
Abstract
Pycnogenol® (PYC) is utilized in the treatment of various diseases ranging from chronic inflammation to circulatory diseases, but its efficacy and functional mechanism in pediatric asthma continue to remain obscure. Therefore, the purpose of this study was to investigate the effectiveness and molecular mechanism of PYC on regulation of asthmatic airway inflammation. We found that PYC with tail intravenous injection of 50 mg/kg or intragastric administration of 100 mg/kg all reduced ovalbumin (OVA)-induced airway injury. Pharmacokinetics of PYC was evaluated by high-performance liquid chromatography assay, indicating that PYC was quickly absorbed into the blood after intragastric administration, and PYC metabolism was later improved gradually with increase of time after PYC administration. PYC has a higher bioavailability of 71.96%, and it was more easily absorbed by the body. PYC inhibited the number of total inflammatory cells and levels of interleukin (IL)-4, IL-5, IL-9, and IL-13 in bronchoalveolar lavage fluid of OVA-induced mice. PYC inhibited IL-13 secretion from the Th2 cells, thereby causing a reduction in expression of the signaling molecules in JAK/STAT6 pathway in airway epithelial cells. STAT6 silence suppressed IL-13-increased acetylcholine level. STAT6 overexpression promoted expression of goblet cell metaplasia-associated molecules (FOXA3, SPDEF, and Muc5ac). PYC suppressed OVA-induced expression of FOXA3, SPDEF, and Muc5ac in lung. Our findings indicate that PYC has a higher bioavailability and it prevents emergence of OVA-induced airway injury and airway inflammation in mice by inhibiting IL-13/JAK/STAT6 pathway and blocking release of acetylcholine to reduce goblet cell metaplasia.
Introduction
A
The occurrence of asthma involves multiple factors, including infection, allergy, familial inheritance, and environmental conditions etc. Although the inducement of asthma is diverse, the basic pathological symptom of asthma is primarily considered as airway inflammation, in which adult asthma is similar to pediatric asthma (Aligne et al., 2000). Over the past two decades, aerosol inhalation corticosteroids (ICS) can effectively control airway inflammation and reduce airway hyperresponsiveness, and they are generally accepted as the first-line choice of anti-inflammatory therapy in the treatment of asthma (Lipworth, 1999). However, approximately one-third of asthma patients have strong concerns about the adverse effects of the ICS at present, including not only clinical symptoms of ICS side effects, but also the unfavorable long-term dependence to ICS (Horne, 2006). To date, the medicines available as treatment are insufficient to meet the needs of all the patients with asthma; therefore, new medications are required.
Pycnogenol® (PYC), a standardized bioflavonoid extract from the bark of the French maritime pine (Pinus maritime), contains ∼70% procyanidins, which exhibit antioxidant, anti-inflammatory, anticarcinogenic, and immunomodulatory effects. It has been traditionally used in the treatment of inflammatory disease in Europe and North America (Maimoona et al., 2011). This herbal medicine is utilized across the world as an herbal remedy to treat various diseases, ranging from chronic inflammation to circulatory diseases (D'Andrea, 2010; Lee et al., 2011). Approximately 60% of subcutaneous edema patients treated with PYC experienced complete disappearance of the edema and pain at the end of the treatment, while almost all the patients reported a reduction in leg heaviness, which disappeared in ∼33% of the patients (Arcangeli, 2000).
Schoonees's study exposed the therapeutic function of PYC in adults or children with any chronic disorder (Schoonees et al., 2012). Prior studies have also revealed that PYC supplementation is associated with a decrease in platelet aggregation (Araghi-Niknam et al., 2000), lowering the low-density lipoprotein cholesterol, and improving hypertension (Devaraj et al., 2002). Therefore, PYC has positive and extensive effectiveness on diverse diseases. However, thus far, only a few studies are concerned with the functional mechanism of PYC.
Therefore, in this study, we evaluated absorption and mechanism of PYC in vivo and investigated the effects and mechanism of PYC on asthmatic airway injury and inflammation.
Materials and Methods
Mice and treatment
Male infant Balb/c mice (9–10 g) were purchased from the experimental animal center, Xi'an Jiaotong University Colleges of Medicine. All the mice were housed in specific pathogen-free conditions, and all experiments were performed as per the approval of the institutional animal care and use committee.
Mice were sensitized on each of seven alternate days through an intraperitoneal injection of 10 μg ovalbumin (OVA, grade II; Sigma-Aldrich) added in 0.5 mL of phosphate-buffered saline (PBS) buffer (pH 7.4) (Nakae et al., 2003). Mice employed as control were injected with equal PBS. Four weeks after the final injection, mice were challenged with 20 μL of either PBS (control) or PBS containing 100 μg of OVA through intranasal administration eight times for 8 days (Nakae et al., 2003). Control and asthmatic mice were given PYC (Sigma-Aldrich) through tail intravenous injection (20, 30, 50 mg/kg) and intragastric administration (50, 100, 150 mg/kg) 1 h before the OVA challenge (Shin et al., 2013). The airway injury was evaluated 24 h after the last treatment using a respiratory rate tracer (Godart).
