
Abstract
Highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) that emerged from classic PRRSV causes more severe damage to the swine industry. The earlier reports indicating inhibition of interferon-β (IFN-β) expression by PRRSV through total blockage of IFN-regulatory factor 3 (IRF3) nuclear translocation made us investigate the mechanism of IFN-β expression in HP-PRRSV infection. For this purpose, the IRF3 nuclear translocation in the control group [Poly (I:C)] and test group [Poly (I:C)+HP-PRRSV] was detected by immunofluorescence, and the results showed that IRF3 nuclear translocation in cells with PRRSV was weaker than cells without PRRSV, which was different from the previous study. In addition, the IFN-β mRNA and protein expression was observed to be inhibited by HP-PRRSV along with decreased IRF3 mRNA and total protein, and IRF3 nuclear translocation of test group was suppressed in MARC-145 and porcine alveolar macrophage cells in comparison with the control group. The quantity of phosphorylated IRF3 protein was also reduced after HP-PRRSV infection. However, CREB-binding protein (CBP) expression did not change between the control and test group. These results indicate that the inhibition of IFN-β expression is mainly due to the quantitative change in the amount of phosphorylated IRF3 in the cytoplasm, but not dependent on the complete blockage of IRF3 nuclear translocation or the restraining of CBP expression in the nucleus by HP-PRRSV.
Introduction
P
The first defense response of the host to RNA virus invasion is type I interferon (IFN) secretion (Sadler and Williams, 2008) and IFN-α was shown to suppress PRRSV replication (Le Bon et al., 2001), as well as recombinant swine IFN-β to protect PAMs and MARC-145 cells from PRRSV infection (Overend et al., 2007). However, the nonstructural protein 1 (NSP1) and the nucleocapsid of PRRSV were observed to inhibit IFN-β transcriptional production (Beura et al., 2010; Sagong and Lee, 2011), and PRRSV 3C-like protease prevented the production of IFN-β (Dong et al., 2015). Furthermore, IFN-β inhibition by PRRSV might be achieved through the retinoic acid-inducible gene 1 (RIG-I) signaling pathway (Loo et al., 2008; Li et al., 2015), especially through blocking the nuclear translocation of IFN-regulatory factor 3 (IRF3) (Beura et al., 2010). RIG-I could trigger IFN-β promoter stimulator 1 (IPS-1) (Jacobs and Coyne, 2013), and IPS-1 was responsible for activating transcription factors, such as tumor necrosis factor receptor-associated-factor 3 (TRAF3) and IRF3, and IRF3 further experienced phosphorylation (Morin et al., 2002; Chen et al., 2008), which led to structural changes and exposed its nuclear localization signal that subsequently entered the nucleus and interacted with the CREB-binding protein (CBP) to promote IFN-β transcription (Dragan et al., 2007; Edwards et al., 2007). During this process, IRF3 is an important transcription factor for IFN-β expression that plays an essential role for antiviral function (Lin et al., 2014).
Furthermore, several investigations have been done to find the effects of classic PRRSV infection on host immune responses confirming that the classic PRRSV is able to inhibit type I IFN expression, and two major mechanisms for the inhibition of type I IFN by PRRSV are raised out, one is the total blockage of IRF3 nuclear translocation (Luo et al., 2008) and the other is the interference with CBP but not IRF3 nuclear translocation (Kim et al., 2010). However, the exact mechanism for HP-PRRSV inhibiting type I IFN expression is not clear yet; therefore, we aim to determine the mechanism by detecting the IRF3 expression in MARC-145 and PAM cells.
Materials and Methods
Cells and virus
MARC-145 cells were cultured in Dulbecco's modified Eagle's medium, which was supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco), 100 U/mL penicillin, and 10 μg/mL streptomycin sulfate (Hyclone) and then was kept in an incubator at 37°C with 5% CO2. The HP-PRRSV strain WUH3 was conserved in the Key Laboratory. First, MARC-145 cells were divided into four groups, including cells treated with and without HP-PRRSV, and cells treated with polyinosinic:polycytidylic acid [Poly (I:C); Invitrogen] or both Poly (I:C) and HP-PRRSV, and then, type I IFN expression was detected. For the group treated with both Poly (I:C) and PRRSV, cells needed to be pretreated by adding with Poly (I:C) and absorbed with virus for 1 h at 37°C in the incubator (MOI = 0.5) for stable adhesion (Sun et al., 2012). After the absorption, culture media and cells were harvested for preparation of cellular extracts at indicated postinfection time, including 1, 12, 24, 36, 48 hpi, respectively, and to keep the number of collected cells at different time point proximities by cell counting.
PAMs were harvested by lung lavage from 4- to 6-week-old crossbred (Duroc×Large White×Landrace) male- and female-specific pathogen-free pigs, and all pigs were identified as HP-PRRSV negative by enzyme-linked immunosorbent assay (ELISA). Bronchoalveolar lavage was performed as described previously (Chiou et al., 2000). PAMs obtained from lavage were suspended in the RPMI-1640 medium (Gibco Laboratories) with 10% heat-inactivated FBS (Gibco Laboratories), 100 U/mL penicillin, and 10 μg/mL streptomycin sulfate (Hyclone). The PAMs were treated by the same methods as MARC-145 cells as described previously.
