
Abstract
Drug resistance is an obstacle in the chemotherapeutic treatment of lung cancers. In the present study, the effects of high-mobility group box 1 (HMGB1) protein in chemotherapeutic resistance and the relationships between HMGB1 and chemotherapy drug-induced cell apoptosis or necrosis were clarified. We used cisplatin-sensitive A549 cells and cisplatin-resistant A549/DDP cells as cell models with IC50 of 11.58 and 46.95 μM, respectively. A549/DDP had higher level of HMGB1 compared with A549 cells. Interestingly, with the increasing concentration of DDP, HMGB1 was gradually located into cytoplasm in cisplatin-sensitive A549 cells. Moreover, interference with endogenous HMGB1 sensitized the effects of chemotherapeutic drugs, including 5-Fu, DDP, and OXA. Furthermore, results from an in vivo tumorigenesis experiment demonstrated that serum concentration of HMGB1 was much lower in the group inoculated with HMGB1 shRNA-transfected A549 cells than in the N.C. shRNA-transfected A549 inoculated group, as well as the tumor volume, suggesting that serum HMGB1 contributed to tumor growth in a mouse model. In conclusion, higher levels of HMGB1 probably contributed to chemotherapy drug resistance, and higher serum concentration of HMGB1 promoted in vivo tumor growth. The study would provide new clues to overcome drug resistance in chemotherapy of human lung cancers.
Introduction
L
The drug resistance mechanism is relatively complex. A variety of factors contribute to drug resistance, including the expression of multidrug resistance gene and antiapoptotic proteins, inactivation or deletion of proapoptotic proteins leading to tumor tolerance to cell apoptosis, and cancer stem cells enhancing DNA repairing, all of which change the survival signaling pathways to promote cell survival (Shi et al., 2015; Fennell et al., 2016). Thus, the main obstacle to cancer therapy is the acquired drug resistance after a period of chemotherapy (Zhou et al., 2015). Most chemotherapy drugs are toxic and effective to induce cell death of tumor cells, such as necrosis and apoptosis. Platinum drugs are the mostly used chemotherapeutic drugs in the therapy of lung cancers, such as cisplatin (O'Byrne et al., 2011; Wang et al., 2014). To improve the effectiveness of chemotherapy and reduce lifelong side effects, mechanisms of drug resistance are extremely urgent to be explored to seek effective countermeasures to reverse drug resistance.
High-mobility group box 1 (HMGB1) protein is a highly conserved nuclear protein, which plays an important role in maintenance of genomic stability, autophagy, immune regulation, and tumor growth (Pistoia and Pezzolo, 2016). HMGB1 bound to the autophagy regulator Beclin1 and regulated the formation of the Beclin1-phosphatidylinositol 3-kinase class 3 (PI3KC3) complexes to promote cell autophagy in osteosarcoma (Huang et al., 2012). HMGB1 is a critical factor to promote chemoresistance in chemotherapy. Moreover, HMGB1-regulated autophagy is also recognized as a significant contributor to docetaxel resistance in human lung adenocarcinoma (Pan et al., 2014). Besides cell autophagy, apoptosis and necrosis are the two distinct processes of cell death (Long and Ryan, 2012; Walsh, 2014; Li et al., 2016). Till now, the relationship between HMGB1 and cell apoptosis or necrosis has not been extensively studied in human lung carcinomas (Naumnik et al., 2009; Feng et al., 2016.
In the present study, we used two sublines of human lung carcinomas, cisplatin-sensitive A549 cells and cisplatin-resistant A549 cells, as cell models. The effects of HMGB1 protein in chemotherapeutic resistance and the relationships between HMGB1 and chemotherapy drugs were clarified. Therefore, it offers a novel target for improving human lung carcinoma therapy.