Airway injury indices detection
The airway resistance (R L), dynamic lung compliance (C dyn), and enhanced pause (Penh) were detected using a respiratory rate tracer (Godart). Mice were anesthetized with 1% pentobarbital sodium and then performed tracheal intubation; the tracheal catheter was connected with the respiratory rate tracer to detect the tidal volume (V T) and airway flow (V). Then, the distal tracheal catheter was inserted into the middle and lower part of the pleural cavity, the proximal one of tracheal catheter was connected with a pressure transducer (Medical Electronic Instruments Factory of Shanghai) to detect the transpulmonary pressure (Ptp). The R L and C dyn were expressed as: R L (cmH2O × s/mL) = Ptp/V and C dyn (mL/cmH2O) = V T/Ptp. The Penh was monitored using the pulmonary function detector (Buxco). All the mice were given aerosol inhalation methacholine at a dose level of 0, 3.125, 6.25, 12.5, 25, 50, and 100 mg/mL. The Penh values were recorded.
High-performance liquid chromatography assay
High-performance liquid chromatography (HPLC) analysis was carried out on a Hewlett Packard 1100 system equipped. The column, a Nova-Pak C18 (Waters Corp.) column (4.6 × 250 mm), with a 4 μm particle size was used and the temperature maintained by the column oven set at 30°C. The injection was by means of an autosampler and the volume injected was 30 μL. Elution (0.5 mL/min) was performed using a gradient solvent system composed of solvent A (water:methanol:5 M HCl, 79.9:20:0.1) and solvent B (acetonitrile). The mobile phase consisted of 95% solvent A for 10 min; then decreased linearly to 50% solvent A at 50 min, back to 95% solvent A at 55 min; and held at these conditions for a further 5 min.
Authentic standard solutions were analyzed as precision controls by being randomly placed with the samples as unknowns. The monitoring wavelength for the chromatograms was detection at 320 nm. Peak identification of each component was by retention time and spectral characteristics of each peak by photodiode array detection from 200 to 600 nm. Confirmation was achieved by spiking samples with authentic standards.
Bronchoalveolar lavage fluid
Euthanasia was performed on mice within 48 h after the last treatment through intraperitoneal pentobarbital (50 mg/kg; Hanlim Pharm. Co.). Bronchoalveolar lavage fluid (BALF) samples were obtained as previously described (Chen et al., 2008). Briefly, five separate 30 mL aliquots of 0.9% sterile saline were instilled in the most affected lobe radiographically, or the right middle lobe or left lingula, then aspirated and pooled. BALF was immediately centrifuged at 14,000 g for 20 min at 4°C and cell-free supernatant was aliquoted and stored at −80°C. The total inflammatory cell numbers were determined by counting the cells in at least five squares of a hemocytometer.
Airway inflammatory cytokines detection
The levels of the inflammatory factors (interleukin [IL]-4, IL-5, IL-9, and IL-13) in the BALF and immunoglobulin E (IgE) in serum were measured using the ELISA Kits (Sigma-Aldrich).
The count of IL-13-positive cells
Cells were fixed with 4% neutral paraformaldehyde and permeabilized with saponin. Cells were stained with fluorochrome-conjugated anti-mouse IL-13 (eBioscience) and counted under an optical microscope (Olympus).
Isolation and treatment of airway epithelial cells in mice
As described by Lane et al. (2005), the airway epithelial cells were isolated and characterized by Hematoxylin and Eosin staining. The D10.G4.1 (Th2 cell lines, mouse) was obtained from the American Type Culture Collection. All the cells were cultured in DMEM (Invitrogen) supplemented with 10% fetal bovine serum (Invitrogen), 100 U/mL penicillin, and 100 mg/mL streptomycin (Invitrogen), and maintained at 37°C in a humidified incubator using 5% CO2.The D10.G4.1 cells and isolation airway epithelial cells were cultured, respectively, in DMEM with the presence or absence of 100 μg/mL OVA for 72 h (Zuany-Amorim et al., 2002). The cells were then incubated with 100 μg/mL PYC for 24 h (Shin et al., 2013). Supernatants were then collected and cytokine levels determined by enzyme-linked immunosorbent assay (ELISA).
The transfection of overexpressed vector
The pcDNA3.1(+)/STAT6 expression vector was constructed by cloning a STAT6 fragment from normal mouse cDNA into pcDNA3.1(+) (Invitrogen) between BamHI and EcoRI sites to express STAT6 in abundance in Escherichia coli DH5α cells. The primers for STAT6 were as follows: forward primer: 5′-gga CGG ATC CG C TGT CCT TCC TGC CAT ATC-3′, reverse primer: 5′-ctc GGA ATT CCT GTT CTG TGC GGT CAT AC-3′. The recombinant plasmid was identified by endonuclease digestion and DNA sequencing. Then, the pcDNA3.1(+)/STAT6 was transfected into isolated airway epithelial cells using Lipofectamine 2000 (Invitrogen). The stably transfected clones were screened by G418 resistance.
Detection of acetylcholine concentration
The STAT siRNA was transfected into isolated airway epithelial cells using Lipofectamine 2000 (Invitrogen). The primers of STAT siRNA_94: 5′-GGA GUC UAG UUU CCA AGA UdTdT-3′ (F), 5′-AUC UUG GAA ACU AGA CUC CdTdT-3′ (R). We used the Choline/Acetylcholine Quantification Kit (Sigma-Aldrich) to measure acetylcholine concentration after IL-13 stimulation.