RNA extraction and quantitative real-time RT-PCR
Total RNA was extracted from different infected time points (1, 12, 24, 36, 48 hpi) using an E.Z.N.A. total RNA kit (Omega Bio-tech) by following the company's instructions.
The cDNA was generated by reverse transcription of 500 ng total RNA by applying the RevertAid First Strand cDNA Synthesis Kit (Fermentas). Quantitative real-time RT-PCR was performed using SYBR Green Real Time PCR Master Mix (Bio-Rad) on the Light Cycler 480 PCR system (Roche Applied Science). Seven pairs of gene-specific primers were used during this study (Table 1). The cycling conditions of program were 10 min of polymerase activation at 95°C, followed by 40 cycles at 94°C for 15s, 60°C for 60s and 60°C for 60s. Melting curves were generated at each run to show the process of PCR. Individual transcriptions of each sample were detected by three repetitions and normalized with the mRNA level of house-keeping gene β-actin as a control gene. Fold changes in gene expression were calculated using the comparative cycle threshold (Ct) method (Kim et al., 2010).
CBP, CREB-binding protein; IFN-β, interferon-β; IRF3, IFN-regulatory factor 3.
Cellular extracts and Western blot assay
Both MARC-145 and PAM cells were cultured in six-well plates and pretreated with Poly (I:C) (300 ng/mL; Invitrogen) and HP-PRRSV as described before, and then, total proteins were extracted from both cells in different infected time points (1, 12, 24, 36, 48 hpi). Washed cells were lysed in RIPA buffer added with 1% phenylmethylsulfonyl fluoride (PMSF) as the protease inhibitor to get proteins, and the protein concentrations were determined to standardize the concentrations before Western blot analysis. The process of phosphorylated protein extraction was similar as the total protein extraction, except that the protease inhibitor was not PMSF, but Ser-specific phosphorylation inhibitor.
Cytoplasmic and nuclear proteins were extracted from MARC-145 and PAM cells at different infected time points (1, 12, 24, 36 hpi) using the Qproteome Cell Compartment Kit (Qiagen, Inc.) referring to the manufacturer's protocol. The protein concentration was also measured and standardized before detecting the target protein expression.
For the preparation of Western blot, the extractions of the cells were boiled in sodium dodecyl sulfate (SDS), a protein sample buffer purchased from Sangon Biological Engineering Technology Company, and then, the protein expression of different protein fractions by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was analyzed. The individual proteins were transferred onto a PVDF membrane (Bio-Rad), which were blocked with 5% (w/v) dried skimmed milk dissolved by Tris-buffered saline containing Tween 20. Specific antibodies were used in this method; anti-IRF3 was used to detect total IRF3 (4302s, rabbit mAb D83B9; Cell Signaling Technology), anti-phosphorylated