
Abstract
Lung cancer is one of the most malignant cancers with a high metastatic potential. The purpose of this study was to study the role and the underlying mechanism of miR-214 in lung cancer progression. The expression of miR-214 in normal lung and lung cancer tissue was analyzed by quantitative real-time PCR analysis. Furthermore, H1299 cells were infected with miR-214 lentivirus, and the effect of infection on cell viability and migration was analyzed. Carboxypeptidase-D (CPD), as a potential target of miR-214, was characterized in either normal lung or lung cancer tissues. The interaction of CPD expression with the tumor suppressing effect of miR-214 was characterized. We demonstrated that low miR-214 expression is a hallmark of lung cancer, especially high-grade and metastatic cancer. In vitro studies in H1299 cells confirmed that low miR-214 expression is associated with enhanced proliferation and migratory abilities. Similarly, CPD overexpression coincides with high-grade lung cancer and the CPD overexpression could reverse the inhibitory effects of miR-214. miR-214 is a tumor suppressor in lung cancer. miR-214 inhibits lung cancer progression by targeting CPD. The miR-214-CPD axis may be a therapeutic axis for lung cancer patients.
Introduction
L
Carboxypeptidase-D (CPD) is a membrane bound metalloprotein that cleaves C-terminal arginine for conversion to nitric oxide (NO) by nitric oxide synthase, through which a number of cellular processes, including tumor progression, are regulated (Jin et al., 2013; Thomas et al., 2015). Due to this reason, CPD gene has been regarded as an oncogene in cancers such as breast cancer (Abdelmagid and Too, 2008) and prostate cancer (Petrylak et al., 2004), and CPD overexpression is tightly correlated with oncogenic cues such as prolactin, estrogen, and testosterone inductions (Abdelmagid and Too, 2008; Thomas et al., 2012). However, the lack of studies of CPD in lung cancer warrants further investigation of CPD's role in lung cancer progression. In this study, we hypothesized that the interplay between miR-214 and CPD may play a significant role in lung cancer progression. Understanding how miR-214 and CPD work in concert in lung cancer progression may shed light on viable therapeutics to prevent lung cancer metastasis.
Therefore, the aim of this study is to evaluate the role of miR-214 in lung cancer progression and investigate on the correlation between miR-214 and CPD expression. We demonstrated that low miR-214 and high CPD expression are both characteristics of malignant and metastatic lung cancer. The shining point of our study is that we, for the first time, showed that miR-214 suppresses cancer progression by targeting CPD gene, and the interplay between miR-214 and CPD is crucial for lung cancer progression. Considering the urgent unmet need for a diagnostic and therapeutic method for lung cancer, miR-214 and CPD as biomarkers could potentially be applied in clinics for developing new strategies that improve lung cancer diagnostic and therapeutic efficiencies.
Materials and Methods
Cell culture and viability assay
H1299 non-small cell lung cancer cell lines, BEAS-2b, A549, and PC9 cells, were purchased from American Type Culture Collection (ATCC, Manassas, VA) and were maintained in RPMI medium containing 10% fetal bovine serum. Cells were seeded in 96-well plates before viability analysis. Cell Counting Kit-8 (CCK-8) solution (10 μL) was added into each well. After a 4-h incubation, absorption value at 450 nm was measured by a plate reader.
Migration assay
Scratch wound assay were performed by enforcing a 10 μL pipette tip on a confluent plate. After wound gaps were made, phase contrast images of the gaps were taken at a time interval of 4 h. Decrease in the gap area was quantified as ratio of initial gap area. Images were analyzed using ImageJ. Cell migration was also evaluated using a Transwell Matrigel invasion assay in 24-well Transwell units (Corning, New York, NY). Each well contained 105 H1299 cells suspended in RPMI. Seeded wells in the upper chamber were coated with Matrigel; 500 μL RPMI was applied in the lower chamber. After 24 h, invading cells in the bottom were fixed and analyzed by measuring absorbance at 570 nm.
Quantitative reverse-transcription PCR
Tissue for miR-214 and CPD characterization was collected from The First Hospital of Shijiazhuang. Total RNA from cells was extracted using TRIzol reagent (Invitrogen, Pleasanton, CA) and genomic DNA removed using the TURBO DNase Kit (Ambion) and quantified by NanoDrop. The cDNA was synthesized using PrimeScript RT Reagent Kit (Takara Bio, Kusatsu, Japan) following the manufacturer's instructions. This study was approved by the Ethics Committee of The First Hospital of Shijiazhuang, and informed consents were obtained.
