
Abstract
Introduction
B
Hematopoietic PBX-interaction protein (HPIP) is originally identified as a transcription factor PBX1-interacting protein in the hematopoietic system (Abramovich et al., 2002). Moreover, it is found to bind to microtubule (Manavathi et al., 2006). Accumulative evidence from studies on breast cancer, liver cancer, colorectal cancer, and thyroid cancer supports an important role of HPIP in tumor initiation and progression (Wang et al., 2008, 2015; Xu et al., 2013a, 2013b; Feng et al., 2015). In breast cancer, previous studies showed that HPIP enhanced breast cancer cell proliferation in both cell line (MCF-7) and xenograft models (Wang et al., 2008). The possible mechanism involves estrogen signaling and activation of AKT and MAPK, which are integrated into the HPIP-microtubule complex having estrogen receptor (ER), Src kinase, and the p85 subunit of phosphatidylinositol 3-kinase recruited (Manavathi et al., 2006).
In terms of the crucial role of HPIP as an MBP coordinating multiple signaling pathways, we aim to determine the impact of HPIP on sensitivity of breast cancer cells to paclitaxel. The potential association between HPIP level and paclitaxel sensitivity in breast cancer may guide patient selection for paclitaxel-based regimens in the future.
Materials and Methods
Cell line and culture condition
Breast cancer cell lines, MCF-7 (ER-positive) and MDA-MB-231 (triple-negative), were obtained from American Type Culture Collection and maintained in DMEM supplemented with 10% fetal bovine serum (GIBCO, Life Technologies, Beijing, China) in a humidified incubator at 37°C in the presence of 5% CO2. The transfection of MCF-7 and MDA-MB-231 cells was done using Lipofectamine 3000 reagent (Invitrogen, Life Technologies, Beijing, China) according to the manufacturer's instructions.
Plasmid
The DNA fragment encoding HPIP with an N-terminal Flag tag was subcloned into mammalian cell expression vector pcDNA3.1. Breast cancer cells cultured in a 60-mm dish were transfected with 5 μg of pcDNA3.1-Flag-HPIP or the empty vector as control. To express HPIP in the prokaryotic system, HPIP cDNA was inserted into pGEX-4T-1 vector using BamHI/NotI sites, which fused HPIP to the C-terminal of glutathione S-transferase (GST).
Purification of recombinant GST-HPIP
A single colony on the master plate of Escherichia coli BL21DE3 transformed with pGEX-4T-GST-HPIP was picked for amplification in LB medium. The bacteria culture was induced by 1 mM IPTG (isopropyl-β-D-thiogalactoside) at 37°C for 3 h to transiently express the fusion protein, GST-HPIP. Cells were harvested after centrifugation and resuspended in 1 × phosphate buffer saline (PBS) for sonication to prepare cell lysate. Affinity purification was conducted using the GST fusion protein purification kit provided by GenScript (Nanjing, China). The purity of GST-HPIP in eluted fractions was monitored using SDS-PAGE.
Western blot analysis
Cells were harvested at 48 h after transfection and washed twice before the lysis in buffer containing 25 mM Tris-HCl, pH 7.4, 135 mM NaCl, 0.5% NP-40, and 0.1% SDS supplemented with proteinase inhibitor cocktail (Sigma-Aldrich, MO). Protein samples (20 μg per lane) were run on 10% SDS-PAGE for separation and then electrotransferred onto a nitrocellulose membrane. The blots were incubated in TBS buffer (20 mM Tris-HCl, pH 7.4, 150 mM NaCl) with 5% nonfat milk and 0.02% Tween 20 at room temperature for 1 h, then washed for primary antibody incubation overnight at 4°C. HRP-conjugated secondary antibody (Beyotime Biotechnology, Beijing, China) incubation was conducted at room temperature for 1 h. The blots were developed using Western Lightning-Enhanced Chemiluminescence Substrate (Perkin Elmer) and imaged using X-ray films.