Materials and Methods
Cell lines and agents
Human H460 cells (ATCC® HTB177™) and WI-38 cells (ATCC® CCL75™) were purchased from American Type Culture Collection (ATCC). A549 cells and A549/DDP cells were purchased from Cell Bank of Chinese Academy of Medical Sciences. All the cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% of fetal bovine serum (FBS), 100 U/mL of penicillin, and 100 μg/mL of streptomycin. DMEM and FBS were purchased from Hyclone Corporation. The cells were cultured at 37°C in a humidified incubator with 5% CO2 concentration. Cisplatin (Cat. No. 479306) and 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; Cat. No. M2003) were obtained from Sigma-Aldrich. The human HMGB1 shRNAs (Cat. No. TG316576) and control shRNA (Cat. No. TR30013) were obtained from Origen Corporation. Lipofectamine 2000 (Cat. No. 11668-027) was purchased from Invitrogen.
MTT assay
MTT assay was performed in 96-well plates in triplicate. The lung cancer H460 cells were plated at a density of 3 × 103 cells/well overnight and treated with chemotherapy drugs, including 5-Fu, DDP, and OXA, for 24, 48, or 72 h. In another treatment, A549 cells or A549/DDP cells were treated with increasing concentration of DDP for 48 h, and cell viability was determined. IC50 values of DDP were calculated using the GraphPad 5.0 software.
Antibodies
Anti-HMGB1 antibody [EPR3507] (ab79823) was purchased from Abcam Corporation. Mouse monoclonal anti-β-actin antibody (TA310155) was obtained from OriGene Corporation. Lamin B1 antibody (Cat. No. 66095-1-Ig) was purchased from Proteintech. B-Tubulin antibody (H-235): sc-9104 is a rabbit polyclonal IgG provided at 200 μg/mL and obtained from Santa Cruz Biotechnology. Goat anti-rabbit IgG-HRP (Santa Cruz; sc-2004) and goat anti-mouse IgG-HRP (Santa Cruz; sc-2005) were both obtained from Santa Cruz Corporation.
Western blotting analyses
Western blot analyses were performed for detection of endogenous HMGB1 expression. Briefly, cell lysates were prepared in a buffer containing 50mM Tris pH 8.0, 150 mM NaCl, 1% NP-40, 0.5% deoxycholate (DOC), and 0.1% sodium dodecyl sulfate (SDS). The total proteins of each sample were separated by electrophoresis on SDS-polyacrylamide gel and transferred into a nitrocellulose membrane. The membrane was incubated with antibodies against HMGB1 (1:1000), or anti-β-actin as a control, and then incubated with a peroxidase-linked secondary antibody (1:1000) after washing with TBST. Enhanced chemiluminescence was used to visualize each band.
ELISA
The concentration of HMGB1 in cell supernatant or serum was determined with human HMGB1 ELISA kits (E-EL-H1554c; Elabscience) according to the manufacturer's instructions, using a Benchmark Microplate Reader (Bio-Rad). All of the samples were analyzed in duplicate. The concentrations of HMGB1 in the samples were determined from standard curves. For serum samples, whole blood samples were kept at room temperature for 2 h and the supernatant was obtained after 1000 g for 20 min.
In vivo experiment
Athymic nu/nu BALB/c mice (6–8 weeks old) were purchased from Charles River Corporation. The mice were weighed and randomly divided into three groups (n = 6). The body weight was weighed every 5th day. A549 cells were transfected with HMGB1 shRNA and negative control shRNA, and the stable cell lines were screened. HMGB1shRNA-transfected lung cancer cells, N.C. shRNA-transfected cells, or untreated A549 cells (1 × 107 cells/per mouse) were injected subcutaneously to one side of the dorsal flank of the mice. Serum HMGB1 level was examined at 15th day. The experiment was approved by the Committee for Animal Experimentation of Central Hospital of Tianjin.
Results
HMGB1 level in A549/DDP cells is higher than that in A549 cells
To detect whether HMGB1 promotes chemotherapy resistance, we used the cisplatin-sensitive cell line A549 and cisplatin-resistant cell line A549/DDP as cell models. First, A549 cells and A549/DDP cells were treated with increasing concentrations of DDP for 48 h, and cell viability was determined by MTT assay. The IC50 values were calculated by GraphPad 5.0 software and IC50 value of DDP in A549 cells and A549/DDP cells was 11.58 μM and 46.95 μM, respectively (Fig. 1A). Moreover, the levels of HMGB1 were also tested by western blotting analysis. As shown in Figure 1B, the expression level of HMGB1 in A549/DDP cells was obviously higher than that in A549 cells. All of the data suggested that high levels of HMGB1 probably contributed to chemotherapy drug resistance.