Reverse transcription-polymerase chain reaction
Total RNA was extracted using TRIzol reagents (Invitrogen) from the lung and airway epithelial cells, respectively. Isolated RNA was electrophoresed on 1% agarose gel to detect the purity of the total RNA. The first-strand cDNA was synthesized using 1 μg total RNA and SuperScript® III Reverse Transcriptase (Invitrogen). Polymerase chain reaction (PCR) amplification was performed using the PCR Amplification Kit (TaKaRa Biotechnology). The specific primers were designed using Primer Premier 6.0 software and synthesized by Sangon Biotech. The PCR production was electrophoresed on 1% agarose gel and visualized by the Gel Imaging System of Bio-Rad Corp (Bio-Rad). Each band was analyzed by Quantity One 4.62 software (Bio-Rad). The primers for FOXA3 (mouse) were 5′-TTG AGA ACG GCT GCT ATC-3′ (forward) and 5′-TAA GGT GTG GAC GAA GGA-3′ (reverse). The primers for SPDEF (mouse) were 5′-CCA TCC GCC AGT ATT ACA A-3′ (forward) and 5′-AGC AGA GAC CAG TGA CAT-3′ (reverse). The primers for Muc5ac (mouse) were 5′-CTG TCA CCG AGA GGA GAT-3′ (forward) and 5′-CAG TGG TAG ATG TAG ATA GAG T-3′ (reverse). GAPDH served as the housekeeping gene.
Western blotting
Total protein was extracted using the Tissue or Cell Total Protein Extraction Kit (AMRESCO) from the airway epithelial cells. All the primary antibodies were purchased from Abcam (Abcam). The proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis followed by electrotransfer to the nitrocellulose filter membrane. The membranes were probed using antibodies against IL-4R (1:1000), JAK1 (1:1000), STAT6 (1:200), and phosphor (Y641)-STAT6 (1:1000) followed by a horseradish peroxidase-conjugated secondary antibody (Abcam). The nuclear proteins were extracted using the Nucleoprotein Extraction Kit (QIYBO Biological Technology). Bands were revealed with the ECL reagent (Millipore) and recorded on the x-ray films (Kodak). The densitometry of each band was quantified by the gel imaging system and Quantity One 4.62 software (Bio-Rad).
Data and statistical analysis
Data are presented as mean ± standard deviation. Statistical analysis was performed with SPSS13.0 software (IBM). Statistical differences between the means were calculated using one-way analysis of variance where appropriate, followed by LSD multiple comparison tests. Differences were considered significant at p < 0.05.
Results
PYC suppresses OVA-sensitized airway injury in vivo
