
Abstract
UDP-galactose-4-epimerase (UGE) is a key enzyme involved in galactose metabolism by catalyzing the interconversion of UDP-glucose to UDP-galactose. The cDNA encoding UGE was isolated from Phyllostachys edulis by reverse transcription–polymerase chain reaction and by 5′ and 3′ rapid amplification of cDNA ends, and was designated as PeUGE. The full-length cDNA of PeUGE was 1778 bp, which contained an open reading frame (ORF) encoding a peptide of 420 amino acids, with a calculated molecular mass of 46.58 kDa and a theoretic isoelectric point of 9.07. The genomic sequence corresponding to the ORF of PeUGE was 2656 bp containing 10 exons separated by nine introns. Tissue-specific analysis showed that PeUGE was constitutively expressed with the highest level in shoots, which had an increasing trend with the growth of shoots. PeUGE was induced by abiotic stresses such as drought, salinity, and water stresses. Moreover, chlorophyll fluorescence parameters and lateral roots analysis of transgenic Arabidopsis thaliana plants overexpressing PeUGE systematically confirmed the crucial role of PeUGE in improving the tolerance to abiotic stresses. These results indicated that PeUGE might be one of the key genes involved in the biosynthesis of cell wall polysaccharides during the growth and development of bamboo and in response to stresses, which provided a candidate gene for molecular engineering to improve the quality of bamboo products.
Introduction
D
Plant UGE isoforms function in different metabolic situations and their enzymatic properties, gene expression pattern, and subcellular localization contribute to the differentiation of isoform function (Barber et al., 2006). The UGE1 activity in UGE1 overexpressing Arabidopsis plants increased by up to 250%, while that in UGE1-suppressed plants was decreased to 10% compared with the wild type (Dörmann and Benning, 1998). Loss-of-function mutations in UGE4 were found to induce dramatic morphological alterations in roots and correlated with a reduction of cell wall-bound galactose without affecting the level of galactolipids (Seifert et al., 2002). The expression of UGE was induced by various abiotic stresses, such as cold, drought, high salinity, and ABA (Seki et al., 2002; Rabbani et al., 2003). Overexpression of UGE gene can increase the content of raffinose in transgenic plants and boost stress relief, without altering plant morphology (Liu et al., 2007). All these findings indicated that UGE may play an important role in galactose metabolism and in the biosynthesis of carbohydrate content, improving the tolerance to abiotic stresses.
Moso bamboo (Phyllostachys edulis) is one of the most fast-growing plants in the world and represents a significant ecological, economic, and cultural value in China. It can grow up to more than 100 cm in only 24 h and reach to its maximum height of about 24 m during one and a half months at growth spurt in spring (Peng et al., 2013). Both cell division and elongation in individual internodes of bamboo dominate the development of culm at different growing stages (Cui et al., 2012). Apparently, cell wall synthesis and energy supply are indispensable for maintaining normal physiological function during the rapid growth of bamboo. On the other hand, abiotic stresses always cause serious influence on the growth and development of bamboo. Although a large number of UGEs in plants have been extensively studied through expression and functional analysis (Seki et al., 2002; Rabbani et al., 2003; Liu et al., 2007), our knowledge of UGE in bamboo is still limited. It is unknown whether UGE is involved in biosynthesis of cell wall at the fast growing stages and tolerance to abiotic stress in bamboo.
In this study, a full-length cDNA encoding PeUGE was isolated from moso bamboo (P. edulis). The expression patterns of PeUGE in different tissues, especially in shoots at different stages during the rapidly growing period, as well as in roots under drought, salinity, and water stresses were characterized. PeUGE was overexpressed in A. thaliana, and the phenotype and physiological parameter of the transgenic plants under stresses were investigated. A detailed analysis of the chlorophyll fluorescence parameters and lateral roots with the transgenic plants will lead to a better understanding of the PeUGE function in P. edulis, which will facilitate further strategies for the directional improvement of bamboo suitability and shoot quality by molecular engineering.
Materials and Methods
Plant material and growth conditions
One-year-old moso bamboo (P. edulis) seedlings were potted in our laboratory under long-day conditions (16-h light/8-h dark) at 28°C, with a light intensity of 200 μmol·m−2·s−1. The air relative humidity was about 50%. Roots, stems, mature leaves, immature leaves, leaf sheaths, and shoots were collected and quickly frozen in liquid nitrogen for further analyses.
The bamboo seedlings were taken out from pot and cleaned with sterile water for different treatments. For water treatment, seedlings were incubated in sterile water for 2 h. For salinity stress assay, the roots were submerged into the solution of 400 mM NaCl for 2 h (Sun et al., 2016). For drought treatment, the seedlings were exposed to the air for 2, 3, and 4 h. Roots were collected from the seedlings after treatments, respectively.
Identification of UGE genes in moso bamboo
BLASTN was performed to identify putative UGE genes in moso bamboo using the bamboo genome database (BambooGDB,
Total RNA isolation and gene cloning
Total RNA was isolated from different tissues of moso bamboo seedlings and the roots after treatments as mentioned above using Trizol reagent solution (Invitrogen), following the manufacturer's protocol. All samples collected from each treatment were at least three individuals with biological triplicates. The total RNA was treated with RNase-free DNase (Promega) for 30 min at 37°C and the absence of DNA was verified by polymerase chain reaction (PCR) using the primer of PeEF1 (Elongation factor 1, GenBank: FP095360.1) (Sun et al., 2016). The integrity of total RNA was verified through agarose gel electrophoresis, as well as the purity and concentration of RNA was determined by Nanodrop 2000 (Thermo). First strand of cDNA was synthesized by reverse transcription system (Promega). For each 20 μL reaction, 1000 ng of total RNA were used, the synthesis was carried out at 42°C for 45 min and 99°C for 5 min, and the final cDNA product was diluted fivefold before use.
