was reduced after Ang II stimulation in H9C2 cells (Yan et al., 2013).
Moreover, TQ enhanced AMPK phosphorylation against hepatic fibrosis in mice (Bai et al., 2014). TQ may also act as a ligand of PPARγ, and the antitumor effect of TQ may also be mediated through modulation of PPARγ activation (Woo et al., 2011). In this study, we demonstrated that inhibition of AMPK with Compound C and inhibition of PGC-1α with siRNA transfection abrogated the activation of TQ on Ang II-inhibited AMPK/PPARγ/PGC-1α signaling, but abolished the inhibitory effects of TQ on VSMC proliferation and migration, as well as ROS generation induced by Ang II.
Collectively, these results demonstrated that TQ inhibited Ang II-induced VSMC proliferation and migration through the AMPK/PPARγ/PGC-1α pathway. These findings suggest that TQ may be a potential drug candidate to prevent the development of AS in clinical practice and support further investigation of the role of TQ in the prevention and treatment of vascular-related disorders and diabetic complications.
Footnotes
Disclosure Statement
No competing financial interests exist.