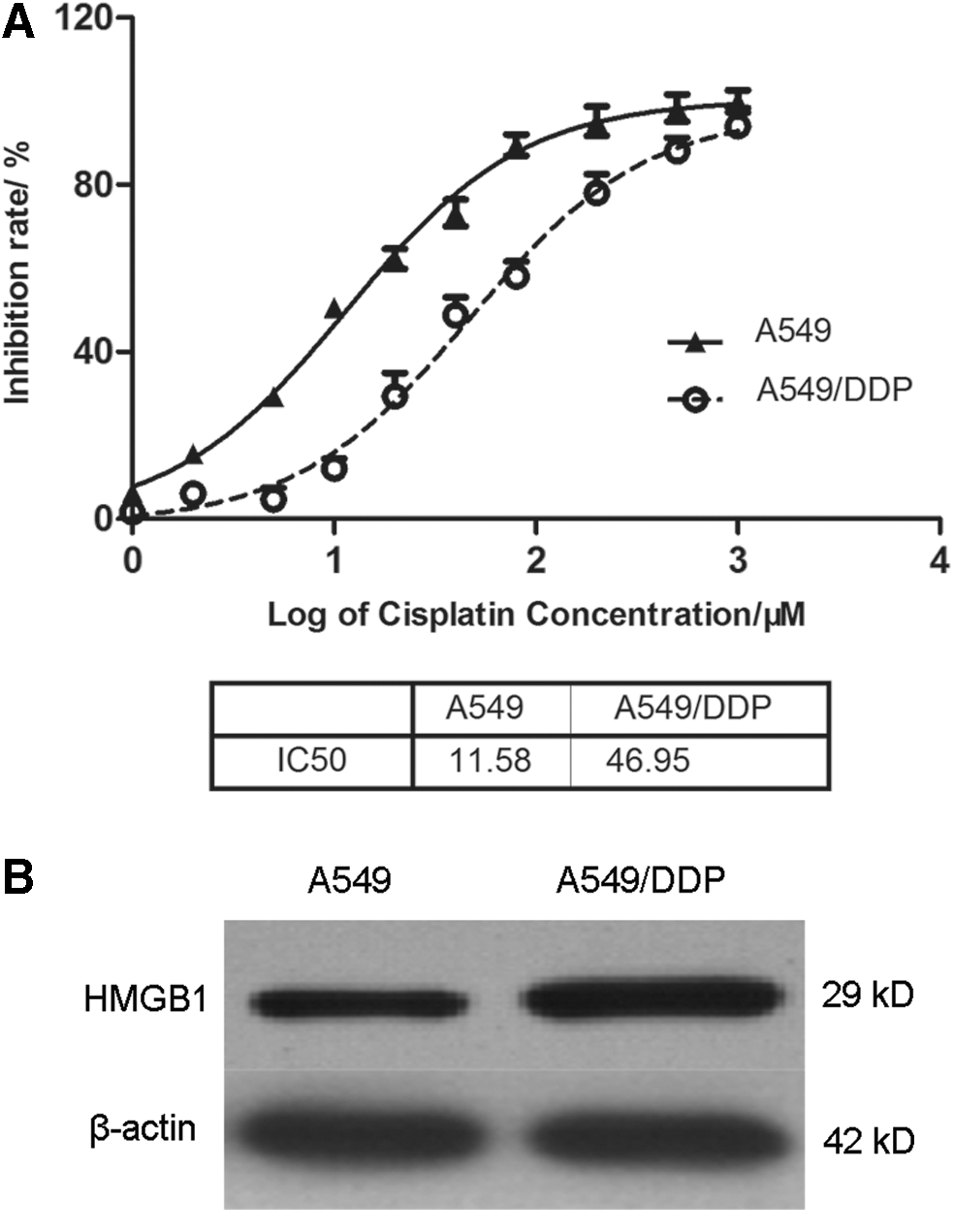
HMGB1 level in A549/DDP cells is higher than that in A549 cells.