The conserved sequence of PeUGE was amplified using the primers (PeUGE-F: 5′-ATGCTGCCCACCAACAGGAGC-3′ and PeUGE-R: 5′-CAAAATCATGGCCTGCGGC-3′), based on the sequence of OsUEL-1 (AB096864) in rice. The specific 5′ rapid amplification of cDNA ends (RACE) and 3′ RACE primers were designed on the basis of the conserved sequence. The primers used for 5′ RACE were 5-1 (5′-TCAGTGATAGGCATCTTCTCAGGCTCTC-3′) and 5-2 (5′-CAGCAGCCGAAGAGCAGCATGTGAACC-3′) and those for 3′ RACE were 3-1 (5′-TGACTATGAGACGGCTGATGGAACTTGC-3′) and 3-2 (5′-GTAGGTCAGTGAAGGAGTTTGTGGAAGC −3′). Touchdown PCR was performed with 5-1, 3-1, and a universal primer mix (UPM; as supplied with the SMARTTMRACE cDNA Amplification Kit [TakaRa, Japan]). The PCR amplicons from primer pairs of 3-1 and UPM were used as template for a subsequent nested PCR using primer pairs of 3-2 and the NUP primer supplied in the kit, and those of 5-1 and UPM were used for another subsequent nested PCR with 3-2 and NUP. PCR products were cloned into pGEM T-easy vector (Promega) using standard protocol and sequenced using an ABI 3730 sequencer (Applied Biosystems). The full-length cDNA was obtained by the combination of conserved sequence with the 5′ and 3′ end sequences.
Genomic DNA was extracted from leaves of P. edulis using CTAB method (Gao et al., 2006). The cDNA and genomic DNA were used as templates, respectively, in PCR, which were performed with the primer pairs of PeUGE-F and PeUGE-R using PrimerSTAR polymerase (Takara, Japan). The promoter region sequence upstream the open reading frame (ORF) of PeUGE was also isolated with leaf genomic DNA using primer pairs of PeUGE-F-pro (5′-ATCGGTACCCCAGCCTAGTGGAC-3′) and PeUGE-R-pro (5′-CTGCTCCTGTTGGTGGGCAGC-3′). The amplified products were inserted into the pGEM-T easy vector and subsequently confirmed by sequencing.
Bioinformatic analysis of PeUGE
The cDNA and genomic DNA sequence of PeUGE were analyzed by VectorNTI, the gene structure, including the number of introns and the position of each intron, was shown using WinPlas (Version 2.7). Sequences homology searches were performed using BLASTN with default parameters (
Tissue-specific gene expression analysis
For the analysis of gene expression in different tissues, the total RNAs isolated from roots, stems, mature and immature leaves, leaf sheaths, and shoots of bamboo seedlings were used for cDNAs synthesis, respectively. Semiquantitative reverse transcription polymerase chain reaction (RT-PCR) was conducted with primers of PeUGE-F and PeUGE-R. The final volume was 20 μL, including 2 μL of 10 × LA Taq Buffer (Mg2+ Plus), 1.6 μL of dNTP Mixture (2.5 mM each of dATP, dTTP, dCTP, and dGTP), 1.0 μL of PeUGE-F and PeUGE-R (5 μM, each), 1 μL cDNA, 13.2 μL ddH2O, and 0.2 μL LA Taq DNA polymerase (Takara). The PCR program involved an initial denaturation period at 95°C for 4 min, followed by 26 cycles at 95°C for 30 s, 62°C for 30 s, and then 72°C for 80 s; after the last cycle, there was a final extension period at 72°C for 10 min. The cDNA fragment of PeActin (GenBank: GU434145) was amplified using PeActin-F (5′-GATCTTGCTGGGCGTGACCTC-3′) and PeActin-R (5′-CCATCGGGCATCTCGTAGC-3′) as a positive control under the same PCR condition.
Real-time quantitative PCR analysis
Quantitative real-time RT-PCR (qRT-PCR) was performed on qTOWER2.2 (Analytik Jena, Germany) with Roche LightCycler® 480 SYBR Green 1 Master kit, according to the procedure of 95°C for 5 min; 95°C for 10 s, 63°C for 15 s, 40 cycles. The reaction volume was 10 μL containing 5.0 μL of 2 × SYBR Green 1 Master, 0.8 μL cDNA, 0.2 μL of primer PeUGE-QF (5′-AGAGAGAGGGAACCAGGACCAAT-3′) and PeUGE-QR (5′-AGATTGCTCCAGCAGCAGCAC-3′) (5 μM, each), and 3.8 μL ddH2O. For each condition, the qRT-PCR experiments were performed in biological triplicate with three technical replicates. Gene expression was calculated with the 2−ΔΔCt method (Livak and Schmittgen, 2001) using two reference genes, Nucleotide Tract-Binding protein gene (NTB, GenBank: FP091427) and Tonoplast Intrinsic Protein gene (TIP41, GenBank: FP092936) (Fan et al., 2013).
