
Abstract
Osteoarthritis (OA) is one of the most common prevalent chronic joint diseases. Emerging pieces of evidence have demonstrated that chondrocytes survival was closely associated with the destruction of joints in OA patients. Long noncoding RNAs (lncRNAs), defined as >200 nucleotides in length, also have been implicated in a variety of disease states. However, there are few studies on the role of lncRNAs in OA, and the pathological contributions of lncRNAs to OA remain largely unknown. In this study, we examined the expression of lncRNA UFC1 in cartilage samples from OA patients and healthy subjects, and then investigated biological function of UFC1 in OA chondrocyte. We found that the UFC1 was significantly reduced in OA patients. Functional assays demonstrated that UFC1 promotes chondrocytes proliferation and inhibits cell apoptosis. Furthermore, we found that UFC1 regulates survival of OA chondrocytes through physically association with miR-34a. Taken together, our data highlight the important roles of lncRNA UFC1 in the survival of OA chondrocytes. UFC1 may be a potential therapy for OA.
Introduction
O
The human transcriptome comprises not only protein-coding messenger RNAs (mRNAs) but also a large number of transcripts without protein-coding potential (Riquelme et al., 2016). These noncoding RNAs (ncRNAs) have been shown to take part in multiple biological processes (Fitzgerald et al., 2016; Sandhu et al., 2016). Dysregulation of ncRNA is closely associated with diverse diseases including OA (Marques-Rocha et al., 2015; Tornero-Esteban et al., 2015). MicroRNAs are small noncoding RNAs with the length of about 20–25 nucleotides (nt), which have regulatory function on gene expression in a post-transcriptional manner and contribute to the pathogenesis of OA (Li et al., 2016). For example, silencing of miR-34a could effectively reduce chondrocyte apoptosis induced by IL-1β (Abouheif et al., 2010). Recently, long noncoding RNAs (lncRNAs), defined as >200 nt in length, also have been implicated in a variety of disease states (Schmitz et al., 2016; Xin et al., 2016). However, there are few studies on the roles of lncRNAs in OA, and the pathological contributions of lncRNAs to OA remain largely unknown.
Long intergenic noncoding RNA UFC1 (lincRNA-UFC1) is a new identified functional lncRNA. UFC1 promotes proliferation and reduces apoptosis in hepatocellular carcinoma cells through interacting with the mRNA stabilizing protein HuR to regulate levels of β-catenin (Cao et al., 2015). In this study, we examined the expression of UFC1 in cartilage samples from OA patients and healthy subjects, and then investigated the role of UFC1 in OA chondrocyte.
Materials and Methods
Articular cartilage tissues
The OA cartilage was isolated from the knee joints of 30 patients undergoing total knee arthroplasty. OA was confirmed using radiographic images and diagnosed according to the American College of Rheumatology criteria. The normal articular cartilage was isolated from the knee joints of 10 patients with femoral neck fracture without OA or rheumatic arthritis who underwent total hip replacement surgery. All 45 tissue donors included in this study provided informed consent. The study was approved by the Human Ethics Committee of Qianfoshan Hospital of Shandong University.
Isolation and culture of human chondrocyte
The articular cartilage samples were minced and followed by digestion at 37°C with 0.2% collagenase II (Invitrogen) in Dulbecco's modified Eagle's medium (DMEM) for 5–6 h with stirring every 30 min after 2 h. Chondrocytes were isolated after centrifugation. Cells were maintained in DMEM containing 10% fetal bovine serum for 5–7 days before use, and incubated at 37°C in a humidified atmosphere of 5% CO2.
Quantitative real-time polymerase chain reaction
Total RNAs from articular cartilage samples or cultured chondrocytes were isolated by Trizol reagent (Invitrogen) and reverse transcribed. SYBR Green Master Mixture (Roche) was used for GAPDH and UFC1 expression. GAPDH mRNA was employed as an endogenous control. The polymerase chain reaction (PCR) primers specific for UFC1 were 5′-TCCAACCTGAGTGACATAGCGA-3′ (sense) and 5′-CTGACCTCCAACTCCAACGAAT-3′ (antisense). The relative expression of RNA was calculated using the comparative Ct method.