Western blot
For western blot analysis, tissue samples were homogenized using a Dounce homogenizer in 1 mL of lysis buffer, containing 15 mM Tris pH 7.6, 0.25 M sucrose, 1 mM MgCl2, 1 mM EGTA, 1 mM DTT, 1.25 μg/mL pepstatin A, 10 μg/mL leupeptin, 25 μg/mL aprotinin, 0.5 mM PMSF, 0.1 mM Na3VO4, 50 mM NaF, 2 mM Na4P2O7, and 1× phosphatase inhibitor cocktail set II. Protein concentrations in tissue lysates were measured with BCA assay using Pierce BCA Protein Assay Kit (Thermo Fisher, Waltham, MA). Thirty micrograms of proteins were loaded. Electrophoresis was performed using Mini-PROTEAN Tetra Cell system using precast gels and blotted with PVDF membranes.
Lentivirus packaging
To stably express miR-214 and CPD in H1299 cells, the lentiviruses carrying miR-214, negative control (miR-NC), or CPD cDNA were packaged using the Lentiviral Packaging Kit (Thermo Fisher) following the manufacturer's manual. Lentivirus was packaged in H1299 cells and secreted into the medium. H1299 cells were infected by lentivirus carrying miR-214, miR-NC, or CPD cDNA with the presence of Polybrene (Sigma-Aldrich) and selected by Puromycin for 2 weeks to obtain stable cell lines.
Dual-luciferase reporter assay
Wild-type and mutant CPD genes were subclined into pGL3 Basic luciferase reporter plasmid (Promega, Madison, WI). FuGENE 6 (Boehringer, Shanghai, China) was used for transfections. Reporter gene assays were performed with the Dual-Luciferase Reporter Assay System (Promega). Fifty nanograms pRL-TK (Promega) renilla luciferase was cotransfected in each sample as an internal control for transfection efficiency.
Statistical analysis
One- or two-way analysis of variance followed by a Tukey's post hoc test was performed to analyze the significance of differences. Spearman's correlation was used to analyze the relationship between two factors. Differences were considered statistically significant at p < 0.05.
Results
Low miR-214 expression is correlated with lung cancer malignancy
First, to verify that low miR-214 expression can serve as a biomarker for lung cancer, quantitative PCR analysis of miR-213 expression in normal and lung cancer tissues was performed. As indicated in Figure 1A, lung cancer tissues (n = 52) demonstrated a significantly lower miR-214 level compared to that in normal tissue (n = 52). More importantly, the level of miR-214 expression seems to be inversely correlated with the stage of cancer (Fig. 1B). Lymph nodes with metastases also showed lower miR-214 expression (Fig. 1C). In addition, the levels of miR-214 expression in different lung cancer cell lines, including A549, PC9, and H1299 cells, were significantly lower than normal lung/brunch epithelial cell line (BEAS-2b) (Fig. 1D).

miR-214 levels are downregulated in human lung cancer tissues and correlated to metastasis.
Inhibitory effect of miR-214 overexpression on lung cancer proliferation and metastasis
Based on these findings, we further investigated the role of miR-214 in cell proliferation and migration. As shown in Figure 2A, infection of H1299 cells with miR-214 lentivirus resulted in much significantly higher miR-214 density than H1299 cells infected with miR-NC (negative control). As a result, a reduction in cell viability was shown in H1299 cells infected with miR-214 (Fig. 2B). Wound healing assay and Matrigel invasion assay also indicated a compromised migratory ability of miR-214 expression in H1299 cells (Fig. 2C and D). Similar results could be found in another lung cell line (PC9 cells), in which miR-214 overexpression was able to inhibit cell proliferation and metastasis (Supplementary Fig. S1; Supplementary Data are available online at

miR-214 overexpression inhibited cell proliferation and metastasis of lung cancer.