Male Balb/c mice were sensitized with OVA, then airway injury indices, including the R L (situation of airway obstruction), C dyn (index of lung elasticity), and Penh values, were detected. The results showed that R L was markedly increased (p < 0.01) and C dyn was dramatically reduced (p < 0.01) in OVA-sensitized rats compared with control (Fig. 1A, B). Penh represents asthmatic airway hyperresponsiveness. The methacholine stimulated and then caused smooth muscle contraction, to detect degree of bronchial stenosis used in examination of airway hyperresponsibility (Hargreave et al., 1982). As shown in Figure 1C, with the increasing concentration of methacholine, the Penh value in the OVA-sensitized rats showed a prominent increase in a dose-dependent manner. These results show that OVA had successfully induced asthmatic model.
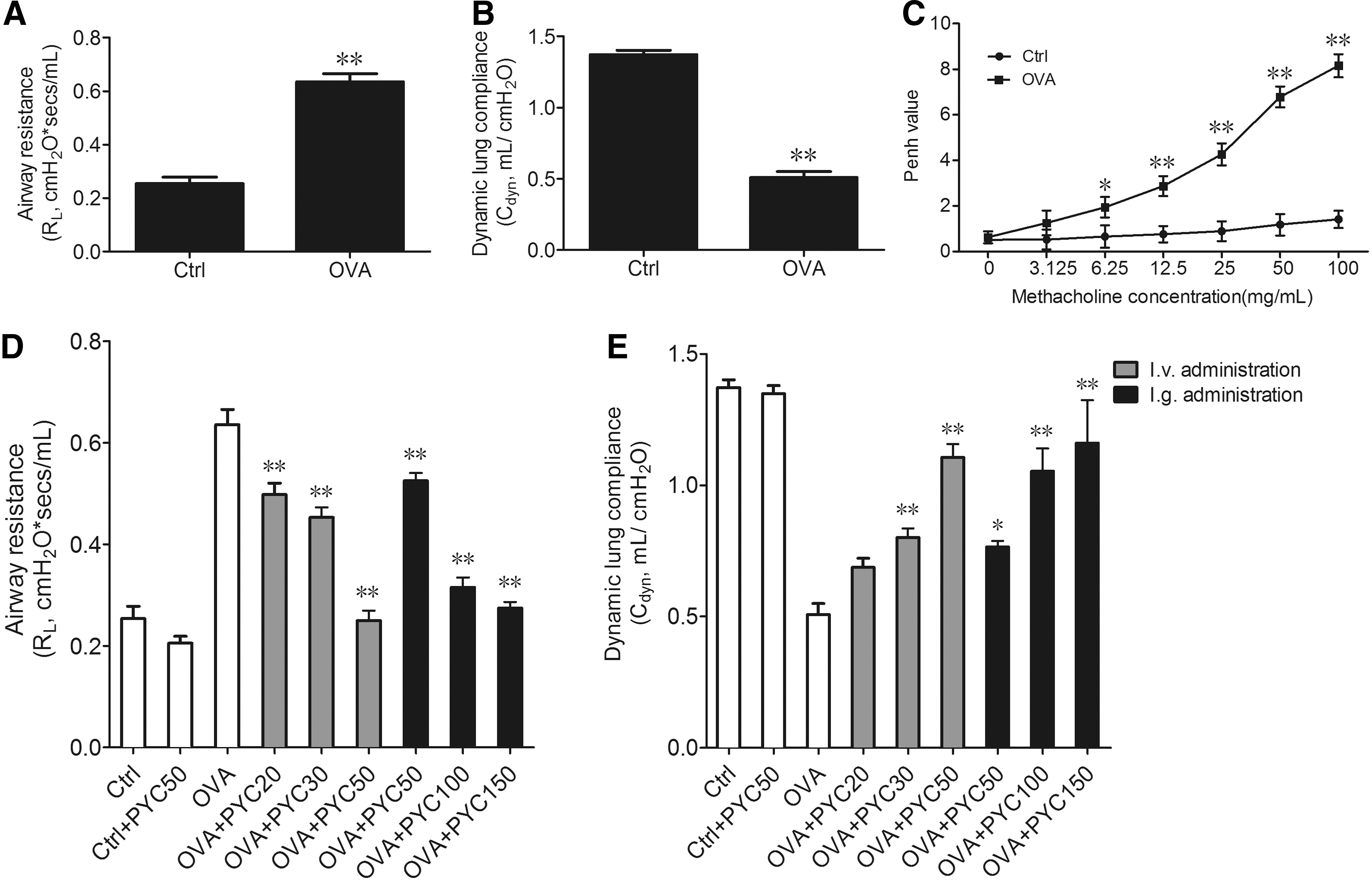
PYC suppressed OVA-sensitized airway injury in vivo
Next, OVA-sensitized rats were treated by PYC with tail intravenous injection (20, 30, 50 mg/kg) and intragastric administration (50, 100, 150 mg/kg). After PYC treatment, airway injury was evaluated by R L and C dyn, results demonstrated that PYC treatment with intravenous (i.v.) administration, especially 50 mg/kg concentration, significantly reduced OVA-induced R L and improved OVA-induced C dyn. PYC treatment with intragastric administration, especially 100 mg/kg concentration due to no obvious difference between 100 and 150 mg/kg concentration, also reduced OVA-induced R L and enhanced OVA-induced C dyn (Fig. 1D, E). Therefore, we chose 50 mg/kg i.v. administration and 100 mg/kg i.g. administration of PYC to investigate absorption and metabolism of PYC in vivo in the next experiments.
Pharmacokinetics of PYC
Ferulic acid is one of complex PYC components and commonly considered as a tracking molecule to evaluate availability of PYC (Virgili et al., 2000). We used HPLC assay to evaluate absorption and metabolism of PYC in OVA-sensitized rats in vivo. The results indicated that PYC concentration in plasma had a sharp reduction and ferulic acid concentration in urine was gradually increased with the increase of time during 50 mg/kg of i.v. administration (Fig. 2A, B), indicating that PYC of plasma has a faster clearup, which is connected with increase of PYC metabolism after i.v. administration. After 100 mg/kg of i.g. administration, PYC concentration in plasma presented a rapid increase, reached peak at 0.6 h after PYC treatment, and was later reduced. Accordingly, ferulic acid concentration in urine exhibited a low ebb within 2 h after PYC treatment, and was later improved gradually (Fig. 2C, D), indicating PYC is quickly absorbed into the blood after i.g. administration, and PYC metabolism is later improved gradually.