IRF3 (pho-IRF3) was used to detect the phosphorylated site (Ser396) (4947s, rabbit mAb 4D4G; Cell Signaling Technology), anti-Histone H3 (0349R; Abcam), anti-β-actin (ab8229; Abcam), and anti-glyceraldehyde 3-phosphate dehydrogenase (Sigma) were used to detect reference proteins, and anti-NSP2 for detecting HP-PRRSV was prepared and conserved in the Key Laboratory. The Western blot results were analyzed by Quantity One software.
IFN-β ELISA
MARC-145 cells were cultured in six-well plates and subsequently pretreated with Poly (I:C) and HP-PRRSV according to the description as depicted before, and then, secreted IFN-β of the culture supernatant at different infected time points (1, 12, 24, 36, 48 hpi) was measured by the human IFN-β-specific ELISA kit (PBL Assay Science) according to the manufacturer's protocol.-
Immunofluorescence
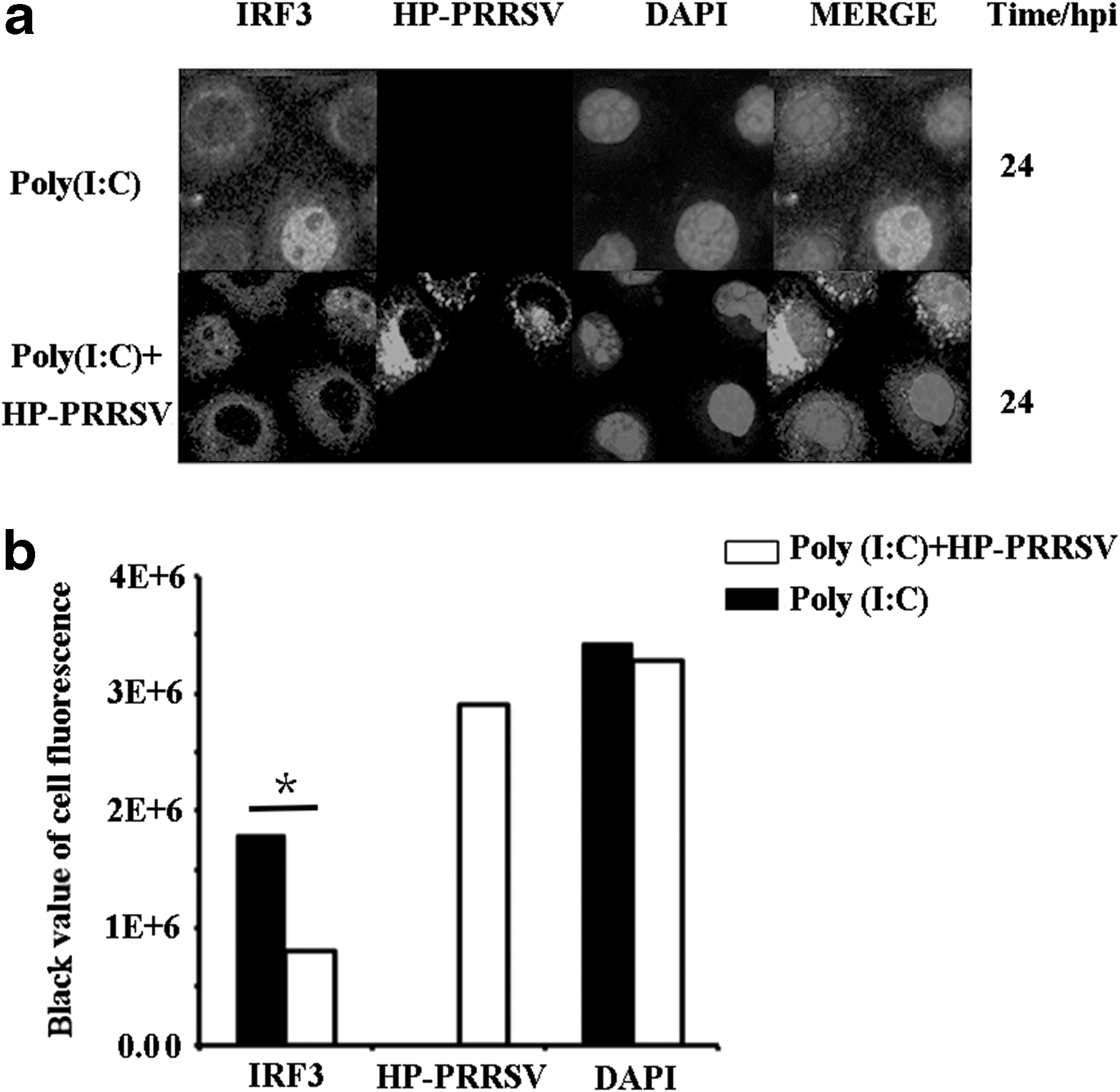
Cells were grown on coverslips and then pretreated with Poly (I:C) and PRRSV as described before. Cells of different time points were washed once with phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde for 30 min, washed thrice with PBS, permeabilized with 0.5% Triton X-100 for 15 min, washed thrice with PBS, and blocked for 1 h with 5% bovine serum albumin. Then, the samples were incubated with anti-IRF3 and anti-NSP2 for 12 h at 4°C, followed by a staining step with secondary anti-rabbit antibody fluorescein isothiocyanate (1:100) and secondary anti-mouse Cy3 antibody (1:100; Boster) for 1 h, washed once, a nuclear counter stain with DAPI (4′,6-diamidino-2-phenylindol; Sigma) performed for 5 min at room temperature, and finally detected by fluorescence under a confocal microscope (Axioskop II; Zeiss). The quantity of cell fluorescence was detected by ImageJ software (McCloy et al., 2014).
Statistical analysis
Data were indicated as average value ± standard error of the mean from three independent tests. Statistical analysis was used to determine the significant level of different time points by t-test, and p-values less than 0.05 and 0.01 were considered as statistically significant and highly statistically significant, respectively.
Results
HP-PRRSV could not block IRF3 nuclear translocation in MARC-145 cells
The IRF3 nuclear translocation in MARC-145 cells treated with Poly (I:C) and with both Poly (I:C) and HP-PRRSV was detected by immunofluorescence, and the cell fluorescence was tested by ImageJ software (McCloy et al., 2014). The nuclear translocation of IRF3 could be detected in both groups at 24 hpi (Fig. 1a), and the cells with HP-PRRSV showed a weaker IRF3 trend than cells without HP-PRRSV (Fig. 1b), which is different from the previous study of total blockage of IRF3 nuclear translocation. However, the accurate quantity of each group was not clear; thus, the IRF3 expression tendency needs deep investigation as well as the impact on the IFN-β expression by HP-PRRSV.