MTT assay and IC50 value
On day 1, cells in a 6-well plate were transfected with 2 μg DNA per well of either pcDNA3.1-Flag-HPIP or the empty vector. Twenty-four hours later, cells were transferred into a 96-well plate at a density of 104 cells per well. On day 3, cells were treated with the indicated amount of paclitaxel or vinblastine. After incubation for 24 h, 20 μL MTT (5 mg/mL; Sigma-Aldrich, St. Louis, MO) was added to each well and the plate was kept for another 4 h in a cell incubator. Upon removal of the medium, 100 μL dimethyl sulfoxide was added into each well. The absorbance of the solution at 490 nm was measured. The IC50 value of paclitaxel or vinblastine was estimated based on the curve of cell growth rate versus drug concentration in log scale.
Matrigel transwell assay
Ten microliters of fibronectin (0.5 mg/mL; Sigma-Aldrich) was evenly distributed onto the bottom surface of the membrane in a transwell chamber (Corning) and left to air dry in a biosafety cabinet. The upper chamber was paved with 50 μL Matrigel (BD Biosciences) and incubated for 40 min at 37°C. Cells at 48 h after transfection were transferred to the upper chambers at 105 cells/well and cultured in Opti-MEM (GIBCO, Life Technologies, Beijing, China). The lower chamber was added with ECM (GIBCO, Life Technologies). Sixteen hours later, nonmigrated cells were wiped using cotton swabs, and migrated cells were fixed with 100% methanol for 30 min and stained using crystal violet for 15 min. Cell images were captured under a microscope with a camera.
Microtubule assembly/disassembly assay
The dynamics of tubulin polymerization was recorded by measuring the optical absorbance at 350 nm of the reaction in time course, which contained 70 μM tubulin (>99% purity, Cytoskeleton, Inc., Denver, CO) and 200 nM paclitaxel in PEMG buffer (50 mM PIPEs, 1 mM EGTA, 1 mM MgCl2, and 4% glycerol) plus 1 mM GTP with or without the purified recombinant HPIP. In microtubule disassembly assay, the paclitaxel-stabilized microtubules were prepared first in the presence or absence of recombinant HPIP. Depolymerization was initiated by adding PEMG buffer for dilution and then further triggered by temperature shift from 37°C to 4°C.
Immunofluorescence microscopy
For immunofluorescence study, cells in the wells that had a transfection efficiency of >90% for pCDNA3.1-Flag-HPIP were chosen for processing. Cells grown on coverslips were fixed with 1% formaldehyde in PBS for 15 min at room temperature, followed by permeabilization in 0.2% Triton X-100. For nonspecific antigen blocking, coverslips were incubated with 2% goat serum in PBS in a humidified chamber for 1 h at room temperature. Rabbit polyclonal antialpha tubulin antibody and mouse monoclonal antiacetylated alpha tubulin antibody were diluted at 1:200 and 1:100, respectively, for primary antibody incubation overnight at 4°C. Secondary antibody incubation was performed at room temperature for 1 h using AlexFluor-594-labeled goat anti-mouse IgG and AlexFluor-488-labeled goat anti-rabbit IgG (Jackson ImmunoResearch, PA) at 1:500 dilution. The coverslips were washed three times and then mounted for imaging under a Leica TCS SP5 laser scanning confocal microscope.
Statistical analysis
Student's t-test was used to examine the difference in mean value between two groups. p < 0.05 was considered to be statistically significant.
Results
Overexpression of HPIP promoted the sensitivity of breast cancer cells to paclitaxel
To observe the impact of HPIP on cytotoxicity of paclitaxel to breast cancer cells, we transfected MCF-7 and MDA-MB-231 cells with the recombinant plasmid pcDNA3.1-Flag-HPIP or the pcDNA3.1 backbone. Western blot analysis confirmed the ectopic expression of HPIP in cells transfected with pcDNA3.1-Flag-HPIP, but not in cells mock transfected or transfected with pcDNA3.1 backbone (Fig. 1A, C). Next, breast cancer cells were treated with series of paclitaxel dosage. The cell growth rate under each dosage was evaluated by MTT assay and plotted against the log concentration of paclitaxel for IC50 estimation. Results showed that in both MCF-7 and MDA-MB-231 cell lines, cells transfected with pcDNA3.1-Flag-HPIP had a significant lower IC50 value of paclitaxel than did the cells mock transfected or transfected with pcDNA3.1 backbone, almost a 50% reduction in IC50 value of paclitaxel compared with control (Fig. 1B, D). Interestingly, overexpression of HPIP showed no impact on cytotoxicity of the microtubule catastrophe agent, vinblastine, to either MCF-7 or MDA-MB-231 cells.