Plant transformation and generation of transgenic plants
The coding sequence of PeUGE was amplified to introduce an XbaI site upstream of the start codon and a BamHI site downstream of ORF. The PCR fragment was then ligated into pPZP vector digested with XbaI and BamHI to generate the CaMV 35S::PeUGE-GFP construct, which was verified by sequencing. The specific primers used to generate this XbaI- BamHI fragment were PeUGE-F1 (5′-
The construction of CaMV 35S::PeUGE-GFP was introduced into Agrobacterium tumefaciens (strain EHA105) by electroporation. Transformations of Arabidopsis plants were conducted using the floral dip method (Clough and Bent, 1998). Putative transgenic plants harboring PeUGE were selected on a medium supplemented with 50 mg·L−1 kanamycin. The kanamycin-resistant seedlings were further confirmed by RT-PCR amplification using the primers PeUGE-F1 and PeUGE-R1. T3-resistant seedlings showing no evidence of segregation were selected for further analysis. The expression of PeUGE in two independent T3 lines was investigated by RT-PCR analysis using the primer pairs of PeUGE-F and PeUGE-R. AtUbiquitin (NM180850) was selected as an internal control under the same PCR conditions using primers of AtUbiquitin-F (5′-ATGGCTGAAGAGGATATCCAGC-3′) and AtUbiquitin-R (5′-GAAACACTTCATATGGACGATGG-3′) (Yang et al., 2015). qRT-PCR was also conducted to find the relative expression quantity of PeUGE in the transgenic plants using the primer pairs of PeUGE-QF1 (5′-ATGAGACGGCTGATGGAACT-3′) and PeUGE-QR1 (5′-GCCTCTTTCTGCCTTGTTGA-3′), and AtUBQ (NM_116771) (Chen et al., 2015) was selected as the reference gene.
Drought and salinity stress treatments
The seeds of A. thaliana ecotype Columbia (Col-0) and transgenic lines (L5 and L6) were cultured in the MS medium and vernalized at 4°C for 2 days in darkness. The germinated seeds were maintained under a 16-h light/8-h dark cycle at 22°C for 1 week, and then both transgenic and Col-0 plants were transferred to a fresh MS medium supplemented with 100 mM mannitol or 50 mM NaCl for vertical plate culture. After 2 weeks of incubation, the numbers of lateral root were monitored with a binocular microscope directly. At the same time, another set of transgenic and Col-0 plants was transplanted to containers filled with a mixture of soil and vermiculite (1:1) where they were regularly watered for 2 weeks. For stress tolerance analysis, Arabidopsis plants were irrigated with 10% PGE-6000 and 200 mM NaCl for 8 days, respectively. Chlorophyll fluorescence parameters of transgenic and Col-0 plants were measured using Dual-PAM-100 (Walz, Germany). Both the maximum quantum yield in Photosystem II (F v/F m) and the nonphotochemical quenching (NPQ) were used to estimate the stress degree of Arabidopsis plants (Maxwell and Johnson, 2000). Parameters were measured for 30 replicates per treatment per line. Phenotype of transgenic plants was observed and photographed simultaneously.
Statistical methods
Student's t-test was used for the statistical analysis of the unit lateral root numbers, the data are presented as mean value ± standard deviation. For the analyses of qRT-PCR data and the fluorescence parameter values, software SPSS ver. 19.0 was used. Statistical significance was evaluated by one-way analysis of variance. p < 0.05 was considered significant.
Results
Identification, isolation, and sequence analysis of PeUGE
Based on the UGE homologous sequence analysis, five putative UGE sequences in the genome of P. edulis were mined from BambooGDB, including PH01000264G0930, PH01000300G0530, PH01000313G1050, PH01000331G1010, and PH01000338G0970. Further analysis showed that PH01000331G1010 had the closest genetic relationship with OsUGE1, which had the function of altered tolerance to abiotic stress and cell wall carbohydrate partitioning in O. sativa (Liu et al., 2007; Guevara et al., 2014). UGE homologue genes in plant species with closer genetic relationship have higher similarities, which facilitate the designing of primers for RT-PCR to isolate the cDNA sequence (PeUGE) of PH01000331G1010. As a result of the RT-PCR with the primer pairs of PeUGE-F and PeUGE-R, a 1260-bp nucleotide fragment was obtained. Subsequently, a 527-bp fragment from the 5′-end region and a 506-bp fragment from 3′-end region were obtained by 5′ and 3′ RACE, respectively. After analysis of the obtained sequences, a 1778-bp full-length cDNA was identified, containing a 1263-bp ORF, a 251-bp 5′-untranslated region (UTR), and a 264-bp 3′-UTR (GenBank: KX227715). Finally, the ORF and the genomic sequence corresponding to the ORF were determined by end-to-end PCR using cDNA and genomic DNA as template, respectively. The genomic sequence corresponding to the ORF was 2656 bp containing 10 exons separated by nine introns (Supplementary Fig. S1; Supplementary Data are available online at
Besides, the promoter region sequence upstream of the ORF of PeUGE was obtained from leaf genomic DNA using primer pairs of PeUGE-F-pro and PeUGE-R-pro, which was 1539 bp containing essential promoter elements, such as TATA-box and the CAAT-box. Some stress responsive elements, such as ABRE, CGTCA-motif, HSE, and MBS, as well as meristem expression elements of CAT-box and CCGTCC-box were also identified in the promoter regions (Supplementary Table S1), indicating that PeUGE may be involved in stress responses and meristem growth.
Phylogenetic analysis of PeUGE
The deduced amino acid sequence of PeUGE was analyzed with homologous proteins in other organisms (Supplementary Fig. S2). Multiple sequence alignment demonstrated that the sequence encoded by PeUGE shared 94% homology with that of Brachypodium distachyon and rice, and more than 80% homology with those of monocots such as Setaria italica, Sorghum bicolor, and Triticum aestivum, which indicated that UGE was highly conserved during evolution. A Neighbor-Joining phylogenetic tree was constructed based on the homologous sequences of UGE. The clustering data clearly showed that UGEs were mainly clustered into three groups by their origin of higher plants, algae, and bacteria, respectively (Fig. 1). As bacteria, Methanothermobacter thermautotrophicus, Leptolyngbya valderiana, and Escherichia coli were found to form a clade distinct from the plants, P. edulis and other monocots were clustered in the same clade close to that of dicots, which was away from those of moss, fern, and algae successively. This result is consistent with the findings in the stated phylogeny and classification of plants.