RNA immunoprecipitation
Cells were cotransfected with pcDNA3.1-MS2, pcDNA3.1-MS2-UFC1, or pcDNA3.1-MS2-UFC1-mut and pMS2-GFP (purchased from addgene) by using Lipofectamine 3000 according to the manufacturer's protocol (Invitrogen). After 48 h, cells were subjected to perform RNA immunoprecipitation (RIP) assays using a GFP antibody (Abcam) and the EZ-Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to the manufacturer's protocol.
For anti-AGO2 RIP, cells were transfected with miR-34a mimics or microRNA negative control (miR-NC). After 48 h, cells were subjected to perform RIP assays using an AGO2 antibody (Millipore) as already described.
Fluorescence-activated cell sorting
Cells (3 × 105) were trypsinized and resuspended to generate single-cell suspensions. Cells were stained with fluorescein isothiocyanate-conjugated Annexin V and 7-AAD (KeyGEN) according to the manufacturer and analyzed by using a flow cytometer. The data were studied using FlowJo software.
Luciferase reporter assay
A wildtype or mutated full-length UFC1 was cloned into pmirGLO reporter plasmid. The wildtype or mutated reporter vectors and miR-34a mimics were cotransfected into cells. After 48 h, luciferase activity was measured by a dual-luciferase reporter assay system (Promega).
siRNA transfection
The siRNAs against UFC1 were purchased from Riobo Company. siRNAs were transfected to cells by Lipofectamine 3000 according to the manufacturer's protocol (Invitrogen).
Data analysis
Data shown are the means ± standard deviation. Statistical analyses for data were performed by using SPSS 19.0 software. Data were analyzed between two groups using the Student's t-test. p < 0.05 was considered as significant difference.