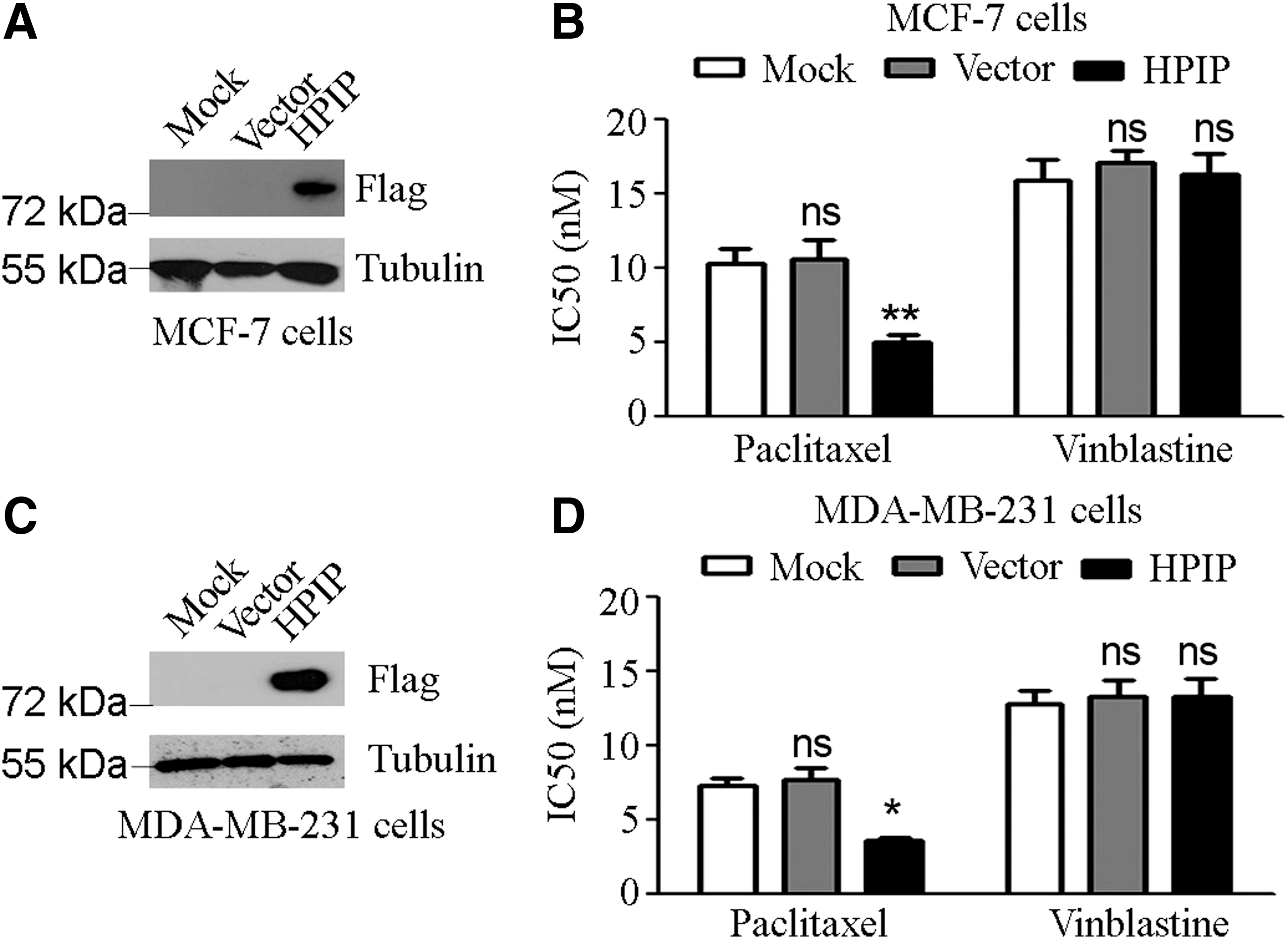
Overexpression of HPIP promoted breast cancer cell sensitivity to paclitaxel.