miR-214 inhibits lung cancer cell by targeting CPD
To account for the inhibitory function of miR-214 in lung cancer, we hypothesized that CPD is a target gene of miR-214. Indeed, seed matching sites between miR-214 and 3′ UTR of CPD (Fig. 3A). In contrast, CPD mutant gene (CPD-MUT) possesses mutant sites that disrupt the interaction between CPD-MUT and miR-214. This hypothesis was verified by luciferase reporter assay on wild type of mutant cells, in which wild-type cells displayed a significant decrease in luciferase activity upon miR-214 infection, indicating that miR-214 bound to CPD, whereas CPD mutant cells didn't respond to miR-214 infection (Fig. 3B). Opposite to the tumor suppressing function of miR-214, CPD overexpression can promote cancer progression. We examined CPD mRNA level in paired normal and lung cancer tissue and found that lung cancer tissue showed a significantly higher CPD expression (Fig. 3C). The increase in CPD expression was also positively correlated with the grade of cancer (Fig. 3D). CPD was also found to be highly expressed in lymph node with metastases (Fig. 3E). Clearly, a negative correlation can be found between miR-214 expression and CPD expression, as indicated by Spearman's correlation analysis (Fig. 3F), and infection of miR-214 lentivirus decreased the CPD expression in protein level (Fig. 3G).

miR-214 directly targets and inhibits CPD expression, which correlated to cancer metastasis.
Silence of CPD suppresses lung cancer cell viability, migration, and invasion
To further validate the role of CPD in cell proliferation and migration, we suppressed CPD expression in H1299 cells using a CPD small hairpin RNA (shRNA), shCPD, which induced reduction of CPD mRNA level to about 27% of the original level (Fig. 4A). As a result, reduced cell viability and migratory ability were also seen in cells transfected with shCPD (Fig. 4B–D).