DDP increases the levels of HMGB1 in cytoplasm of A549 cells
To detect whether DDP treatment affects the HMGB1 location in A549 cells and A549/DDP cells, A549 cells and A549/DDP cells were treated with different concentrations of DDP for 48 h. The DDP concentration used here was 0, 5, and 10 μg/mL, respectively. The cytoplasmic and nuclear proteins were extracted according to kit protocols. As shown in Figure 2, the cytoplasmic and nuclear HMGB1 amounts in A549 cells were gradually increased as the increasing concentration of cisplatin. Moreover, the abundance of HMGB1 in the cytoplasm was higher compared with that in the nucleus. However, the levels of cytoplasmic and nuclear HMGB1 in A549/DDP cells were not obviously changed as the increasing concentrations of cisplatin. The results demonstrated that DDP treatment promoted the cytoplasmic location of HMGB1 in cisplatin-sensitive A549 cells and had no obvious effects in the cisplatin-resistant A549/DDP subline.
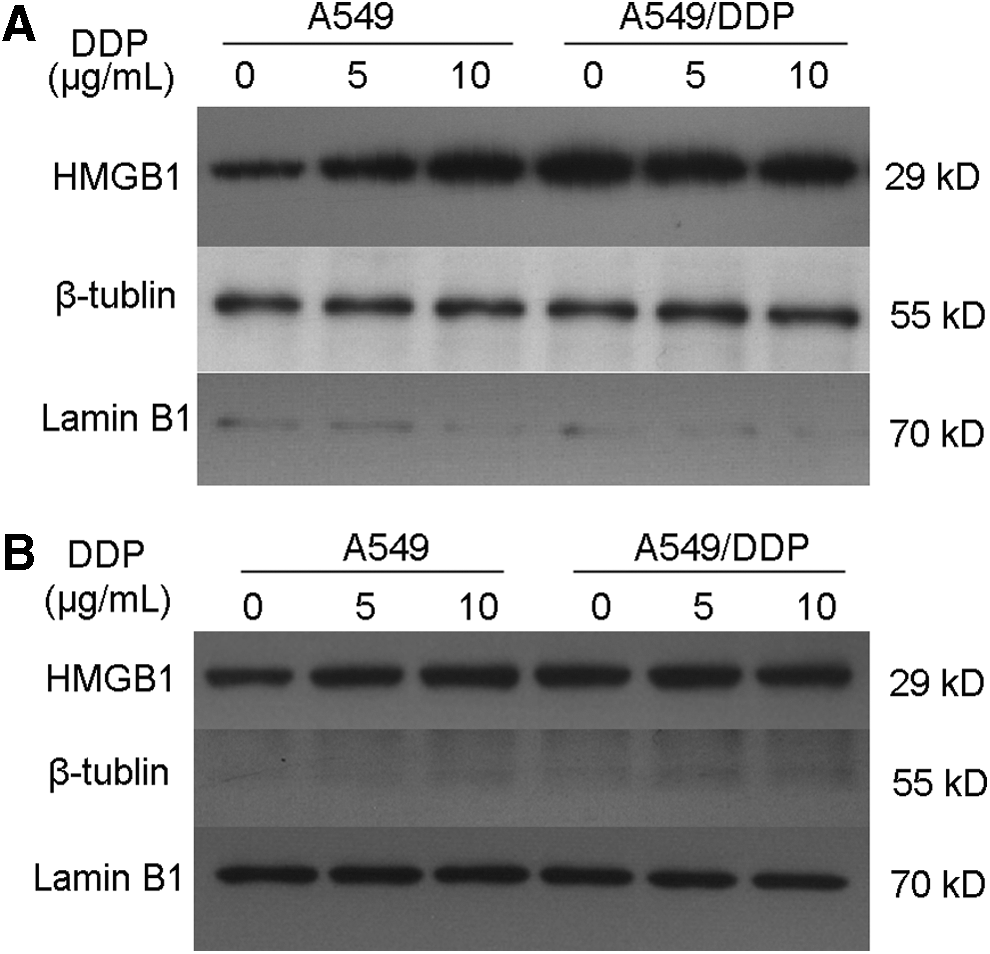
DDP increases the levels of HMGB1 in cytoplasm of A549 cells. A549 cells and A549/DDP cells were treated with different concentrations of DDP for 48 h. The DDP concentration was 0, 5, and 10 μg/mL, respectively. The cytoplasmic and nuclear proteins were extracted according to the kit protocol. The levels of HMGB1 in cytoplasm
Interference with endogenous HMGB1 sensitizes the effects of chemotherapeutic drugs