HPIP enhanced the inhibition of breast cancer cell migration by paclitaxel
Matrigel transwell assay was conducted to observe the effects of HPIP overexpression on migration of breast cancer cells treated with paclitaxel. Counting of the migrated cells revealed a trend that paclitaxel appeared to inhibit the migration of both MCF-7 and MDA-MB-231 cells transfected with pcDNA3.1 vector compared with the vehicle control, but the inhibition was not statistically appreciable (Fig. 2). To be noticed, paclitaxel treatment resulted in a significant inhibition of cell migration to breast cancer cells transfected with pcDNA3.1-Flag-HPIP compared with the vehicle control (Fig. 2).
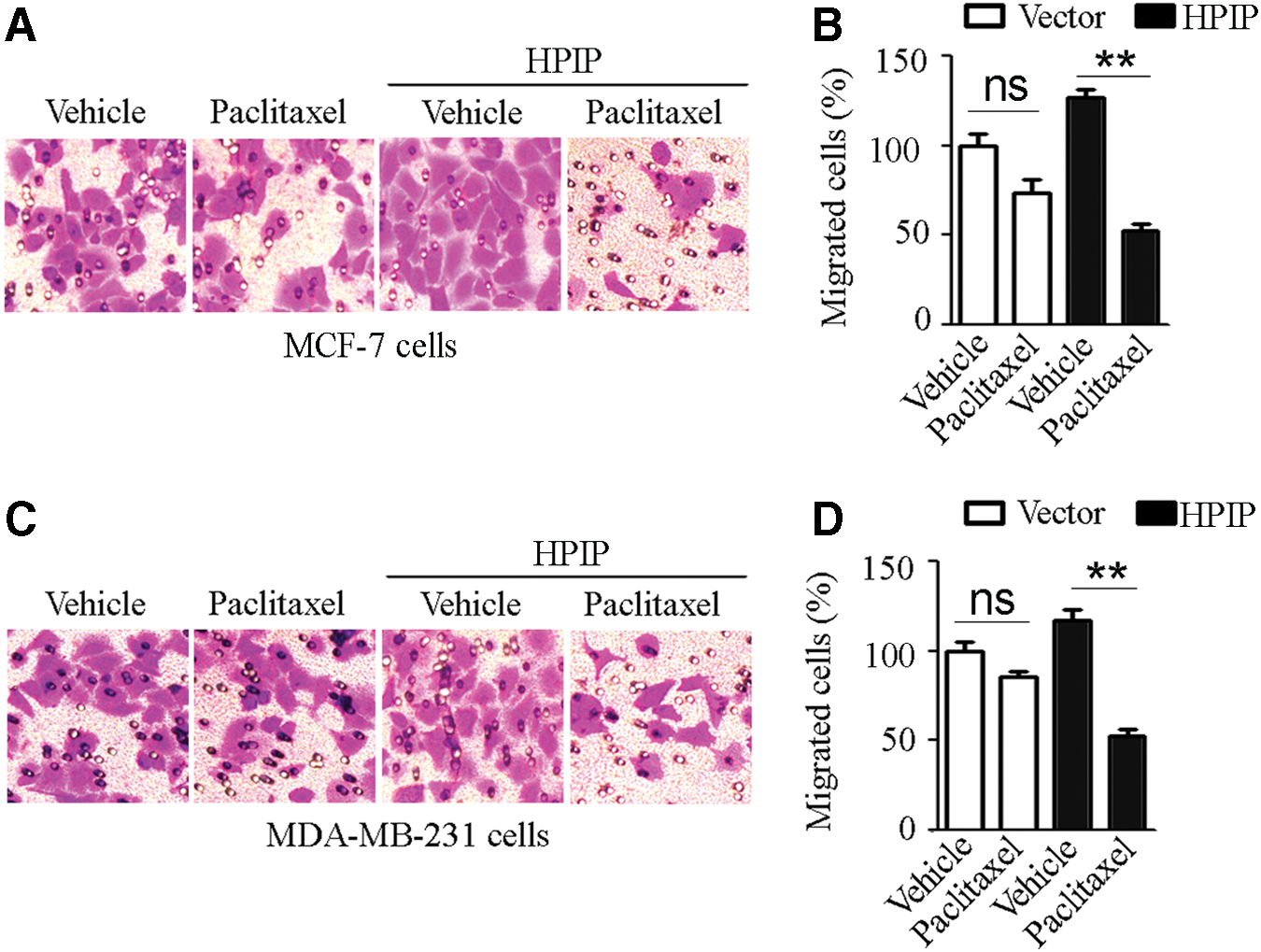
HPIP enhanced the inhibitory effect of paclitaxel on breast cancer cell migration.