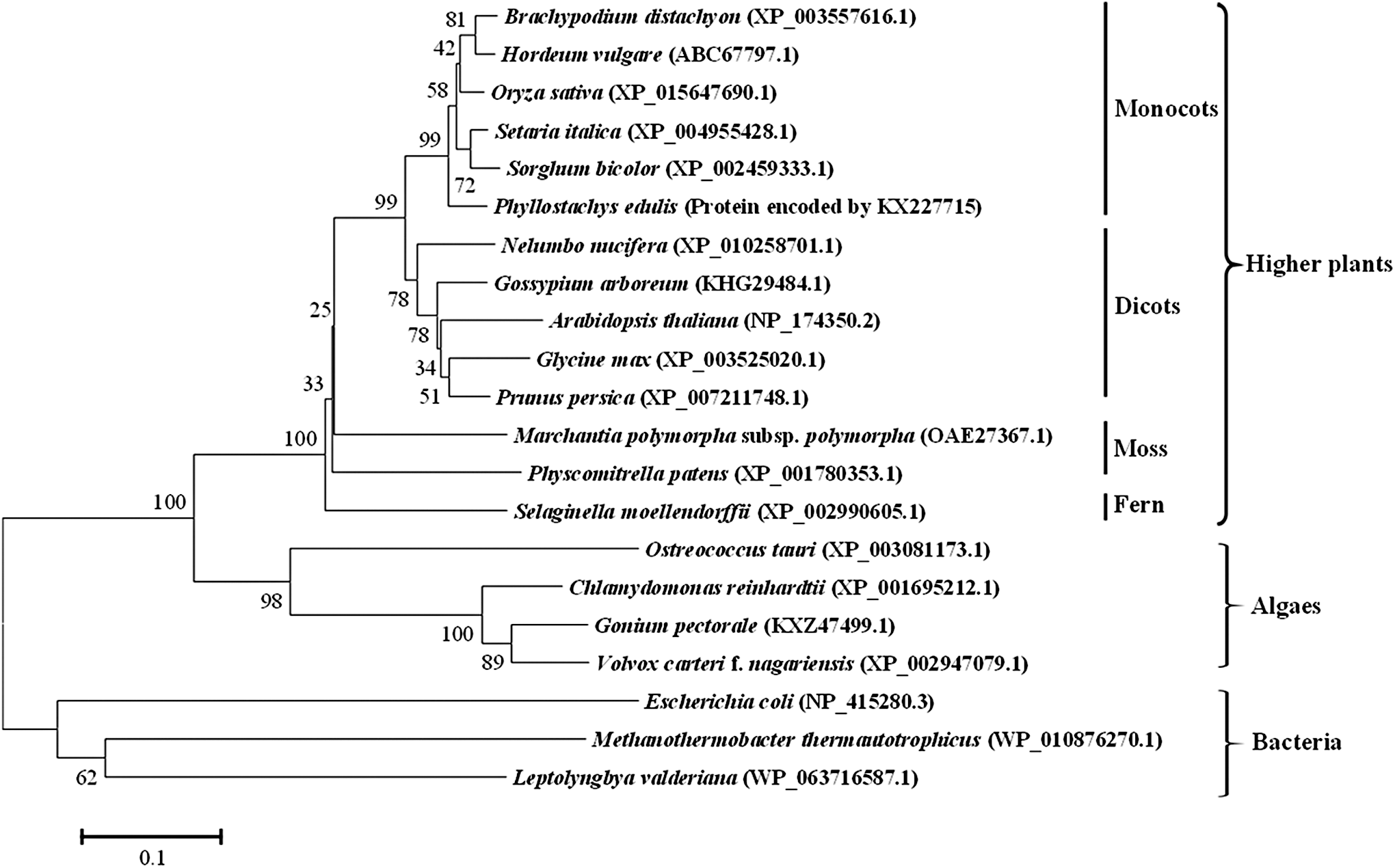
Phylogenetic analysis of PeUGE with other UDP-galactose-4-epimerases (UGEs) from different organisms based on amino acid sequences by MEGA 6.0. Numbers above major branches indicated bootstrap value estimates for 10,000 replicates. Three clusters were illustrated by their origin of higher plants, algae, and bacteria, respectively. A total of 21 UGEs used in phylogenetic tree are PeUGE encoded by KX227715 from Phyllostachys edulis, NP_174350.2 from Arabidopsis thaliana, XP_003557616.1from Brachypodium distachyon, XP_015647690.1 from Oryza sativa, XP_003525020.1 from Glycine max, KHG29484.1 from Gossypium arboreum, ABC67797.1 from Hordeum vulgare, XP_010258701.1 from Nelumbo nucifera, XP_007211748.1 from Prunus persica, XP_004955428.1 from Setaria italica, XP_002459333.1 from Sorghum bicolor, OAE27367.1 from Marchantia polymorpha subsp. polymorpha, XP_001780353.1 from Physcomitrella patens, XP_002990605.1 from Selaginella moellendorffii, XP_001695212.1 from Chlamydomonas reinhardtii, KXZ47499.1 from Gonium pectorale, XP_002947079.1 from Volvox carteri f. nagariensis, NP_415280.3 from Escherichia coli, WP_010876270.1 from Methanothermobacter thermautotrophicus, and WP_063716587.1 from Leptolyngbya valderiana.