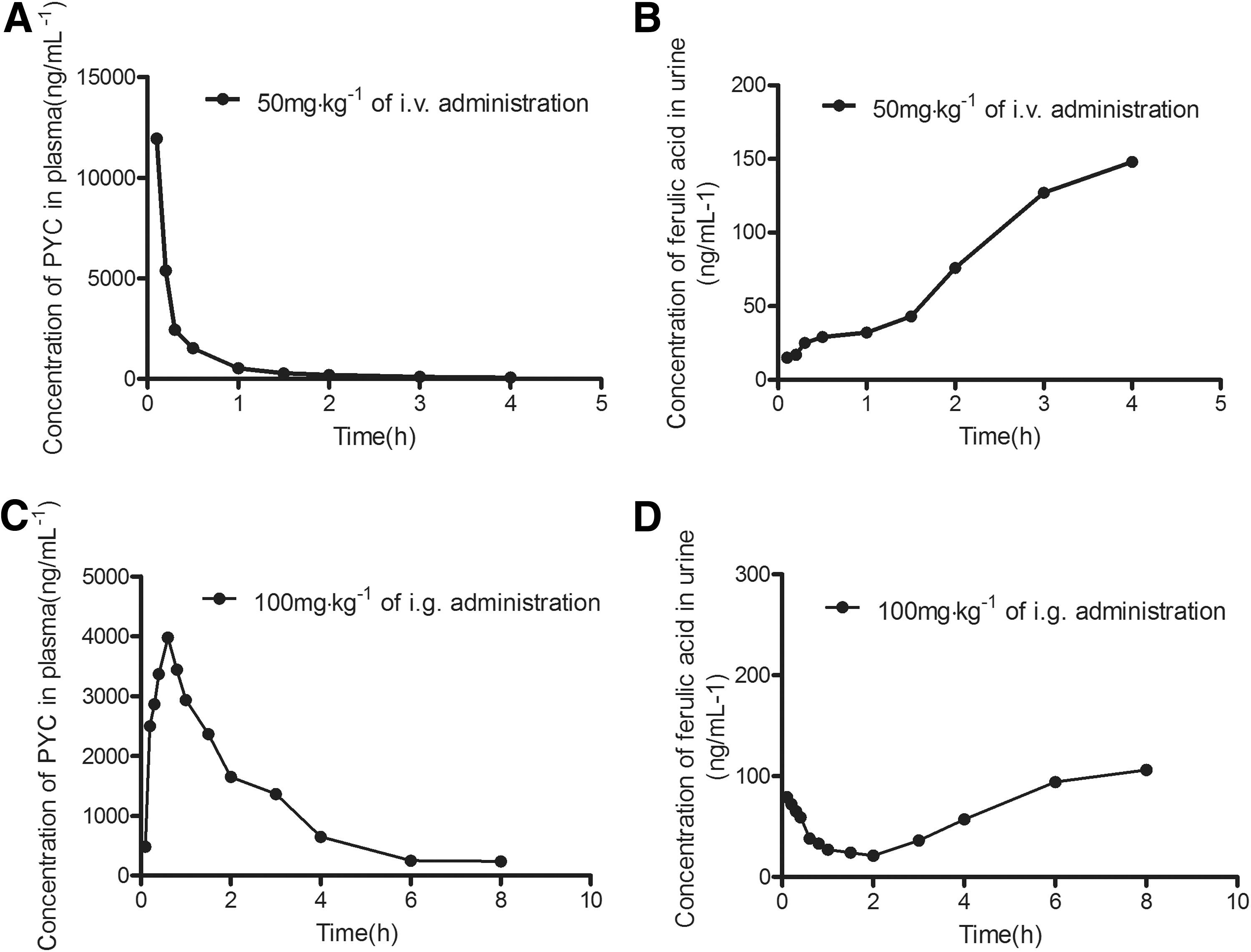
Pharmacokinetic parameters of PYC in plasma and ferulic acid in urine are displayed in Table 1. We compared difference of the two treatments to identify which one is better beneficial for absorption and mechanism of PYC. As shown in Table 1, the T 1/2 of PYC in i.g. administration was almost twofold compared with it in i.v. administration. Area under the curve (AUC0–4h/8h), mean retention time, total clearance, and the apparent volume of distribution at steady state of PYC and ferulic acid in i.g. administration were all higher than those in i.v. administration. Therefore, we considered that PYC treatment with 100 mg/kg of i.g. administration is relatively effective and is the appropriate dose and administration method in this study. Additionally, the bioavailability of PYC is 71.96%, indicating that PYC is more easily absorbed by the body.
PYC, Pycnogenol®, i.g., intragastric; i.v., intravenous; SD, standard deviation; C max, peak concentration of drug; T max, peak time of drug; T 1/2, half-life period; Cl, total clearance; MRT, mean retention time; Vss, the apparent volume of distribution at steady state; AUC0–4h/8h, area under the curve of drug time; F, bioavailability; F (%) = (AUC i.g./Dose i.g.)/(AUC i.v./Dose i.v.) × 100%.
PYC inhibits OVA-induced airway inflammation in vivo
The total number of inflammatory cells in BALF were elevated by OVA stimulation compared with the control (p < 0.01), but they were markedly decreased after PYC treatment (p < 0.01, Fig. 3A). Next, the inflammatory factors in BALF were detected by ELISA. Results show that OVA-induced mice presented a marked elevation of IL-4, IL-5, IL-9, and IL-13 levels compared with the controls. In contrast, levels of IL-4, IL-5, IL-9, and IL-13 were significantly reduced after PYC treatment (Fig. 3B). In general, PYC suppressed OVA-induced secretion of proinflammatory cytokines in vivo.

PYC suppressed OVA-induced airway inflammation in BALF in vivo.
PYC suppresses activity of IL-13-related signaling pathway in vitro
According to above experiments, we found that IL-13 had a strong response to the OVA stimulation, and it was more significant that PYC treated to the level of IL-13 induced by OVA. Therefore, to further explore the mechanism of PYC regulation on IL-13 secretion, the purchased Th2 cells were stimulated by OVA and then treated with PYC. As shown in Figure 4A, the IL-13 level in the OVA-induced Th2 cells presented an observable increase compared with it in control (p < 0.01), and this process was inhibited by PYC stimulation. Meanwhile, the number of IL-13-positive cells was counted by microscope. The results showed that the numbers of IL-13-positive cells in the OVA-induced Th2 cells were elevated compared with it in control (p < 0.01), and this elevation was inhibited by PYC stimulation (Fig. 4B). IL-13 is a critical mediator of airway inflammation and contributes directly to airway hyper-reactivity (Steenwinckel et al., 2009). Therefore, IL-13 and its related signaling pathway would be a potential mechanism for exploring methods of asthmatic treatment.