HPIP enhanced microtubule assembly and stability in vitro
Coomassie blue staining after SDS-PAGE showed that the prepared GST-HPIP was in good purity for in vitro assays (Fig. 3A). The dynamics of tubulin polymerization in the presence or absence of HPIP were recorded by measuring the absorbance at 350 nm of the reaction. The absorbance of time course revealed that tubulin polymerization at initial stage of the reaction was faster in the presence of HPIP than in the absence of HPIP (Fig. 3B). When the reaction reached steady state, a greater absorbance at 350 nm was observed in the presence of HPIP than in the absence of HPIP (Fig. 3B). Moreover, HPIP appeared to promote tubulin polymerization in a dose-dependent way (Fig. 3B). In microtubule disassembly assay, the prepared microtubule showed higher dynamic stability in the presence of HPIP than in the absence of HPIP, as evidenced by a slower declining of absorbance at 350 nm in the reaction with HPIP (Fig. 3C). Seemingly, HPIP enhanced the microtubule stability dose-dependently in the disassembly assay (Fig. 3C).
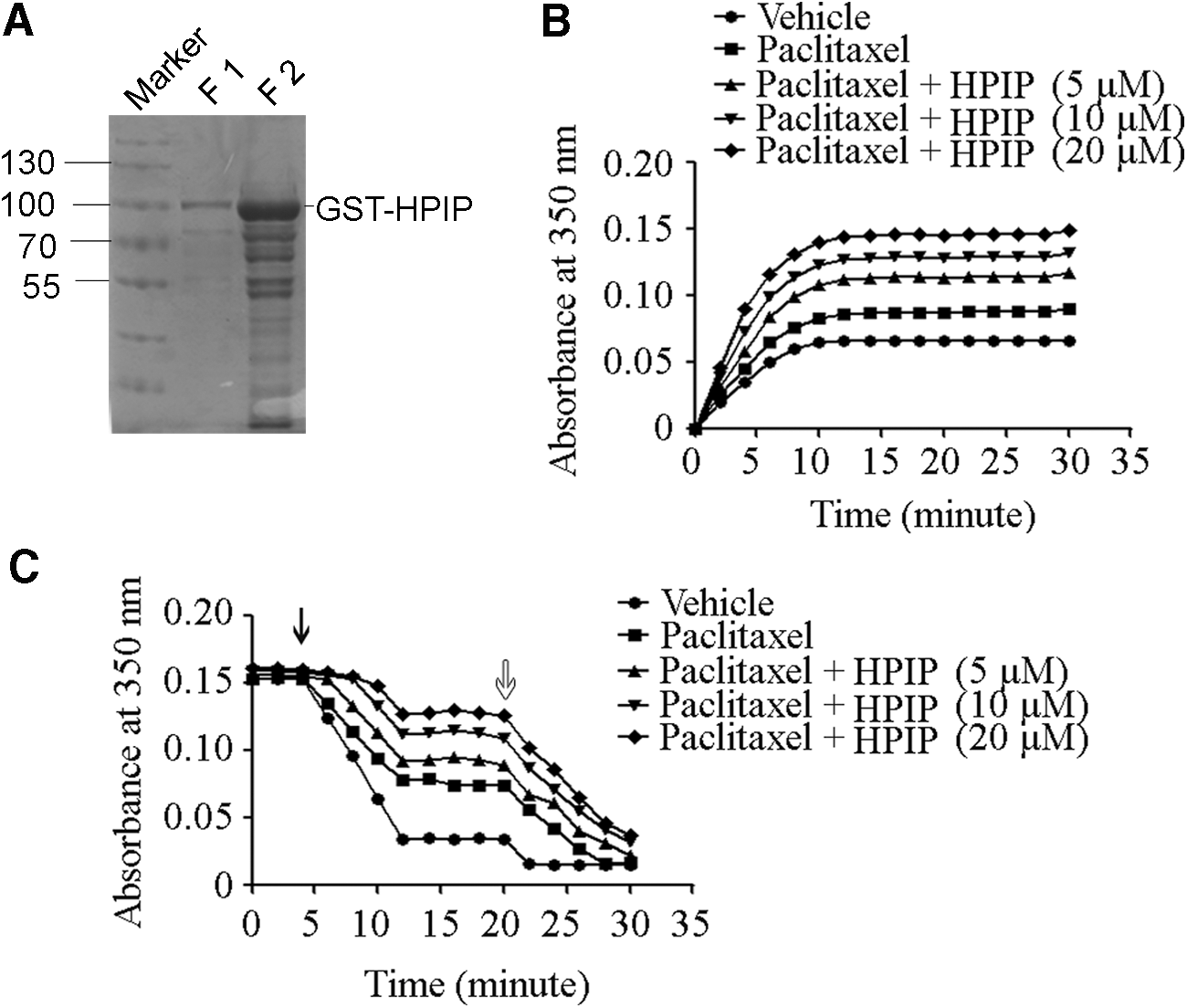
HPIP enhanced microtubule assembly and stability in vitro.