It has been identified that cisplatin-resistant A549/DDP cells had higher levels of HMGB1 than in cisplatin-sensitive A549 cells. WI-38 is normal human diploid cell from normal embryonic (3 months of gestation) lung tissue. We first tested the expression levels of HMGB1 in lung cancer cell lines, H460, A549, A549/DDP, and WI38 cells, by western blotting analysis. As shown in Figure 3A, the results demonstrated that HMGB1 levels in A549 cells, A549/DDP cells, and H460 cells were obviously upregulated than that in WI38 cells. Next, we used shRNA specific to HMGB1 to knockdown the endogenous HMGB1 in H460 cells, which was determined by western blotting analysis (Fig. 3B).
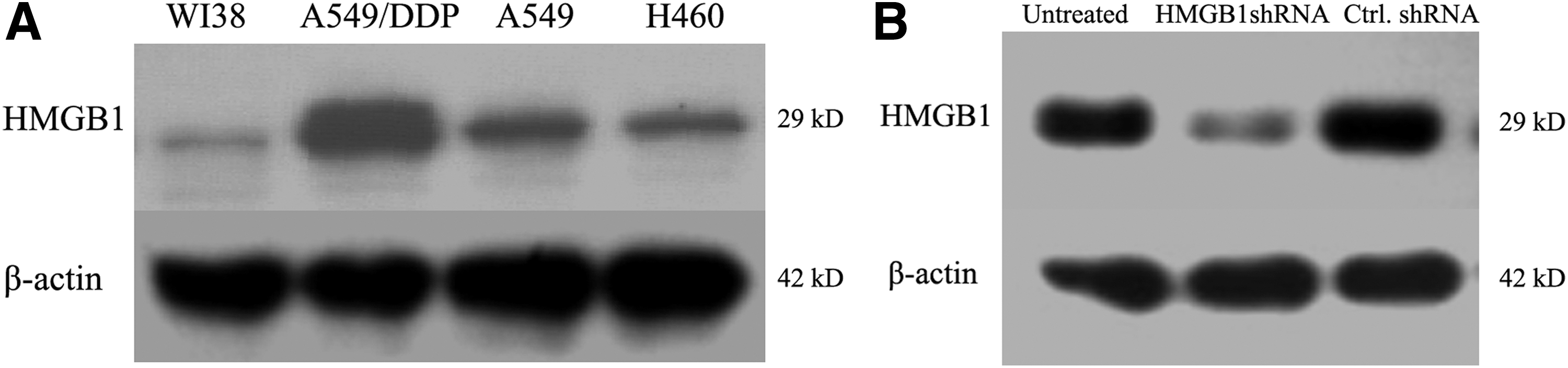
The expression levels of HMGB1 were detected.
To determine whether interference with HMGB1 would sensitize the effects of chemotherapeutic drugs, we used three kinds of chemotherapy drugs, 5-Fu, DDP, and OXA, to treat the cells transfected with HMGB1 shRNA or control shRNA. As shown in Figure 4A, H460 cells were transfected with HMGB1 shRNA and control shRNA and treated with increasing concentrations of 5-Fu for 24, 48, and 72 h, respectively. The results showed that the cell viability was significantly suppressed in H460 cells transfected with HMGB1 shRNA, compared with that transfected with control shRNA, which was consistent with that of H460 cells treated with ADM (Fig. 4B) and OXA (Fig. 4C). All of the data demonstrated that transfection with HMGB1 shRNA in H460 cells significantly inhibited cell proliferation and sensitized the effects of chemotherapeutic drugs.