Results
UFC1 expression was significantly reduced in OA patients
To examine the expression level of UFC1 in the articular cartilage, quantitative real-time (qRT)-PCR was performed in samples from 30 OA patients and 20 healthy subjects. We found that the expression of UFC1 was significantly downregulated in the OA group compared with the healthy group (Fig. 1).
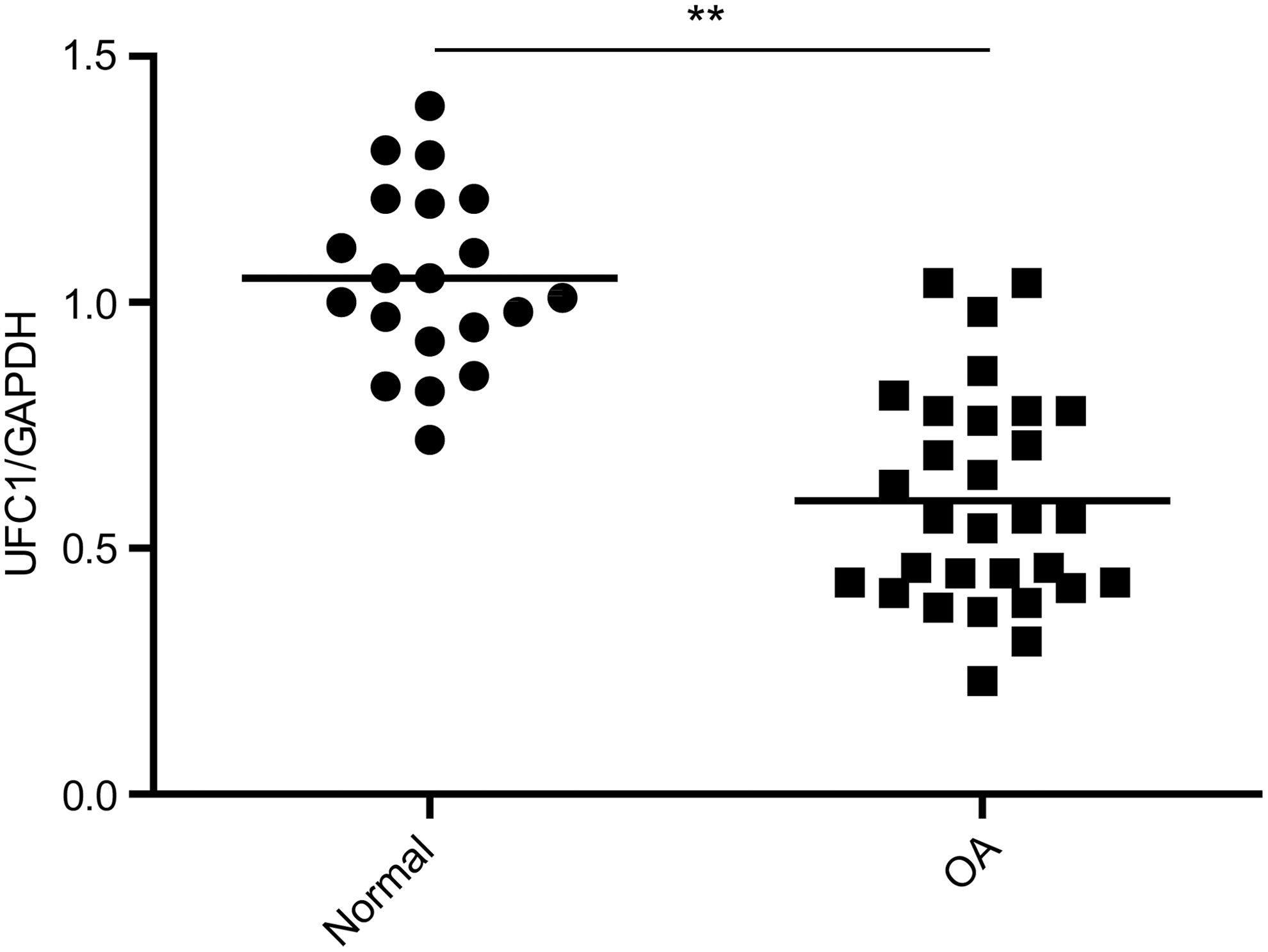
UFC1 expression was significantly reduced in OA patients. The expression of UFC1 in articular cartilage samples from normal objects and OA patients was detected by qRT-PCR. Data are shown as mean ± std. **p < 0.01. OA, osteoarthritis; qRT-PCR, quantitative real-time polymerase chain reaction; std, standard deviation.
UFC1 promotes survival of human OA chondrocytes
To determine the role of UFC1 in OA chondrocytes, we performed gain- and loss-of-function studies. OA chondrocytes were transiently transfected with full-length UFC1 or siRNA against UFC1 (Fig. 2A). By performing CCK-8 assay, we found that overexpression of UFC1 promoted the proliferation of chondrocytes, whereas knockdown of UFC1 suppressed the proliferation of chondrocytes (Fig. 2B). Furthermore, we analyzed differences in cell apoptosis after UFC1 overexpression or knockdown by fluorescence-activated cell sorting. The results showed that overexpression of UFC1 inhibited apoptosis in chondrocytes (Fig. 2C). In contrast, UFC1 knockdown chondrocytes had a significantly higher percentage of Annexin V-positive cells than control cells (Fig. 2C).