HP-PRRSV inhibited IFN-β expression in MARC-145 cells
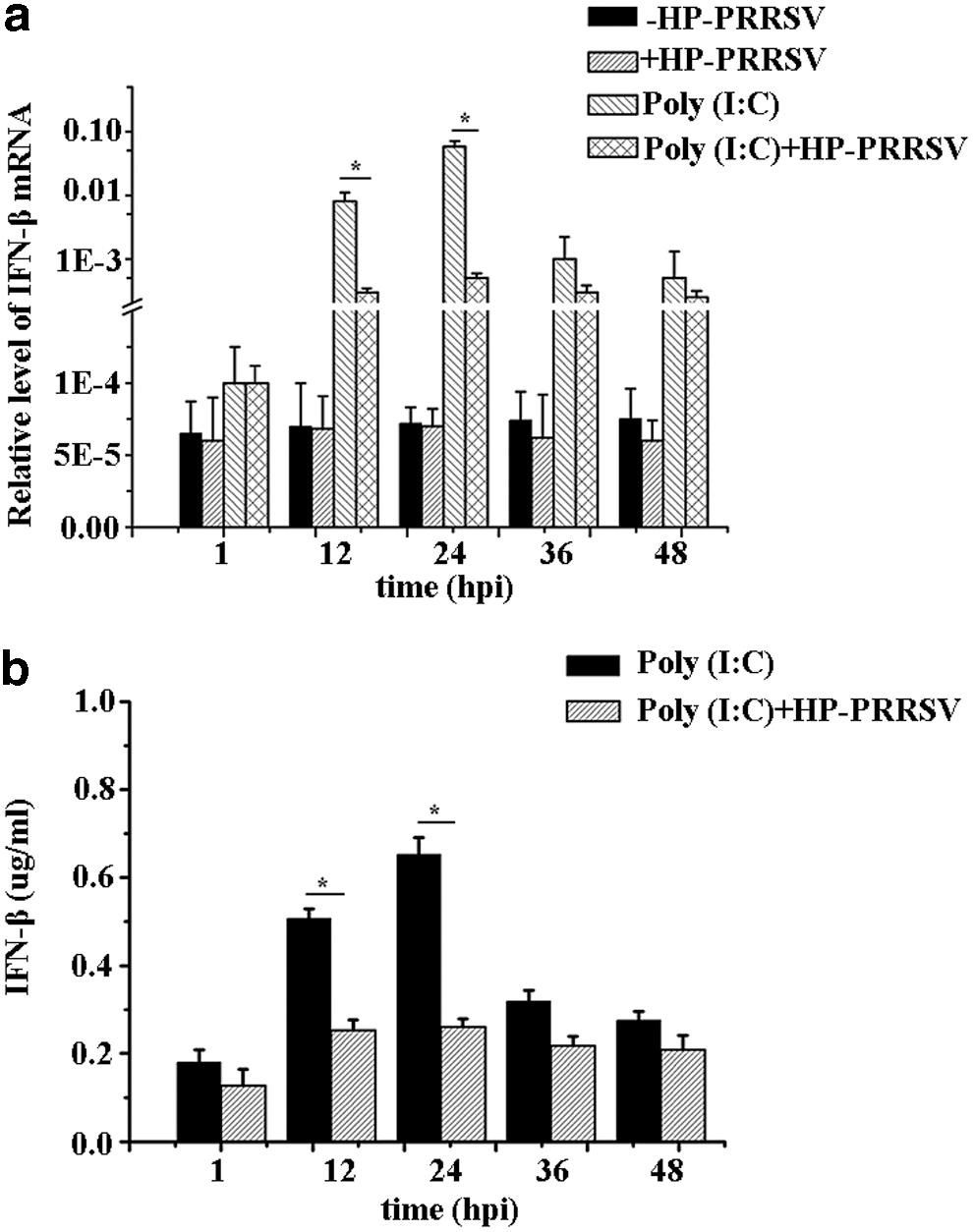
IFN-β mRNA expression in MARC-145 cells with different treatments [HP-PRRSV, +HP-PRRSV, Poly (I:C), Poly (I:C)+HP-PRRSV] was detected at five different time points (1, 12, 24, 36, 48 hpi). IFN-β mRNA expression of HP-PRRSV showed a decreased trend when compared with the uninfected group in MARC-145 cells, however, both of them were extremely low and difficult for further study. It was observed that Poly (I:C) (a common type I IFN stimulator) could increase the expression of IFN-β mRNA, whereas the stimulation could be significantly inhibited by HP-PRRSV infection (Fig. 2a). In addition, the expression tendencies of IFN-β in the four groups were similar. Therefore, cells treated with Poly (I:C) were considered as the control group, and cells treated with both Poly (I:C) and HP-PRRSV were taken as the test group to investigate the mechanism of IFN-β inhibition by HP-PRRSV infection.
IFN-β mRNA and protein expression was detected in MARC-145 cells.
Then, the IFN-β protein in MARC-145 cells was detected. Supernatant from the MARC-145 test group and control group cells was collected to detect IFN-β protein expression at five time points (1, 12, 24, 36, 48 hpi). The IFN-β protein expression of test group was lower than that of the control group, and the expression tendency was similar with IFN-β mRNA in MARC-145 cells (Fig. 2b), indicating that IFN-β protein expression could also be inhibited by HP-PRRSV.
HP-PRRSV inhibited IFN-β mRNA and protein expression in PAM cells
PAM cells are the natural target cells for HP-PRRSV infection. IFN-β mRNA and protein expression was detected in PAM cells. The results showed that IFN-β mRNA expression decreased in the test group when compared with the control group at five different time points (1, 12, 24, 36, 48 hpi), and its tendency changed at different time points, where 24 hpi is observed to be the most significant (Fig. 3a). Furthermore, the supernatant of PAM cells was collected to analyze the IFN-β protein expression that shows a decrease in IFN-β protein expression in the test group compared to the control group (Fig. 3b), which was consistent with the IFN-β mRNA expression in PAM cells.