PYC inhibited activation of OVA-induced IL-13 signaling pathway in vitro.
In subsequent experiments, we investigated whether PYC was involved in the IL-13-related JAK/STAT6 pathway activation in the airway epithelial cells. As depicted in Figure 4C and D, the IL-4R, JAK1, STAT6, and p-STAT6 levels were increased by OVA compared with control, and this increase was suppressed by PYC treatment in airway epithelial cells. In addition, OVA markedly enhanced the expression of nucleic STAT6 protein (p < 0.05), and this process was inhibited by PYC treatment (Fig. 4C, E). These data suggest that PYC inactivates IL-13/JAK1/STAT6 signaling pathway, resulting in blockade of translocation of STAT6 from cytoplasm to nucleus.
Cholinergic nervous system is implicated in PYC-regulated goblet cell metaplasia in vitro
Several studies demonstrated that FOXA3, SPDEF, and Muc5ac played important roles in the regulation of the transcriptional network critical for goblet cell metaplasia in the mouse asthma models (Lambrecht and Hammad, 2012; Ren et al., 2013). To investigate further the effect of PYC on the goblet cell metaplasia, we extracted total RNA from the airway epithelial cells to examine the transcriptional activities of the marker molecules of goblet cell metaplasia. The result showed that the FOXA3, SPDEF, and Muc5ac expressions in the OVA-sensitized cells were notably enhanced compared with the control. After treatment with PYC, a massive reduction in the expression of these molecules was observed compared with the airway epithelial cells of OVA-sensitized mice (Fig. 5A). The result demonstrated that PYC prevented goblet cell metaplasia in vitro.
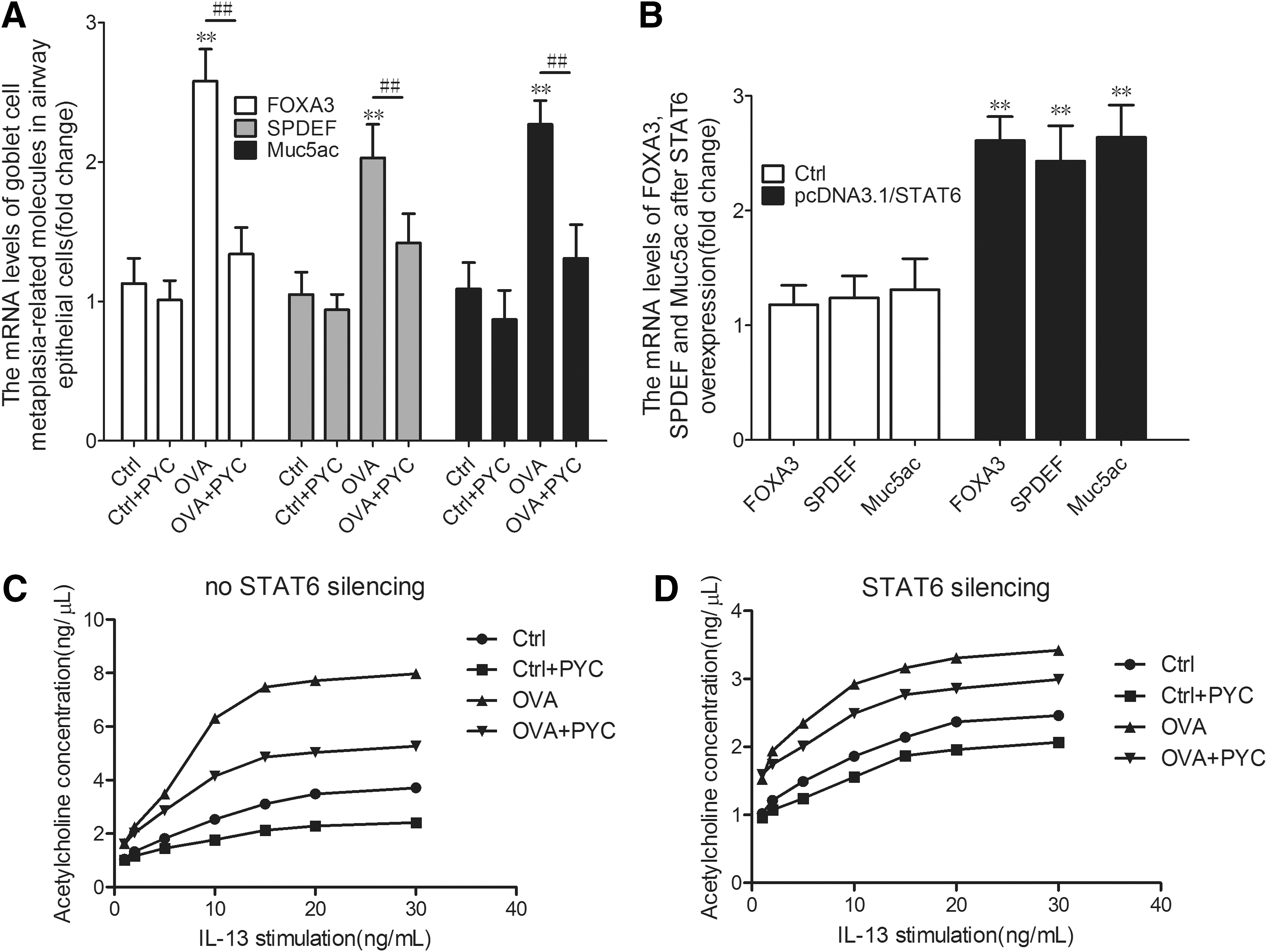
Cholinergic nervous system is implicated in PYC-regulated goblet cell metaplasia in vitro.