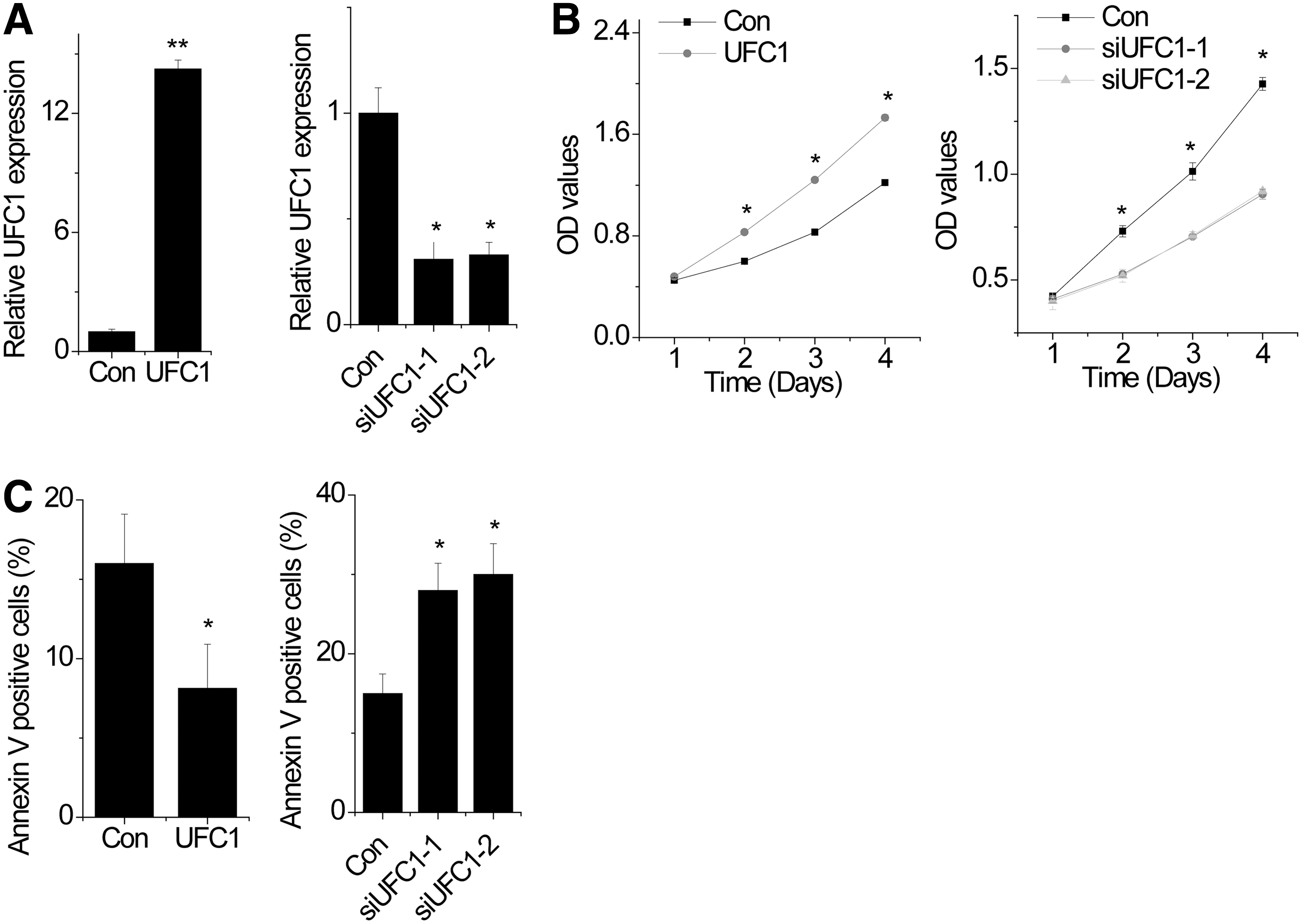
UFC1 promotes survival of human OA chondrocytes.
UFC1 is physically associated with miR-34a in OA chondrocytes
Previous studies have reported that miR-34a could affect the apoptosis of chondrocytes and regulate UFC1 expression (Abouheif et al., 2010; Cao et al., 2015). However, the exact mechanism by which miR-34a modulates UFC1 expression in OA chondrocytes is still unknown. We suspected that UFC1 regulated proliferation and apoptosis through interaction with miR-34a. To validate the direct binding between miR-34a and UFC1, we performed an RIP to pull down endogenous microRNAs associated with UFC1 (Fig. 3A). We found that the UFC1 RIP in chondrocytes was significantly enriched for miR-34a compared with the empty vector (MS2), UFC1 with mutations in miR-34a targeting sites (henceforth named UFC1-mut) (Fig. 3B). In addition, we constructed luciferase reporters containing wildtype UFC1 or mutated miR-34a binding site. We found that overexpression of miR-34a reduced the luciferase activities of the wildtype reporter vector but not empty vector or mutant reporter vector (Fig. 3C). Finally, we found that UFC1 expression was reduced after overexpression of miR-34a (Fig. 3D).