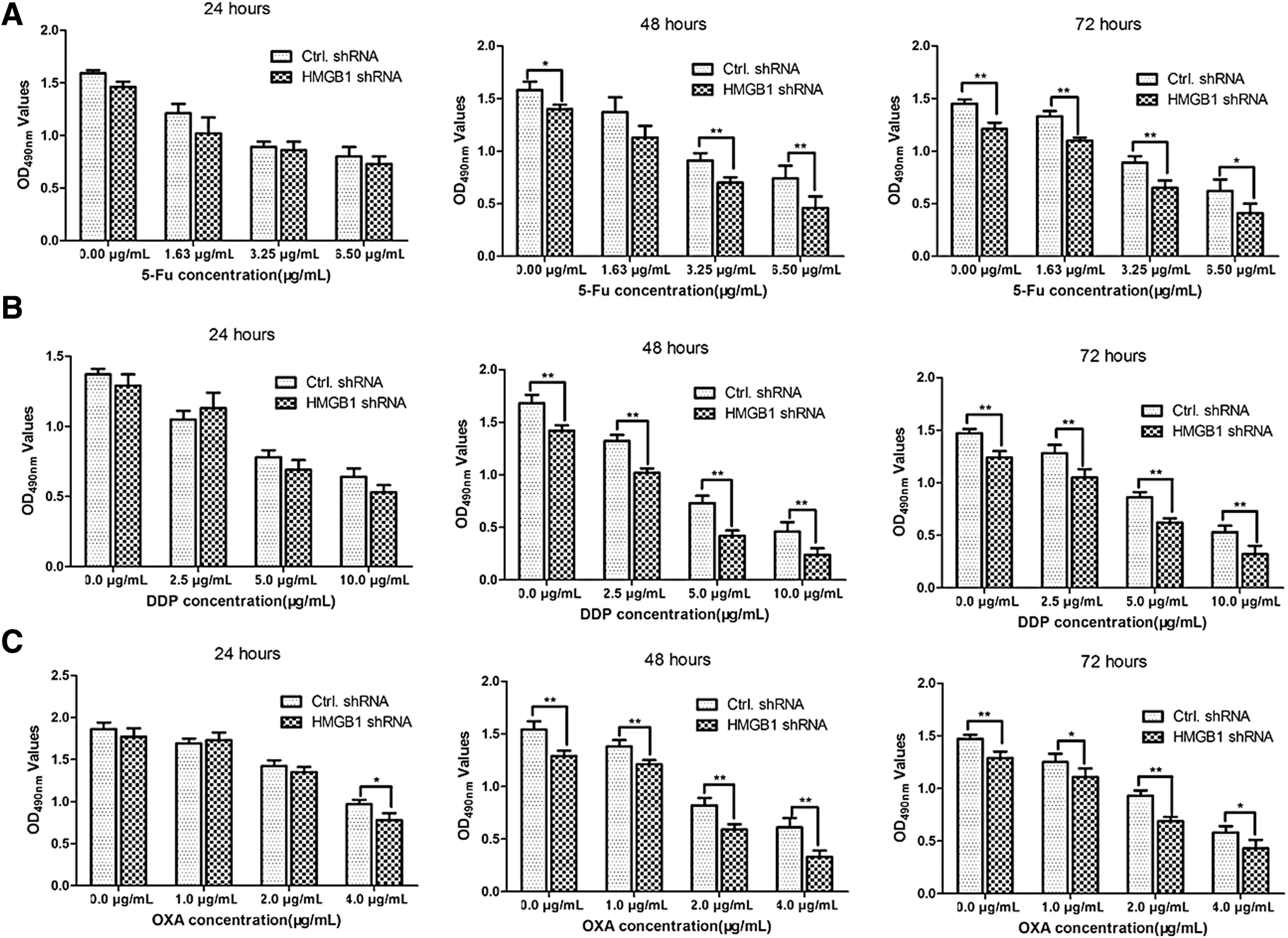
Interference with endogenous HMGB1 sensitizes the effects of chemotherapeutic drugs. H460 cells were transfected with HMGB1 shRNA and negative control shRNA for 48 h. The H460 cells transfected with HMGB1 shRNA or control shRNA were treated with increasing concentrations of 5-Fu
Knockdown of HMGB1 suppresses tumor growth in a mouse tumor model
To further examine the effect of HMGB1 in the progression of lung cancers, athymic nu/nu BALB/c mice were injected with HMGB1 shRNA-transfected A549 cells, negative control shRNA-transfected A549 cells, and untreated A549 cells (1 × 107 cells/per mouse) on the back of the mice at 0th day (Fig. 5A). The tumors were gradually seen on the 5th day in the group inoculated with N.C. shRNA-transfected A549 cells or untreated A549 cells. The tumors in the group inoculated with HMGB1 shRNA-transfected stable cells were seen since the 7th day. The body weight of the mice in every group was monitored and there was no statistical difference between the groups (Fig. 5B). Moreover, the serum concentration of HMGB1 was determined by ELISA and the result demonstrated that the mice inoculated with HMGB1 shRNA-transfected A549 cells had a higher level of HMGB1 in serum compared with the negative control shRNA group or untreated group (Fig. 5C). Furthermore, the growth of tumors in the HMGB1 shRNA-transfected A549 group was much slower than that in the N.C. shRNA-transfected A549 group (**p < 0.01, Fig. 5D). All of the data revealed that higher levels of serum HMGB1 contributed to lung carcinoma growth in the mouse model.