Expression patterns of PeUGE
To analyze PeUGE gene expression in different tissues, semiquantitative RT-PCR was conducted using the cDNA templates of roots, stems, mature and immature leaves, leaf sheaths, and shoots of P. edulis. The results indicated that PeUGE was detected in all tissues with diverse patterns (Fig. 2). Interestingly, we found PeUGE was expressed at the highest level in shoots, followed by stems, and little in leaves and roots, indicating that it might play an important role in the development of shoots.
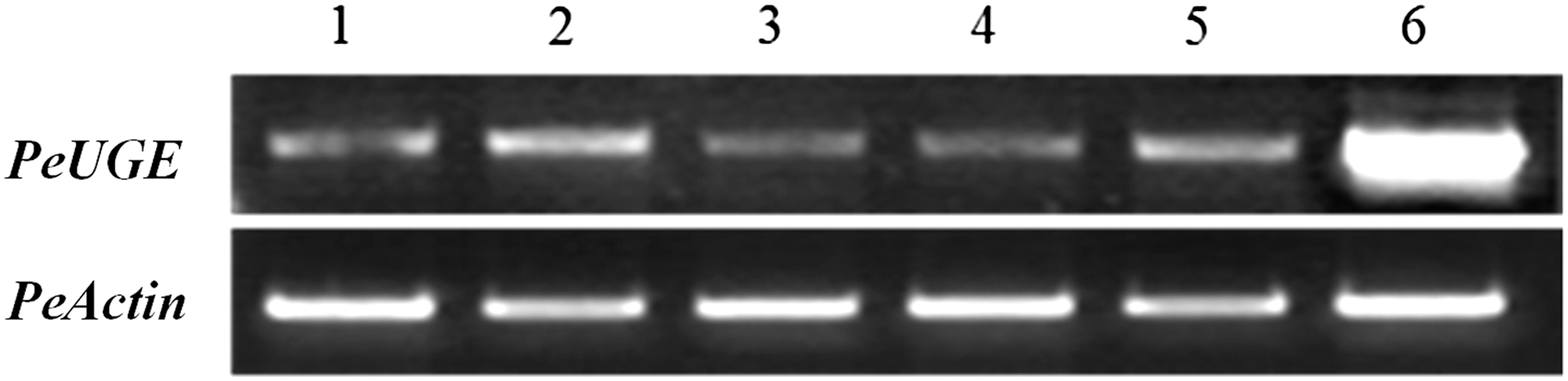
Tissue-specific expression analysis of PeUGE in P. edulis using semiquantitative RT-PCR. 1, Roots; 2, Stems; 3, Mature leaves; 4, Immature leaves; 5, Leaf sheaths; 6, Shoots. The amount of transcript was checked after 26 cycles by RT-PCR. RT-PCR, reverse transcription polymerase chain reaction.
As we all know, shoot is the immature stage of culm, which had no branches and no leaves, only culm sheaths wrapped around the shoot. To test whether the expression of PeUGE related to the development of shoots, we analyzed its transcript levels at different stages of shoots in the rapidly growing period using qRT-PCR with two different internal reference genes (NTB and TIP41) (Fig. 3). The results demonstrated that the expression level of PeUGE was upregulated with the growth of shoots continuously, which was up to about 30 times in 30 cm shoots compared to that in 2 cm shoots. This result confirmed that PeUGE might be involved in energy metabolism because a lot of energy, especially for the nonstructural carbohydrates such as soluble sugars and starch (Song et al., 2016), was required in the fast growth stage of bamboo shoots.
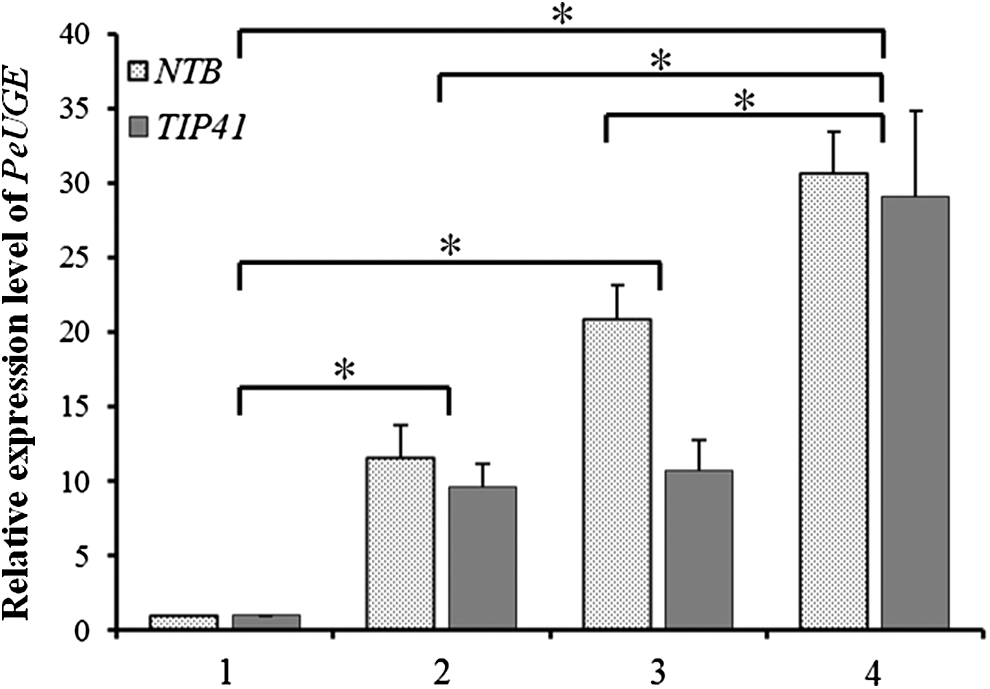
Transcript analysis of PeUGE in shoots at different development stages during rapid growth period. 1, 2 cm shoots; 2, 5 cm shoots; 3, 15 cm shoots; 4, 30 cm shoots. Vertical bars represent standard deviation (SD) of three independent biological replicates. The transcript of PeUGE demonstrated an increasing trend with the growth of shoots, in which 5 cm, 15 cm, and 30 cm shoots were higher than those 2 cm shoots (Asterisks indicate a significant difference at p < 0.01). Both NTB and TIP41 were used as reference genes.