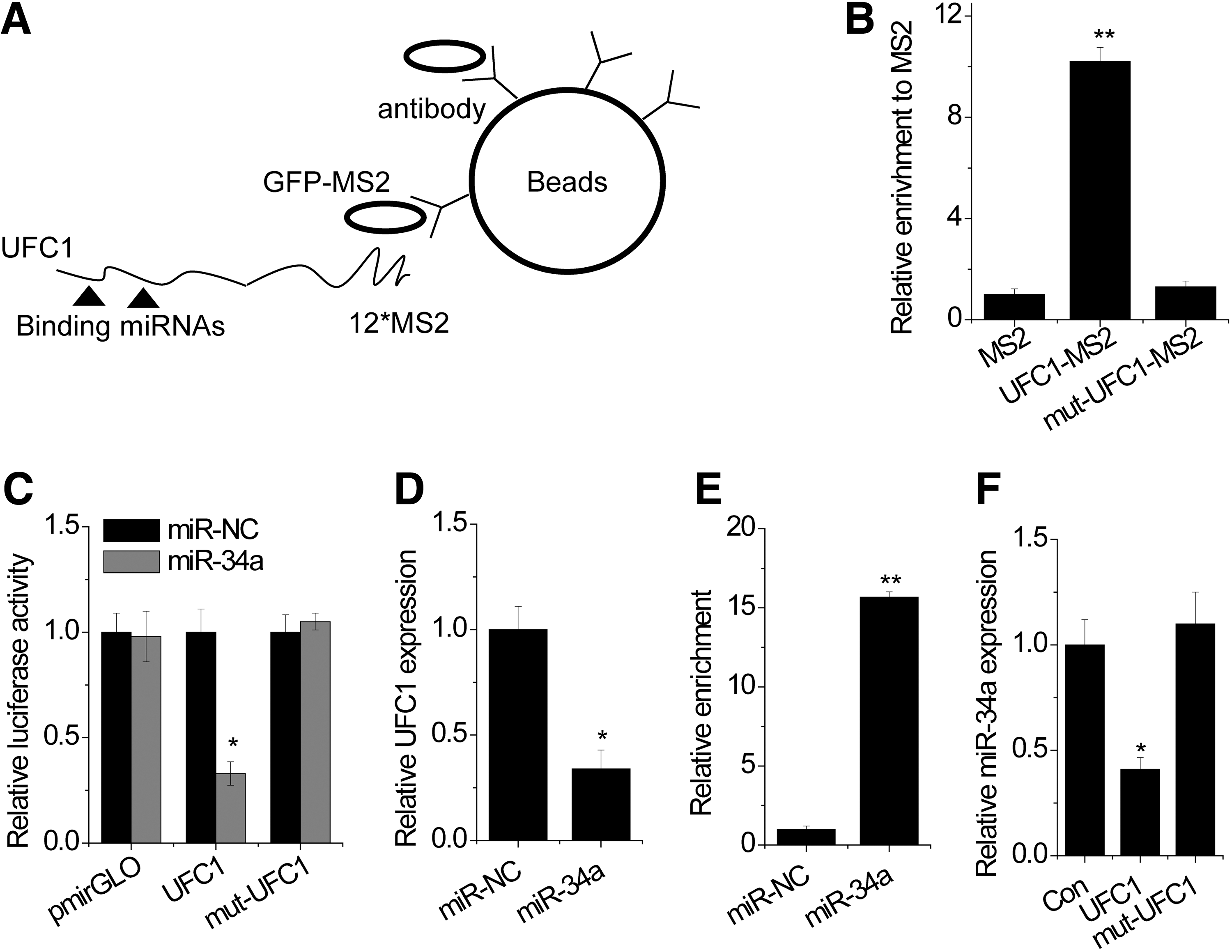
UFC1 is physically associated with miR-34a in OA chondrocytes.