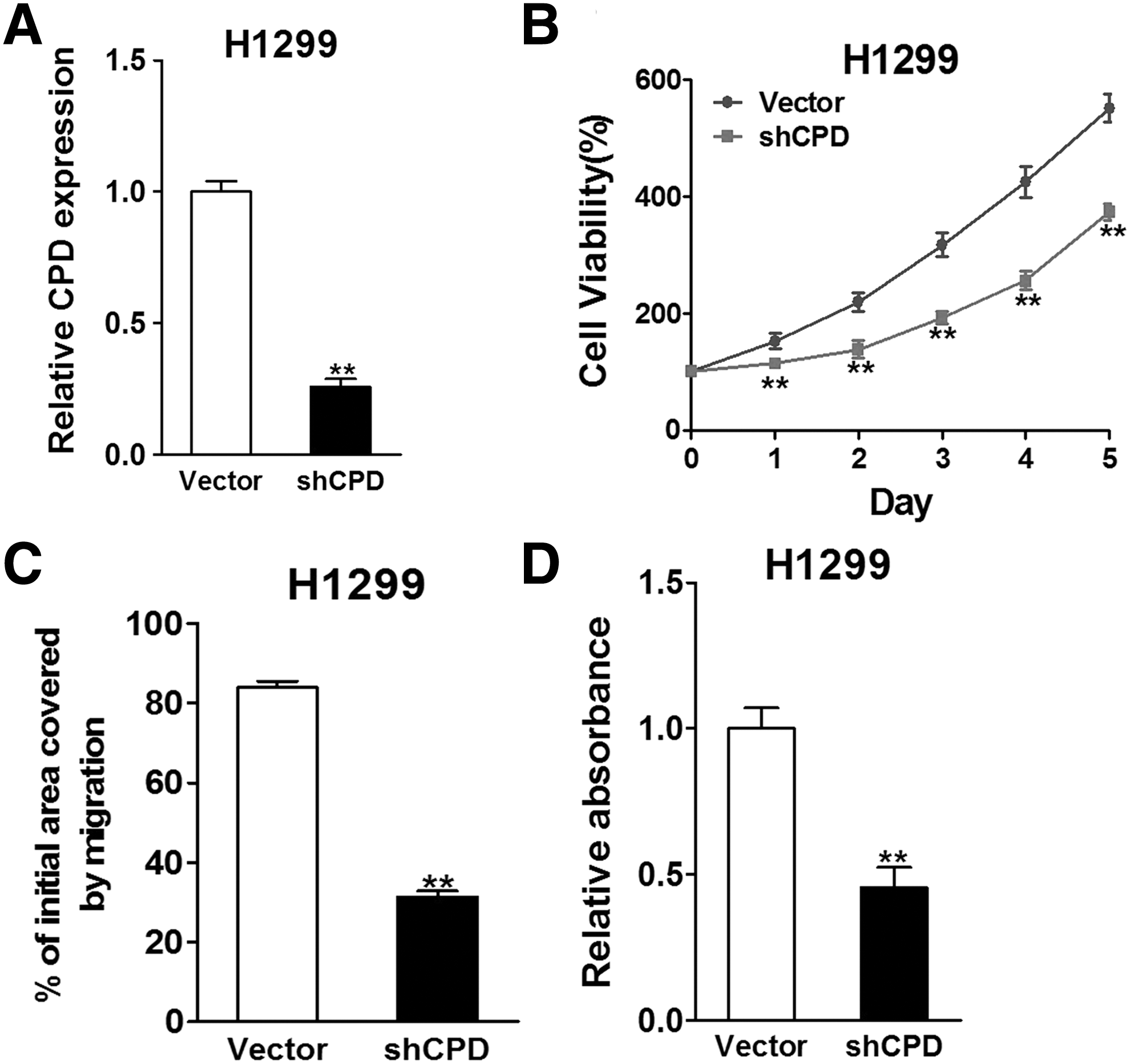
Silence of CPD suppresses lung cancer cell viability, migration, and invasion. H1299 cells were transfected with vector or shCPD.
Overexpression of CPD reverses the inhibitory effects of miR-214
Furthermore, we showed that the inhibitory effect of miR-214 can be reversed by CPD overexpression. Transfection of CPD resulted in an upregulated expression of CPD, even with the presence of miR-214, as indicated by western blotting analysis (Fig. 5A). In cell viability assay, even though cells infected with miR-214 lentivirus (miR-214+vector) showed a decreased cell viability, overexpression of CPD in these cells (miR-214+CPD) reversed the cell inhibitory effect of miR-214, increasing the viability to about 79% of the original level (Fig. 5B). Unsurprisingly, cell migration analysis also indicated an increased migrating ability in H1299 cells received both by miR-214 lentivirus and CPD overexpression (Fig. 5C and D).
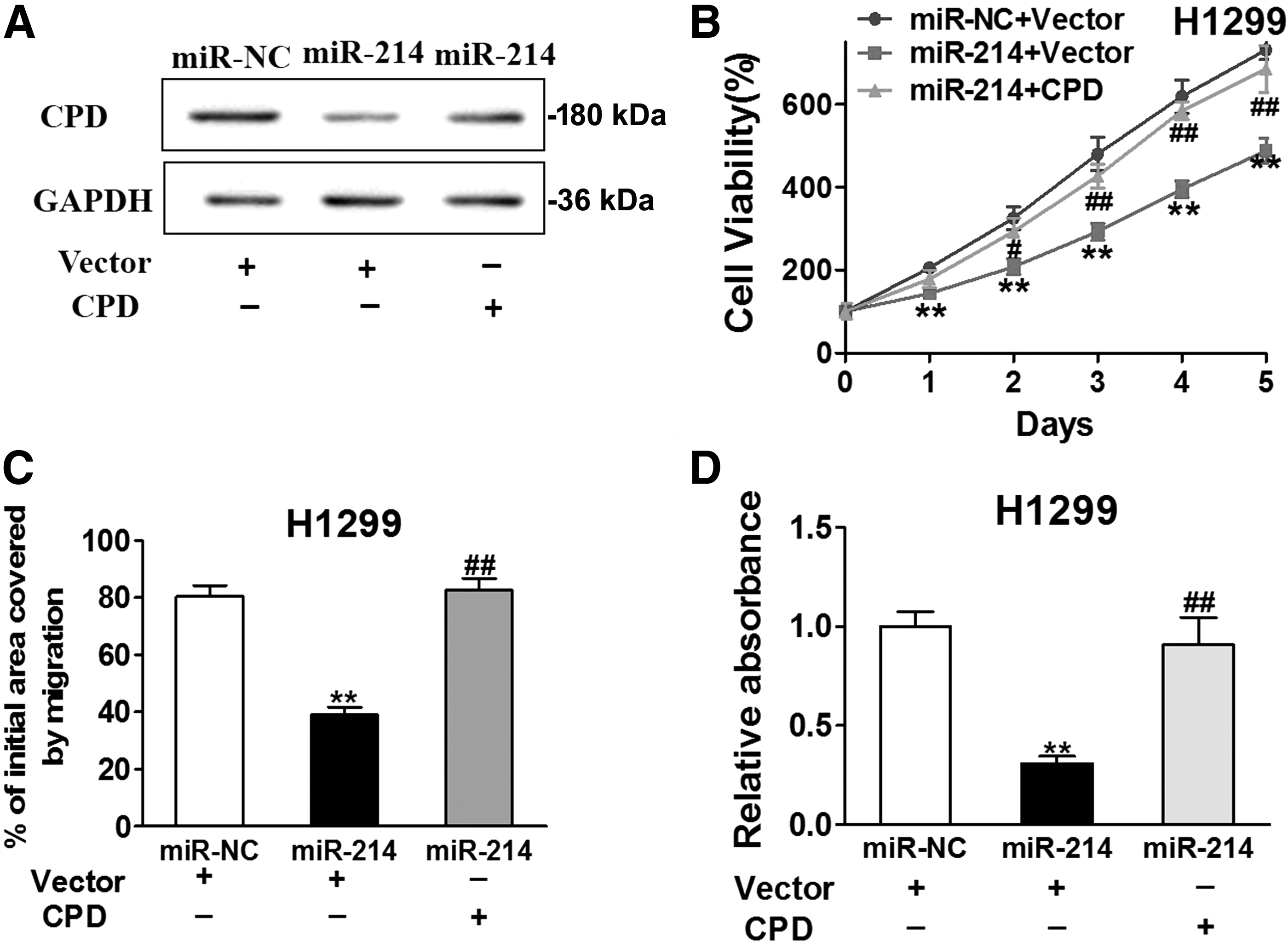
Overexpression of CPD reverses the inhibitory effects of miR-214. miR-214- or miR-NC-overexpression cells were transfected with vector or CPD.
Discussions
Lung cancer is the most common cause of cancer-related death among both men and women, killing almost combined tolls of breast cancer, prostate cancer, and colorectal cancer (Keshamouni et al., 2009). The high metastatic potential of lung cancer is responsible for its severe mortality. Innovative, noninvasive, and sensitive diagnostic and therapeutic strategies entail development of new biomarkers of lung cancer. With our understanding of miRNA expression patterns and function in normal and neoplastic cells starting to emerge, a new wave of studies has been focused on characterizing the clinical relevance of these miRNAs in regulating cancers. In lung cancer, miRNA profiles associated with cancer malignancy have been reported (Yanaihara