Abiotic stresses are known as main factors affecting the growth and development of plants. To investigate the response of PeUGE to abiotic stresses, the expression patterns of PeUGE in roots were further analyzed after treatments of water, NaCl, and drought stresses using qRT-PCR with both NTB and TIP41 as reference genes, respectively. The results demonstrated that PeUGE was significantly induced by both water and NaCl stresses, especially under water stress it was upregulated for more than 40 times of the control (Fig. 4a). Under drought stress, the expression level of PeUGE was increased significantly with the extension of the treatment time, which was upregulated for more than 200 times during 6 h of the control (Fig. 4b). This result implied that PeUGE was involved in the response to osmotic stresses.
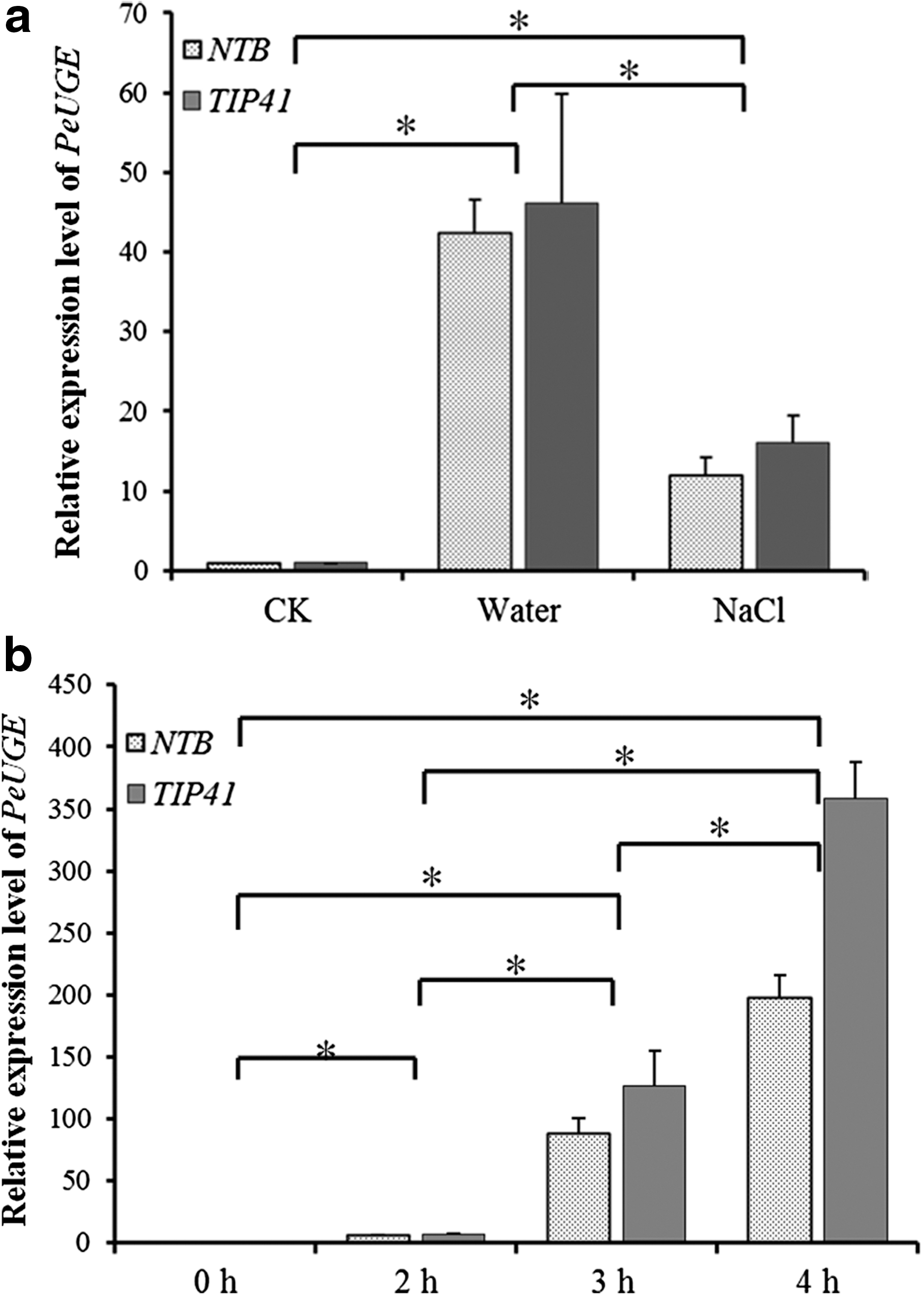
Transcript analysis of PeUGE gene in roots under stresses.