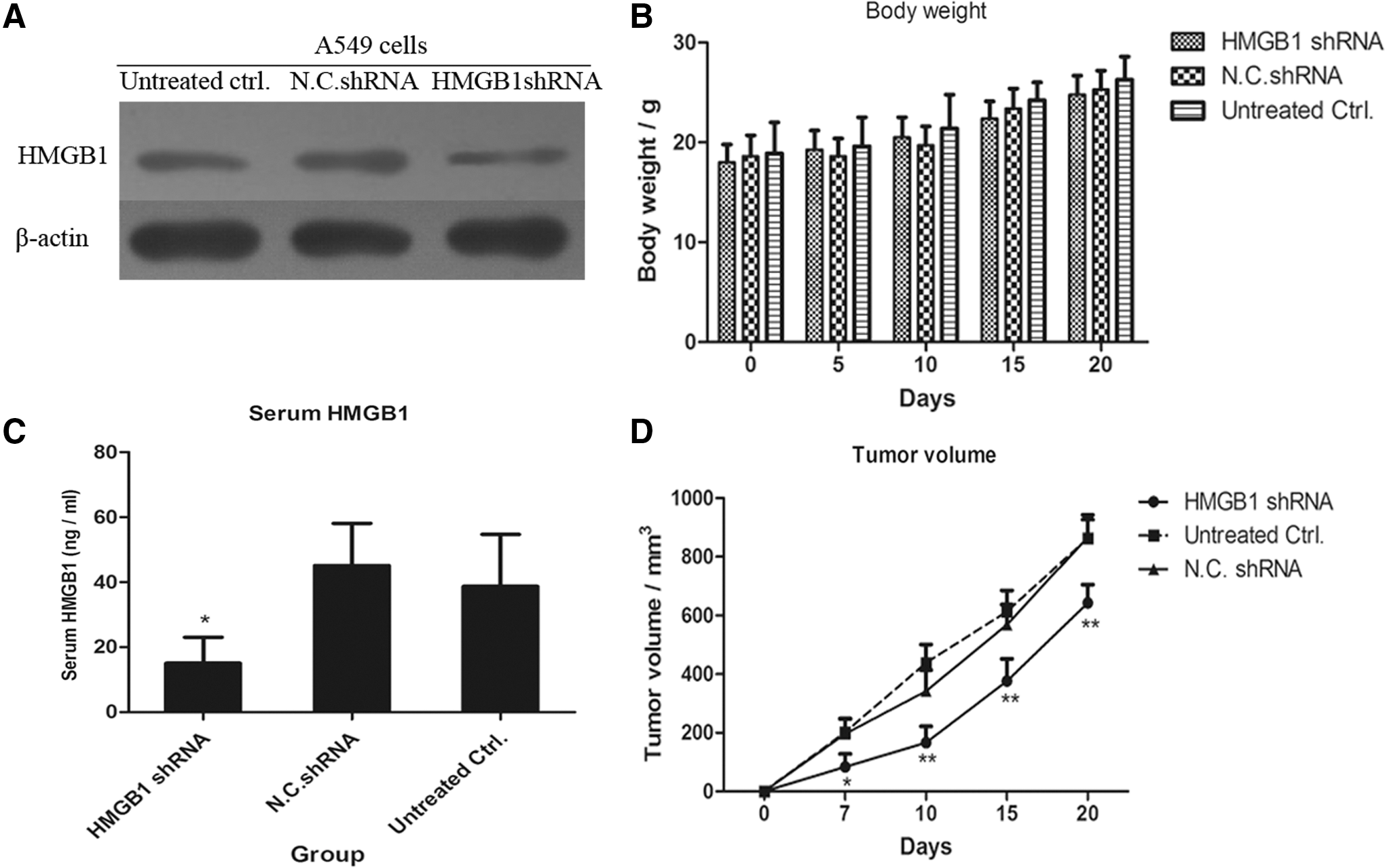
Knockdown of HMGB1 suppresses tumor growth in a mouse tumor model.
Discussion
Lung carcinoma has been the most common cancer with high incidence and poor prognosis in the world for several decades (Koletsis et al., 2009; Dzian et al., 2012). Conventional chemotherapeutic agents usually elicit both apoptosis and necrosis of tumor cells (Vilanova-Costa et al., 2014; Yang et al., 2015). They could effectively induce cell death in early time, but chemotherapeutic drug resistance develops after a period of chemotherapy in the majority of patients in clinical therapy (Hassan et al., 2016; Tsao et al., 2016).
In the present study, the relationship between HMGB1 expression and chemotherapy resistance in lung cancer cells was investigated. The expression level of HMGB1 was detected by western blotting analysis and the results revealed that HMGB1 levels were significantly higher in the cisplatin-resistant cell line, A549/DDP, compared with the cisplatin-sensitive cell line, A549, as well as the HMGB1 concentration in cultured medium. Normally, HMGB1 is the most important chromatin protein located in the nucleus. We extracted the cytoplasmic and nuclear proteins, respectively, and detected the levels of HMGB1 in cisplatin-treated lung cancer cells. The