The microRNAs are known to bind their target mRNAs and induce translational repression or degradation in an AGO2-dependent manner. To determine whether lncRNA UFC1 was regulated by miR-34a in this manner, we performed anti-AGO2 RIP in chondrocytes transiently overexpressing miR-34a. UFC1 pull-down by AGO2 was significantly enriched in miR-34a-overexpressed cells (Fig. 3E). In addition, overexpression of wildtype UFC1 but not the mutant suppressed the miR-34a level (Fig. 3F). All these data demonstrated that UFC1 physically associated with the miR-34a.
UFC1 promotes survival of OA chondrocytes through miR-34a
To investigate whether UFC1 regulates proliferation through miR-34a, rescue experiments were performed. UFC1 knockdown chondrocytes were transfected with miR-34a inhibitor. We found that miR-34a inhibitor abolished the phenotypes of cell proliferation and apoptosis induced by UFC1 knockdown (Fig. 4A, B). In contrast, overexpression of miR-34a rescued the promotion of proliferation and inhibition of apoptosis mediated by UFC1 overexpression (Fig. 4C, D). These results suggest that UFC1 promotes cell proliferation through miR-34a.
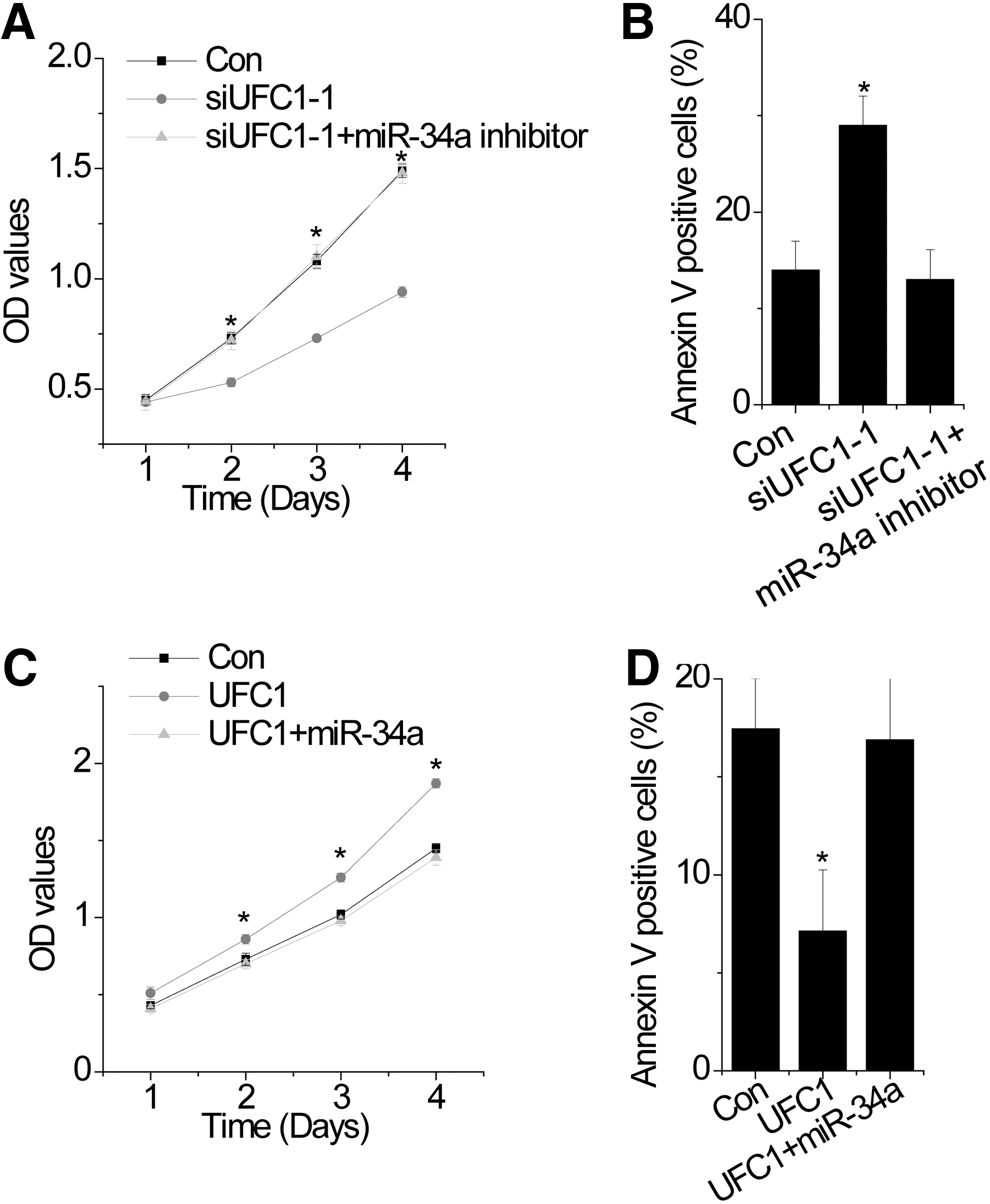
UFC1 promotes survival of OA chondrocytes through miR-34a.