Overexpressing PeUGE in A. thaliana
To verify whether PeUGE was involved in osmotic stress tolerance, an expression vector containing the ORF of PeUGE driven by CaMV 35S promoter was constructed and transformed into A. thaliana. Six lines overexpressing PeUGE were obtained through resistance screening. Transcriptional expression of PeUGE was detected in two random transgenic lines, but not in Col-0 plants (Supplementary Fig. S3). Two T3 transgenic lines (L5 and L6) showing no evidence of segregation were focused on for further analysis. Both the results of qRT-PCR and RT-PCR indicated that PeUGE was transcribed abundantly in transgenic plants, which was 1.47 times in L6 compared to L5, but it was not detected in Col-0 plants (Supplementary Fig. S3). The chlorophyll fluorescence kinetic parameters can be used as a quick, nonintrusive probe in studies of plants under stress. For example, the maximal photochemical efficiency (F v/F m) is an index to estimate the degree of photoinhibition (Maxwell and Johnson, 2000). Under normal conditions, there was no difference in the F v/F m values of the PeUGE-overexpressing plants (L5) and Col-0 plants, and the phenotype of L5 was also similar to that of Col-0. Even though under the treatments of 10% PGE-6000 and 200 mM NaCl, there was still no significant difference in the phenotypes of L5 and Col-0 plants, which was consistent with that of overexpressing OsUGE-1 in A. thaliana (Liu et al., 2007). However, the values of both F v/F m and NPQ of L5 plants were higher than those of Col-0 plants under NaCl (Fig. 5a) and PGE-6000 stresses (Fig. 5b). Real-time PCR analysis revealed that the severity of the chlorophyll fluorescence parameters correlated well with the relative expression level of PeUGE (Supplementary Fig. S3). The differences of F v/F m and NPQ between L5 and Col-0 plants had not reached significant levels. The similar values of F v/F m and NPQ were also observed in the plants of L6 (data not shown).
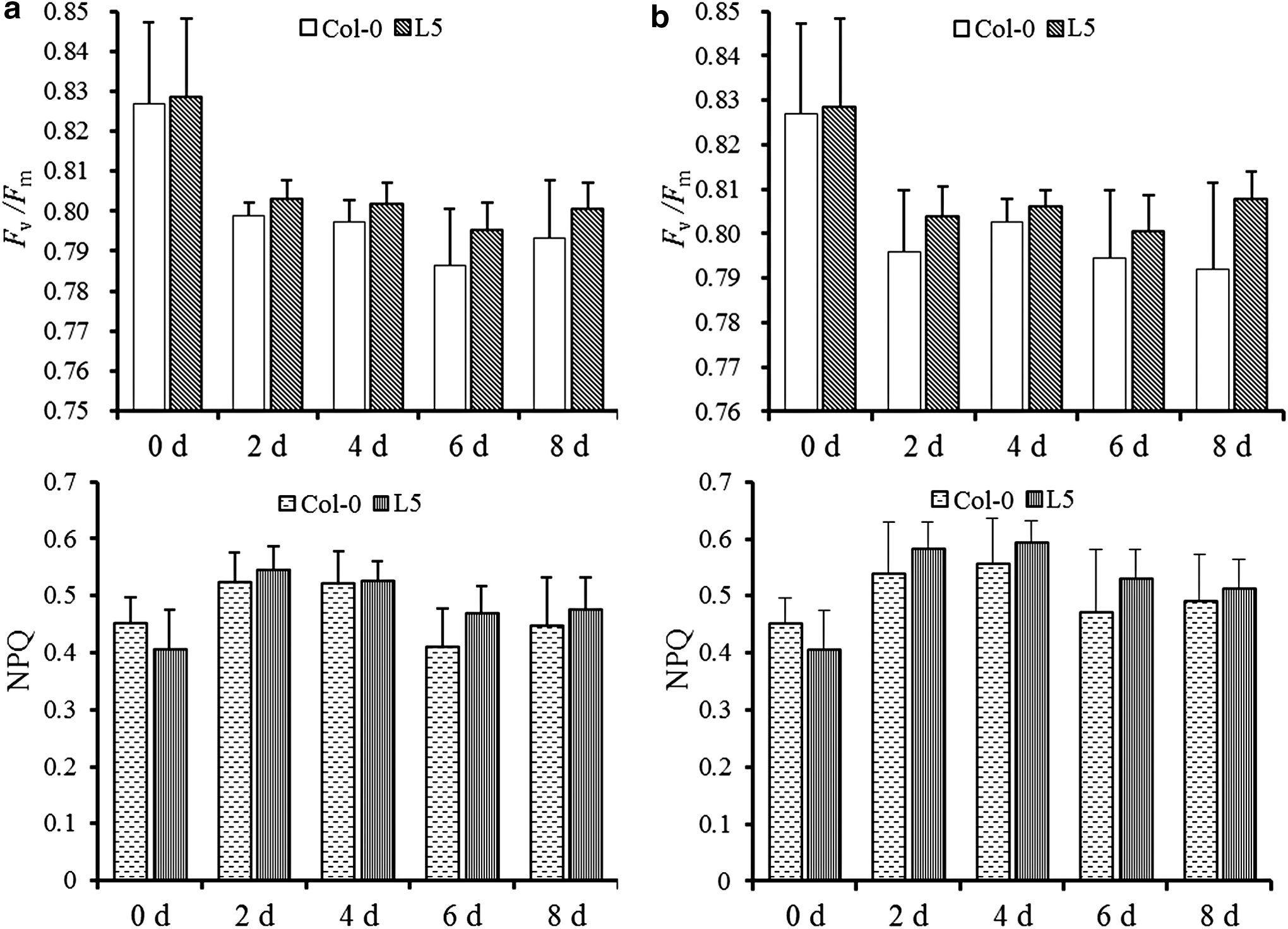
Response of Col-0 and transgenic plants to salt and drought treatments. Col-0 and L5 represented wild-type and overexpressing PeUGE plant line5, respectively. Plants grown in normal conditions were served as the control. Vertical bars represent SD of the mean (n = 30).