HPIP suppressed the expression of HDAC6, leading to increased acetylation of tubulin
Because paclitaxel is a microtubule-stabilizing agent, we inferred that HPIP might act through a microtubule-dependent mechanism. Of interest, Western blot analysis showed that overexpression of HPIP in MCF-7 cells decreased the level of HDAC6, which is an important regulator of microtubule network, and that the acetylated α-tubulin significantly increased (Fig. 4A, C). Using specific antibodies to follow α-tubulin and acetylated α-tubulin under a fluorescence microscope, we observed the enhanced acetylation of α-tubulin in MCF-7 cells transfected with pcDNA3.1-Flag-HPIP compared with MCF-7 cells transfected with pcDNA3.1 vector (Fig. 4B).
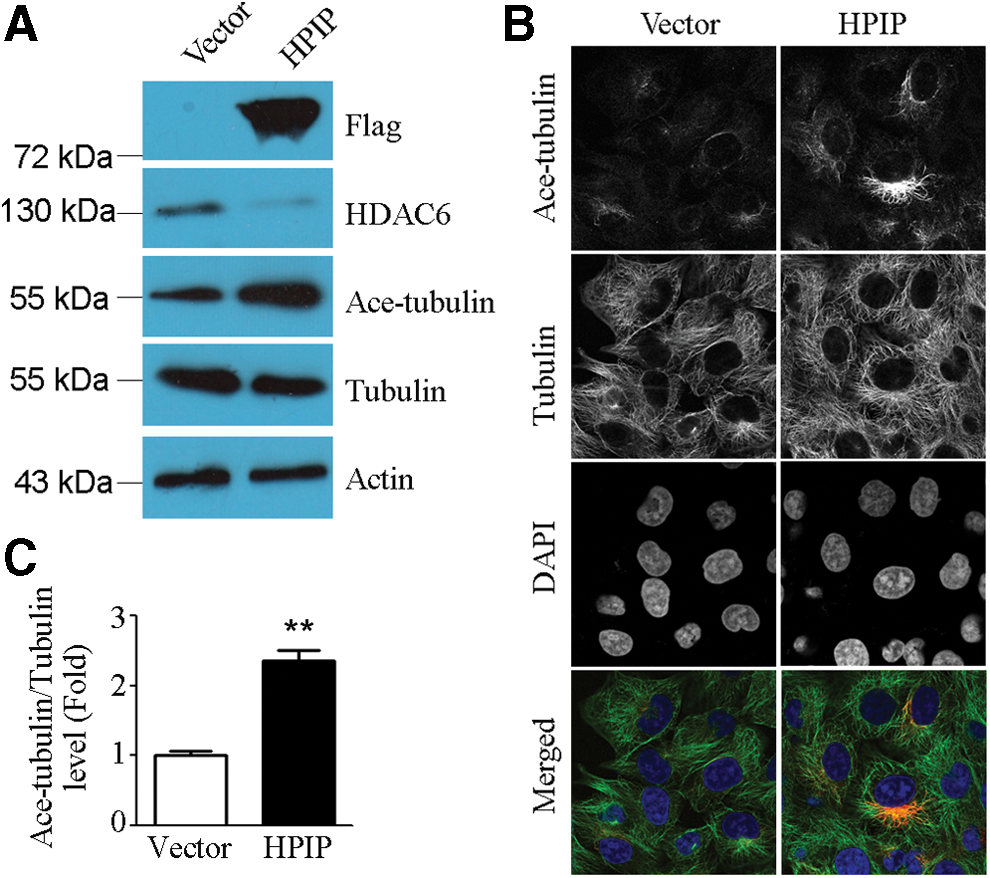
HPIP promoted microtubule stability through suppression of HDAC6 expression.