Furthermore, to investigate the relationship between JAK/STAT6 pathway and goblet cell metaplasia, we transfected pcDNA3.1/STAT6 vector into isolated airway epithelial cells and detected transcriptional activities of FOXA3, SPDEF, and Muc5ac by reverse transcription-polymerase chain reaction. The results demonstrated that overexpressed STAT6 dramatically elevated mRNA levels FOXA3, SPDEF, and Muc5ac in airway epithelial cells (Fig. 5B). Therefore, PYC suppresses goblet cell metaplasia by blocking IL-13/JAK1/STAT6 signaling pathway.
Stimulation of the cholinergic nervous system induces secretion of mucus from goblet cells (Lundgren and Shelhamer, 1990). Therefore, we conjectured that cholinergic nervous system may be downstream of IL-13/JAK1/STAT6 signaling pathway in regulation of mucus secretion of goblet cells. We used IL-13 with different concentrations to stimulate cells of each group, then the level of acetylcholine was detected. As shown in Figure 5C, acetylcholine concentration had a sharp increase of IL-13 in a dose-dependent manner in OVA group compared with control. After PYC treatment, enhancement of acetylcholine concentration was obviously inhibited, indicating that there is a positive relationship between IL-13 and acetylcholine in OVA-induced airway epithelial cells, and PYC inhibits signal transduction between them.
In fact, we found that not just IL-13, but IL-13/JAK1/STAT6 signaling pathway was implicated in regulation of acetylcholine on mucus secretion. After STAT6 silence, acetylcholine concentration was downregulated in each group, demonstrating that there is a positive relationship between STAT6 and acetylcholine. Overall, these data indicated that IL-13/JAK1/STAT6 signaling pathway promotes mucus secretion from goblet cells by activating cholinergic nervous system.
Discussion
PYC consists of a concentrated solution of water-soluble polyphenols, which includes the bioflavonoids, catechin and taxifolin, as well as phenolcarbonic acids. These antioxidant ingredients enable PYC to play diverse roles in different metabolic processes. The study of pharmacokinetics of PYC has demonstrated that the compounds, including catechin, caffeic acid, ferulic acid, Taxifolin, and the metabolite M1 [delta-(3,4-dihydroxy-phenyl)-gamma-valerolactone], were detected in total plasma of 11 volunteers who received a single dose of 300 mg PYC (Grimm et al., 2006). Moreover, the compounds derived from the PYC were rapidly absorbed. According to our study, PYC treatment with both 50 mg/kg i.v. administration and 100 mg/kg i.g. administration were well absorbed into blood, and the cleanup of PYC in plasma and mechanism of PYC were much quicker.
Generally, PYC containing water-soluble polyphenols exhibited strong hydrophilicity, thus it is difficult to cross the biomembrane tissue. However, PYC has a higher bioavailability of 71.96%, thus we speculated that other mechanism may be involved in absorption and mechanism of PYC, which needs to be explored further. Currently, studies have demonstrated the inhibitory effects of PYC on airway inflammation in OVA-induced asthma (Shin et al., 2013) in the experiment on animals; PYC decreased the inflammatory cell count and the IL-4 and IL-5 levels in BALF. This indicated that PYC attenuated the airway inflammation and mucous hypersecretion induced by the OVA challenge (Shin et al., 2013). However, the molecular mechanism by which PYC regulates the airway inflammation and mucus secretion, these intensive problems remain poorly understood.
IL-13 is an immunoregulatory cytokine secreted predominantly by activated Th2 cells. Allergens cause the differentiation of Th2 cells, which then secrete Th2 cytokines, such as IL-4, IL-5, and IL-13. These cytokines are responsible for IgE production, the development of airway high reactivity, eosinophil accumulation, and mucus hypersecretion (Grimm et al., 2006). The study shows that IL-5 deficiency abolishes eosinophilia, airways hyperreactivity, and lung damage in OVA-induced asthma model (Foster et al., 1996). Moreover, Hamid et al. (1991) used specific hybridization signals of IL-5 mRNA to explore localization of IL-5, and results found that IL-5 mRNA was localized in the bronchial mucosa of asthmatics and regulated eosinophil function in bronchial asthma. Besides, IL-9 and IL-9 receptor are strongly related to a calcium-activated chloride channel in regulation of bronchial epithelium on mucus production in asthmatic patients (Toda et al., 2002). However, the detailed mechanism of regulation of IL-5 and IL-9 on asthmatic progression still needs to be further investigated.