Root morphology of transgenic plants
To further test whether overexpressed PeUGE could confer abiotic stress tolerance in plants, the PeUGE-overexpressing seedlings in the MS medium supplemented with 100 mM mannitol or 50 mM NaCl in vertical plate culture were investigated. Under normal conditions, there was no phenotype difference between the transgenic and Col-0 plants (Fig. 6a). After treatments of NaCl and mannitol, there was still no significant phenotype difference in comparison of transgenic and Col-0 plants, but the unit lateral root numbers. It showed that the unit lateral root numbers of transgenic plants increased at least twice compared to that of Col-0 plants after both NaCl (Fig. 6b) and drought (Fig. 6c) treatments (p < 0.05). These results indicated that overexpression of PeUGE could enhance the tolerance of transgenic plants in response to salinity and drought stresses by increasing the number of lateral roots.

The phenotype and unit lateral root number analysis of transgenic plants and Col-0 under different treatments. Seedlings were photographed 14 days after germination. Lateral root numbers were counted with a binocular microscope directly. Numbers of lateral roots per centimeter of primary root in each seedling for two independent transgenic lines (L5 and L6) and Col-0 were presented at bottom (n = 18). Scale bar = 1 cm.
Discussion
With the rapid development of molecular biology and gene-engineering technology, an increasing number of UGEs have been successfully cloned from plants, such as potato (Oomen et al., 2004), A. thaliana (Dörmann and Benning, 1996), pea (Lake et al., 1998), rice (Rabbani et al., 2003), and barley (Zhang et al., 2006). On the basis of the full-length cDNAs of moso bamboo (Peng et al., 2010), there was a predicted UGE homologue gene (FP093917), which was 2146 bp with two terminating codons (TGA) presented in the predicted coding region. To understand the UGE in bamboo, we isolated the full-length cDNA of PeUGE and made further analysis. Similar to UGE in A. thaliana and rice, PeUGE was separated by nine introns (Supplementary Fig. S1), AG and GT in the acceptor and donor splice sites, respectively (Liu et al., 2007). The promoter regions in the genomic sequence of PeUGE contain numbers of stress responsive elements and meristem expression elements (Supplementary Table S1), all of which means PeUGE might be a key gene boosting stress relief and involved in cell wall synthesis, so further analysis on the function of PeUGE was also in need.
In potato tubers, overexpression of StUGE4 could increase cell wall galactose content and improve the toxic tolerance of galactose (Oomen et al., 2004). OsUGE1 in rice was implicated in cell wall carbohydrate metabolism by the repression of the cellulose synthesis-associated genes during limiting nitrogen conditions (Guevara et al., 2014). UGE2 and UGE4 in A. thaliana have been reported to involve in cell wall carbohydrate biosynthesis and shoot growth (Liu et al., 2007). Moso bamboo is one of the most fast-growing plants in the world, which means that cell division and cell elongation should simultaneously and rapidly dominate the development of culm. Study on the internode elongation-associated protein expression profiling in rapidly growing bamboo culms indicated that energy was mainly derived from sucrose degradation during culm development, as photosynthetic capacity was poor (Cui et al., 2012). We found that UGE was mainly expressed in the shoot (immature stage of culm), and the expression of PeUGE was continuously increased with the growth of the shoot. Moreover, lots of meristem expression elements found in the promoter regions also supported the prediction of its crucial roles in cell wall synthesis. All these results implied that PeUGE may be a key gene involved in biosynthesis of cell wall polysaccharides, which still need to be further validated.
Abiotic stresses such as drought, water, and salinity caused serious losses in plant production throughout the world (Boyer, 1982). It had been reported that UGE was induced by drought, cold, and high-salinity stresses in A. thaliana (Seki et al., 2002). The expression level of OsUGE1 in rice was upregulated by cold, drought, high salinity, and ABA treatments (Rabbani et al., 2003), and the gene plays a potential role in abiotic stress tolerance by maintaining proportionally more galactose and glucose, especially during N-deprivation (Guevara et al., 2014). Some studies show that overexpression of UGE can enhance the tolerance of stress, such as PvUGE1 in Paspalum vaginatum can enhance the salinity tolerance in rice (Endo et al., 2005), overexpressing rice UGE can increase the content of raffinose in transgenic A. thaliana, boosting stress relief, without altering plant morphology (Liu et al., 2007). In this study, the expression patterns of PeUGE and the phenotype of its transgenic plants were similar to those of the existing research reports on UGEs.
Although lot of studies have reported that UGE can enhance the stress tolerance of plants, up to date, few studies have investigated the chlorophyll fluorescence parameters and the development of the lateral roots of the transgenic plants under stress. NPQ is a tool to describe the NPQ processes, and the F v/F m is an index to estimate the degree of photoinhibition, the two parameters are indices to mirror the degree of stress (Maxwell and Johnson, 2000). Under normal growth conditions, there was no difference in the lateral root numbers and the two parameters between the transgenic and Col-0 plants. After treatment of PEG-6000 or NaCl, the lateral root number of transgenic plants increased, meanwhile both the F v/F m and NPQ values of transgenic plants were higher than those of Col-0 plants. All the results indicated that overexpressing PeUGE conferred increased tolerance to salinity and drought stresses in transgenic Arabidopsis plants by increasing the number of lateral roots. Nonetheless, further research is needed to help us gain a better understanding about the function of PeUGE in bamboo.
Footnotes
Acknowledgments
The work was supported by the Special Fund for Forest Scientific Research in the Public Welfare from State Forestry Administration of China (No. 201504106), the National Sci-Tech Support Plan (No. 2015BAD04B01 and No. 2015BAD04B03) and Fundamental Research Funds for International Center for Bamboo and Rattan (No. 1632015008).
Disclosure Statement
The authors declare no competing financial interests. All the authors agreed to the submission of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.