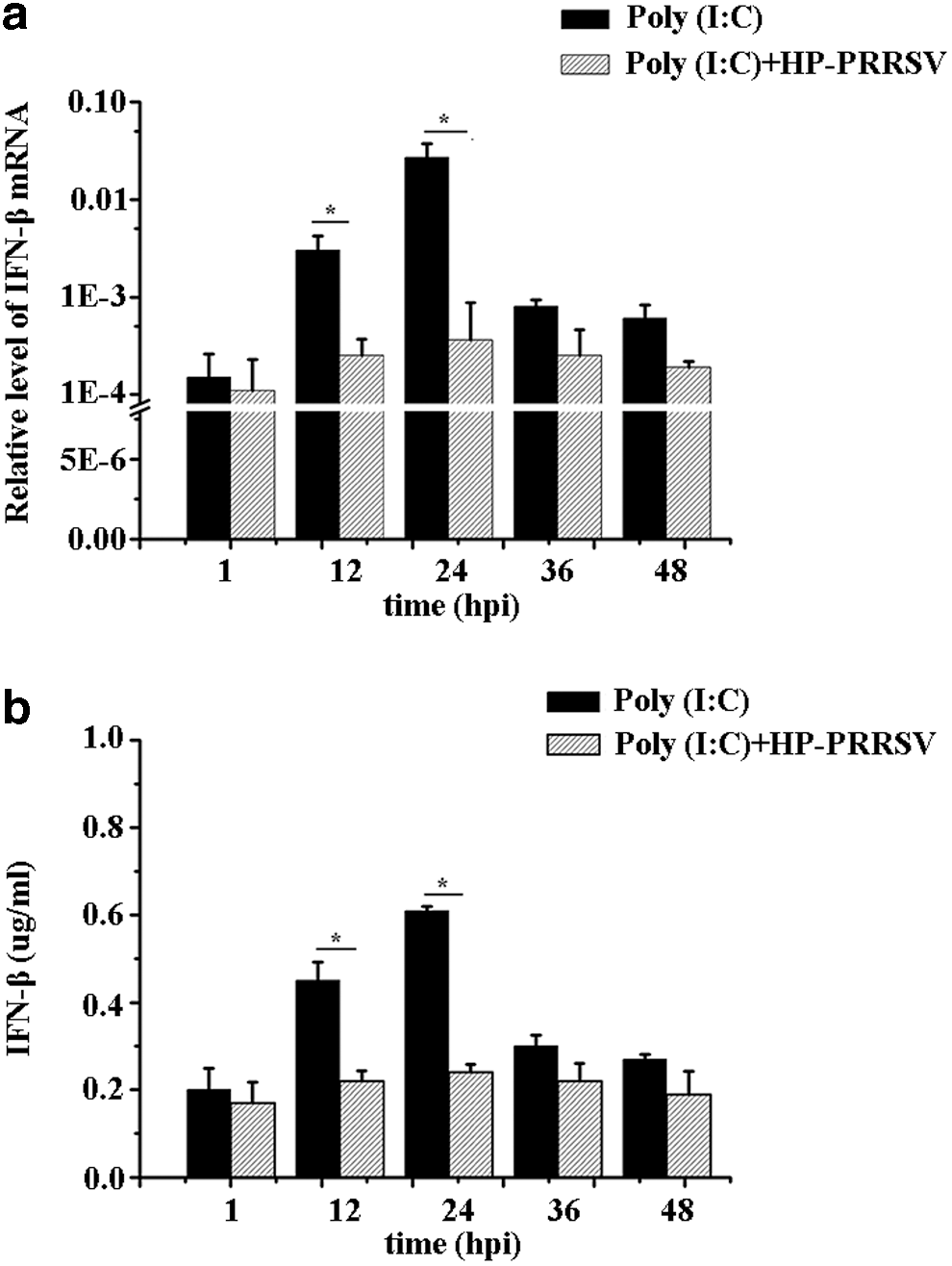
IFN-β mRNA and protein expression was detected in PAM cells.
HP-PRRSV inhibited IRF3 mRNA and total protein in MARC-145 and PAM cells
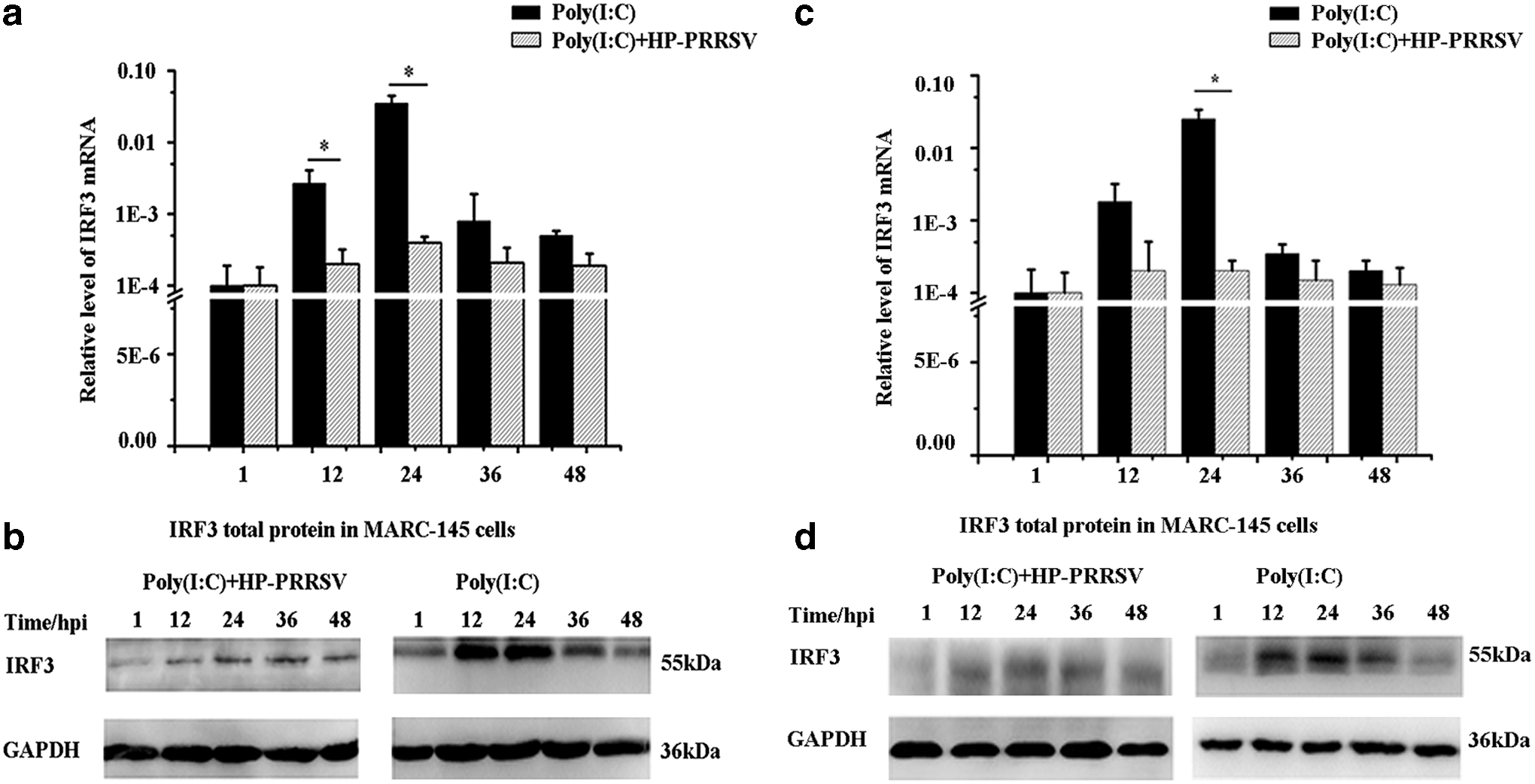
IRF3 is important for regulating IFN-β expression; therefore, IRF3 mRNA and total protein of test group and control group in MARC-145 cells were detected at five different time points. IRF3 mRNA of test group was lower than the control group (Fig. 4a), which was the same with IRF3 total protein expression (Fig. 4b), and the tendency of IRF3 total protein was similar with IRF3 mRNA expression in MARC-145 cells.
IRF3 mRNA and protein expression was detected in MARC-145 and PAM cells.
Furthermore, IRF3 mRNA and total protein of test group and control group in PAM cells were detected at five different time points, and IRF3 mRNA and total protein of test group were lower than control group in PAM cells, respectively (Fig. 4c, d). The results showed that the expression of IRF3 mRNA and total protein was inhibited by HP-PRRSV in both MARC-145 and PAM cells.