results demonstrated that with the increasing concentrations of cisplatin, the levels of HMGB1 in cytoplasm gradually increased, suggesting that cisplatin treatment might promote HMGB1 translocation to cytoplasm of A549 cells, which might contribute to chemotherapy resistance. Furthermore, interference with HMGB1 by shRNA specific to HMGB1 sensitized the effects of chemotherapeutic drugs, including 5-Fu, DDP, and OXA. Therefore, knockdown of the expression of HMGB1 could reduce chemotherapy drug resistance, increase chemotherapy sensitivity, and finally enhance the therapeutic effects of chemotherapeutic drugs in human lung cancer cells. This was consistent with Huang, J.'s finding that knockdown of HMGB1 restored the chemosensitivity of osteosarcoma cells in vivo and in vitro (Huang et al., 2012). Additionally, a related study showed that suppression of HMGB1 by siRNA could reverse resistance to ADM and increase the cell apoptotic rate in K562/A02 cells (Xie et al., 2008).
Thus, from our findings, we concluded that HMGB1 contributed to chemotherapeutic drug resistance. Interference with the endogenous HMGB1 expression or inhibition of cytoplasmic translocation of HMGB1 may reverse the drug resistance and increase chemotherapy sensitivity. The study may establish an effective chemotherapy choice and provide a new drug target in the clinical therapy of lung carcinomas.
Footnotes
Disclosure Statement
No competing financial interests exist.