Discussion
Microtubules are vital for most cellular processes as well as for tumor development in many cancer types, thereby being important targets for chemotherapeutic drugs (Zhou and Giannakakou, 2005). Binding of paclitaxel to tubulin results in enhanced microtubule assembly and stability, which causes mitotic arrest and eventually cell apoptosis. Therefore, cellular factors that regulate microtubule dynamics may have an impact on paclitaxel sensitivity of cancers (Xie et al., 2016). Actually, a number of MBPs have been shown to sensitize cancer cells to paclitaxel or confer a paclitaxel resistance, as exemplified by canonical MBPs, such as EB1 (Mohan et al., 2013), CLIP-170 (Sun et al., 2012), and MCAK (Ganguly et al., 2011), and noncanonical MBPs, for instance, Parkin (Wang et al., 2009) and Survivin (Cheung et al., 2009). Previous studies have demonstrated the oncogenic role of MBP HPIP in many cancers (Wang et al., 2008; Xu et al., 2013a, 2013b; Feng et al., 2015). In this study, from a new perspective, we show that breast cancer cells overexpressing HPIP were more sensitive to paclitaxel treatment. This finding implies that patients with breast cancer exhibiting high level of HPIP might be more responsive to paclitaxel-based regimens, highlighting a clinical value of HPIP in patient selection for paclitaxel chemotherapy.
Although ER-positive breast cancer is reported to be more resistant to paclitaxel than triple-negative breast cancer (Tokuda et al., 2012), our results show that HPIP significantly sensitized ER-positive MCF-7 cells as well as triple-negative MDA-MB-231 cells to paclitaxel, contributing to a roughly 50% decrease on IC50 of paclitaxel with the two cell lines. In contrast, overexpression of HPIP had no effects on the sensitivity of breast cancer cells to the tubulin polymerization inhibitor, vinblastine. This suggests that HPIP is more likely to exert effects through a mechanism related to microtubule stabilization, rather than microtubule catastrophe. Supporting the hypothesis, in vitro assay demonstrated that HPIP further stabilized the microtubule in the presence of paclitaxel, which suggests that HPIP and paclitaxel may act in a synergistic way to facilitate microtubule growth. Next, we show that overexpression of HPIP in breast cancer cells was accompanied with the decreased cellular level of HDAC6 and the increased acetylation of α-tubulin. As a number of studies have demonstrated the regulatory role of HDAC6 in microtubule dynamics by tuning the tubulin acetylation/deacetylation (Boyault et al., 2007; Liu et al., 2015), we infer that an HPIP-HDAC6-tubulin axis may exist in breast cancer, although there is a gap between HPIP and HDAC6 remaining to be filled. Based on the results above, we tend to propose that HPIP might be able to modulate the microtubule network through direct interaction with tubulin or indirectly regulating the acetylation of tubulin. In the presence of both paclitaxel and an overdose of HPIP, they may synergistically enhance microtubule stability to augment paclitaxel-induced mitotic arrest, resulting in breast cancer cells overexpressing HPIP being more sensitive to paclitaxel.
Conclusions
Taken together, our study demonstrates that HPIP sensitizes breast cancer cells in response to paclitaxel treatment, probably through a microtubule-dependent mechanism. Our finding highlights the potential of HPIP as a biomarker predictive of treatment outcomes regarding paclitaxel-based regimens in breast cancer patients. To guide patient selection, further studies are needed to validate the correlation of HPIP level with paclitaxel sensitivity in the clinic.
Footnotes
Disclosure Statement
No competing financial interests exist.