HP-PRRSV inhibited IRF3 nuclear translocation in MARC-145 and PAM cells
The stimulation of IRF3 on IFN-β expression is related with the quantity of IRF3 entering the nucleus; thus, the nuclear IRF3 protein of test group and control group was detected in MARC-145 cells at four different time points. The results indicated that both cytoplasmic and nuclear IRF3 proteins of the test group significantly reduced when compared with the control group (Fig. 5a), especially for the nuclear IRF3 protein. Meanwhile, the quantity of protein was calculated by Quantity One software (Fig. 6a, b), which demonstrated that HP-PRRSV could significantly decrease the quantity of IRF3 entering the nucleus.

Cytoplasmic and nuclear IRF3 protein was detected in MARC-145 and PAM cells.
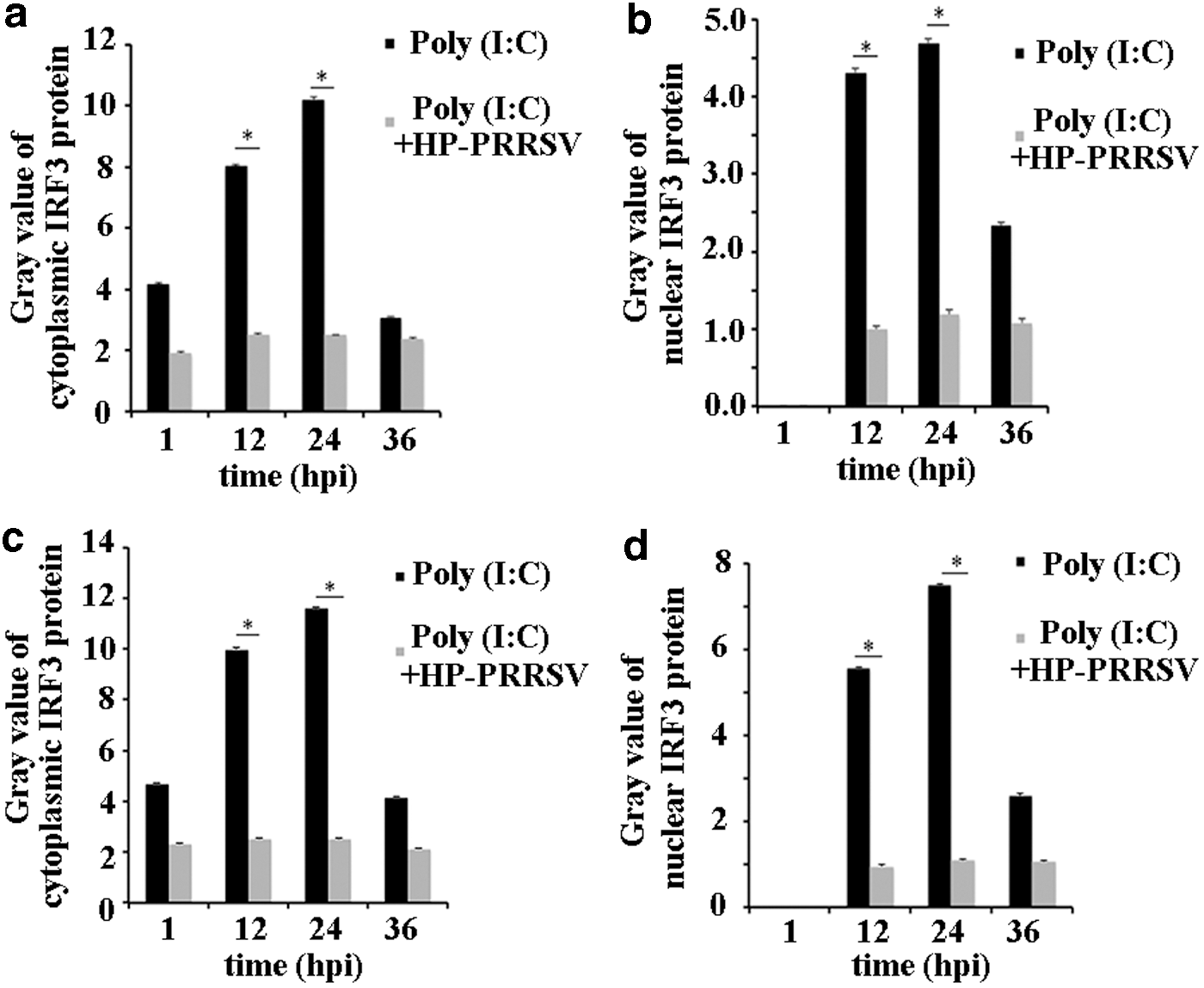
Cytoplasmic and nuclear IRF3 protein was calculated using Quantity One software and showed as gray value. Cytoplasmic