Discussion
In this study, for the first time, we reported that the UFC1 was significantly reduced in OA patients, and UFC1 promotes chondrocytes proliferation and inhibits apoptosis by directly targeting miR-34a. Therefore, UFC1 was determined to have an inhibitory effect on OA. UFC1 may be a potential therapy for OA.
Our results indicated that a lower level of UFC1 in the articular cartilage was associated with OA patients. Recent studies have reported that many lncRNAs are dysregulated in OA. For example, human lncRNA MEG3 is significantly downregulated in OA patients compared with normal cartilage samples (Su et al., 2015). lncRNA GAS5 was upregulated in OA chondrocytes compared with non-OA and normal chondrocytes (Song et al., 2014). Moreover, a recent study has found a series of lncRNAs were dysregulated in OA patients by using lncRNAs microarray (Fu et al., 2015), which indicated that lncRNAs may be new biomarkers for diagnosis or novel therapeutic targets of OA. UFC1 is known to positively regulate cell survival and is aberrantly expressed in several cancers, such as colorectal cancer and liver cancer (Cao et al., 2015; Yu et al., 2016). In this study, we determined the expression level of UFC1 in OA cartilage samples. We found that UFC1 is significantly decreased in cartilage samples from OA patients compared with that from the healthy group. Functional studies demonstrated that UFC1 promotes proliferation and inhibits apoptosis in OA chondrocytes. It is known that increased apoptosis of chondrocytes is critical for pathogenesis of OA (Freitag et al., 2016). Restoring of UFC1 expression in OA chondrocytes may be a potential therapy for treatment of OA.
lncRNAs and microRNAs play an important role in transcriptional or post-transcriptional regulation, which are involved in many human diseases. The direct interactions between lncRNAs and microRNAs have aroused interest recently. For example, lncRNA-ATB serves as an endogenous microRNA sponge of miR-200 family to regulate ZEB1 and ZEB2 expression and tumor metastasis (Yuan et al., 2014), and miR-204 targets lncRNA HOTTIP to suppress HOTTIP expression (Ge et al., 2015). These discoveries trigger us to explore functional association between lncRNA UFC1 and microRNAs. Here, we present strong pieces of evidence that UFC1 is physically interacting with miR-34a in OA chondrocytes. Previous studies have shown that miR-34a was significantly upregulated by IL-1β, which was a main inflammatory factor that induced apoptosis of OA chondrocytes. And silencing of miR-34a could effectively reduce chondrocyte apoptosis induced by IL-1β (Abouheif et al., 2010). In this study, we found that UFC1 could interact with miR-34a and suppressed its expression. These indicated that UFC1 may be a protective molecule for IL-1β-induced apoptosis.
In summary, our study demonstrated that UFC1 facilitates proliferation and inhibits apoptosis in a miR-34a-dependent manner in OA chondrocytes. These findings indicate that UFC1 is a critical molecule for OA progression.
Footnotes
Disclosure Statement
No competing financial interests exist.