Over the past several years, it has become evident that IL-13 is a key mediator in the pathogenesis of airway inflammation (Hershey, 2003). Also, IL-13 and IL-4, acting through a shared receptor, have been demonstrated to act directly on the airway epithelium to increase the number of goblet cells in vitro and in vivo (Grunig et al., 1998; Dabbagh et al., 1999; Atherton et al., 2003). Although the study of the relationship among IL-13, airway inflammation, and goblet cells has faintly emerged, the exact molecular mechanisms of IL-13 pathway interaction with PYC are still unclear. To illustrate the problems mentioned above, we established the OVA-induced asthmatic mice model and then treated with PYC. In the OVA-induced mice, we detected pulmonary dysfunction and severe airway inflammation, which indicated that the asthmatic mice model had been successfully established. Moreover, PYC treatment in OVA-induced mice alleviated the symptoms of pulmonary dysfunction and airway inflammation, manifested as a reduction in the R L and C dyn, and a drop in the levels of the inflammatory cytokines.
PYC is able to decrease OVA-induced IL-13 production, leading to the reduction of interaction between IL-13 and its receptor. The IL-13 receptor includes IL-4Rα and IL-13Rα1, and signaling is mediated predominantly through IL-4Rα (Hershey, 2003). Besides, PYC is also involved in the regulation of IL-13-induced JAK/STAT6 pathway. The increased IL-4R, JAK1 and STAT6 levels produced by IL-13-stimulated airway epithelial cells were reduced by the PYC, demonstrating that PYC regulates the inactivation of the JAK/STAT6 pathway through targeting the IL-13.
STAT6 is sensitive to IL-13 in the airway epithelial cells and this response is modulated by the PYC. PYC can obviously lower the IL-13-induced STAT6 and its phosphorylation level, and the STAT6 translocation to the nucleus drops as well. STAT6 is a cytoplasmic transcription factor regulated by its phosphorylation state and is activated in response to IL-13 (Darnell et al., 1994). After phosphorylation of a critical tyrosine residue, STAT6 dimerizes, translocates from the cytoplasm to the nucleus, and binds to its cognate DNA, leading to a specific transcriptional upregulation of the target genes (Hershey, 2003). IL-13, but not IL-4, also increased enteric contractility to acetylcholine by mechanisms that involved STAT6 and nerves (Zhao et al., 2003). Furthermore, findings of our study indicated that acetylcholine was upregulated by IL-13/JAK1/STAT6 signal pathway in OVA-induced asthma, and it may be one mechanism causing asthma progression. Besides, IL-13/JAK1/STAT6 signal pathway in airway smooth muscle cells may modulate contractility of smooth muscle tissue by acetylcholine-related cholinergic nervous system, resulting in airflow obstruction and airway hyperresponsiveness.
Cholinergic innervation also maintains resting bronchial tone and, to a significant degree, mediates acute bronchoconstriction, which is induced by acetylcholine binding to muscarinic receptors in the airways. Preganglionic parasympathetic nerve fibers release acetylcholine, which binds to nicotinic receptors on postganglionic neurons in the airways. Postganglionic parasympathetic neurons release acetylcholine and vasoactive intestinal peptide, which bind to muscarinic receptors on target tissues. Stimulation of the cholinergic nervous system induces secretion of mucus from goblet cells (Lundgren and Shelhamer, 1990).
Goblet cells are exocrine cells of the airway epithelium and are specialized in mucin exocytosis (Khan et al., 2001). The increase in the numbers of goblet cells can lead to excessive mucus production. The mucins hydrate to form a viscous gel acting as a major barrier between the airway contents and underlying mucosa, and thus cause an obstruction of the airways (Khan et al., 2001). Further support in favor of IL-13 being an important regulator in goblet cell hyperplasia is found in some studies, which show that IL-13-overexpressing mice develop intestinal goblet cell hyperplasia and increase mucin expression through the IL-13-dependent pathway (Steenwinckel et al., 2009). This process indicated the IL-13 activation in the development of goblet cell hyperplasia (Khan et al., 2001). Our data illustrate that PYC prevents the OVA-stimulated transcriptional activities of FOXA3, SPDEF, and Muc5ac, which were closely related to goblet cell metaplasia, and the JAK/STAT6 pathway is important in the regulation of goblet cell metaplasia. The IL-13-induced Muc5ac gene expression is mediated through a complex transcriptional network, in which the expression of the repressor, Forkhead Box protein A2, is reduced in response to STAT6 activation, whereas the SPDEF and FOXA3 expressions are increased and promote Muc5ac gene expression (Dabbagh et al., 1999; Atherton et al., 2003).
To summarize, our findings indicate that PYC prevents the emergence of OVA-induced airway injury and airway inflammation in mice by inhibiting the excessive secretion of IL-13 from Th2 cells, thereby causing inactivation of JAK/STAT6 pathway and obstructed release of acetylcholine to reduce goblet cell metaplasia. Given its preventing effect on asthma, PYC holds promise to extend multiple efficacies of therapy in airway inflammation and asthmatic patients.
Footnotes
Acknowledgments
The authors would like to thank the experimental animal center, Xi'an Jiaotong University Colleges of Medicine, who provided the experimental mice. All experiments were performed as per the approval of the Institutional Animal Care and Use Committee.
Disclosure Statement
No competing financial interests exist.