Furthermore, the cytoplasmic and nuclear IRF3 proteins of test group and control group were detected in PAM cells at five different time points. The cytoplasmic IRF3 protein decreased in the test group compared to the control group, which was the same as the nuclear IRF3 protein (Fig. 5b). In addition, the Quantity One software results indicated that HP-PRRSV was able to reduce the quantity of IRF3 entering the nucleus in PAM cells (Fig. 6c, d).
HP-PRRSV inhibited IRF3 phosphorylation in MARC-145 and PAM cells
IRF3 phosphorylation is a critical step for IRF3 stimulation to enter the nucleus; thus, an antibody of IRF3 (Ser396) was used to detect the phosphorylation of test group and control group in MARC-145 cells. It was found that phosphorylated IRF3 protein of test group was lower than that of control group (Fig. 7a).
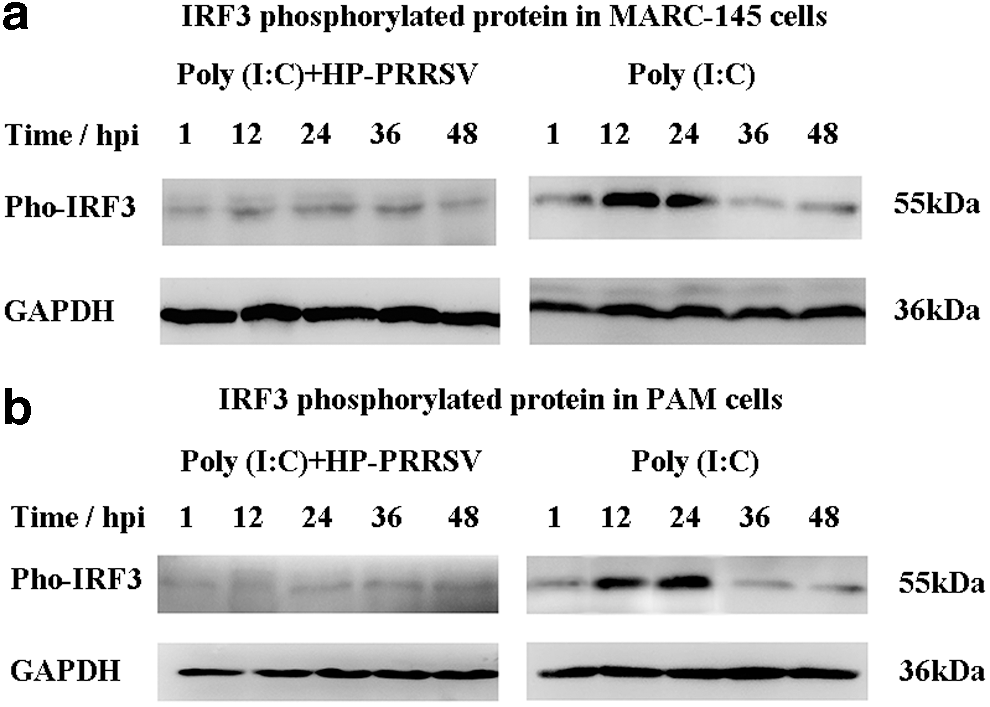
Phosphorylated IRF3 protein was detected in MARC-145
Furthermore, phosphorylated IRF3 protein of test group and control group was detected at five different time points. Phosphorylated IRF3 protein of test group was significantly lower compared with the control group (Fig. 7b), indicating that IRF3 phosphorylation was decreased by HP-PRRSV infection.
HP-PRRSV did not affect CBP expression in MARC-145 and PAM cells
The combination of nuclear IRF3 and CBP is necessary for stimulating IFN-β expression; thus, the CBP expression was detected. However, different from IRF3 expression, the CBP mRNA did not change significantly between the control and test group in both MARC-145 and PAM cells (Fig. 8a, b). The results indicated that HP-PRRSV might have no influence for the CBP expression.

CBP mRNA was detected in MARC-145
Discussion
HP-PRRSV is derived from classic PRRSV and has a higher pathogenicity than classic PRRSV. The type I interferon acts as a barrier against viral infections and has a predominant role in the innate immune system. Although many investigations have been performed to discover the effects of classic PRRSV infection on host immune response that reports PRRSV ability to inhibit IFN-β expression through blocking IRF3 nuclear translocation, the results in this study showed a different mechanism. First, IRF3 nuclear translocation results detected by immunofluorescence indicated that HP-PRRSV might not totally block IRF3 nuclear translocation although weaker IRF3 expression of cells with HP-PRRSV than without virus infection, and IFN-β mRNA and total protein expression was inhibited after HP-PRRSV infection. Thus, the mechanism of IFN-β inhibition was deeply investigated in MARC-145 cells first and further confirmed in PAM cells. IRF3 mRNA and total protein results of MARC-145 and PAM cells demonstrated that they were also inhibited by HP-PRRSV infection, and IRF3 nuclear translocation of test group detected by Western blot decreased when compared with control group. In addition, the phosphorylation was inhibited by HP-PRRSV infection. The results indicated that during the HP-PRRSV infection, the amount of phosphorylated IRF3 was decreased, leading to the inhibition of IRF3 nuclear translocation and the next step of interaction with CBP (Fig. 9). Thus, the mechanism of IFN-β inhibition by HP-PRRSV might be different from classic PRRSV, which is mainly through quantitative change of phosphorylated IRF3, but neither total block of IRF3 nuclear translocation nor interference with CBP.
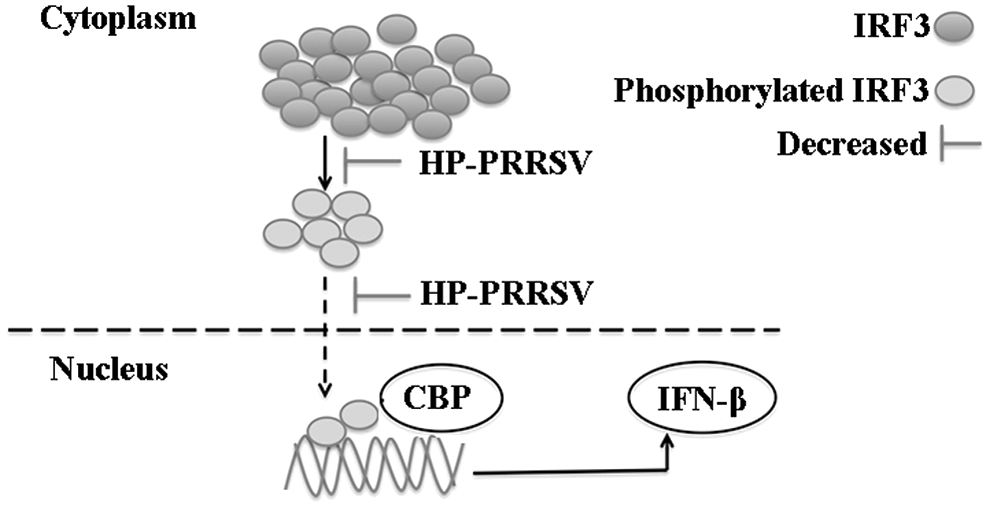
The inhibition of IFN-β attributes to the quantitative change of IRF3 phosphorylation by HP-PRRSV. HP-PRRSV reduces the phosphorylated IRF3, which weakens the next step of IRF3 nuclear translocation, and decreases the quantity of IRF3 interacting with CBP to inhibit IFN-β expression. HP-PRRSV, highly pathogenic porcine reproductive and respiratory syndrome virus.
Furthermore, Ser396 is an important site for IRF3 phosphorylation and interaction with CBP (McWhirter et al., 2004; Panne et al., 2007). The IRF3 phosphorylation related with this site was analyzed using I-Tasser software (
In conclusion, the comparison of IFN-β and IRF3 expression in MARC-145 and PAM cells confirmed that HP-PRRSV infection could reduce but not totally block the phosphorylated IRF3 protein level, which would further decrease the number of IRF3 entering the nucleus, and finally inhibit IFN-β expression. Therefore, the quantitative change of IRF3 phosphorylation is responsible for the inhibition of IFN-β by HP-PRRSV.
Footnotes
Acknowledgment
Financial assistance from the NSFC (Natural Science Foundation of China) (31272427, 31572367) is greatly appreciated.
Disclosure Statement
No competing financial interests exist.