et al., 2006), which paved way for further research for developing novel diagnostic markers and therapeutic strategies. miR-214 was found to be dysregulated in a variety of cancers, including pancreatic cancer (Zhang et al., 2010), breast cancer (Schwarzenbach et al., 2012), gastric cancer (Ueda et al., 2010), ovarian cancer (Yang et al., 2008), etc, in which miR-214 serves as a tumor suppressor and was negatively correlated with cancer malignancy and cancer resistance. In lung cancer, low miR-214 expression has been documented in malignant cancers (Miko et al., 2009). However, no clinical validation of miR-214 expression in human lung cancer tissues has been reported. More importantly, the molecular mechanism of miR-214 and cancer progression remains unclear. In this study, we performed quantitative real time polymerase chain reaction analysis of miR-214 expression on a large array of human normal lung tissues and lung cancer tissues. We demonstrated that low miR-214 expression was closely associated with the presence of lung cancer and, more importantly, high-grade lung cancer and metastatic lung cancer possessed displayed even lower miR-214 expression. This further supports that low miR-214 expression is a clinically significant biomarker for diagnosing lung cancer.
Having shown the clinical relevance of miR-214 expression and cancer, we next examined the effect of miR-214 on cell proliferation and migration. In our study, infection of human H1299 cancer cells with miR-214 lentivirus resulted in upregulation of miR-214, which unsurprisingly led to decreased cell proliferation and migration. This finding mirrors the role of miR-214 as an “oncogene” or “tumor suppressor gene.” The deregulation of miR-214 in lung cancer cells compared with normal lung cells may account for the aberrant growth and migratory behaviors of lung cancer cells.
To explore the underlying mechanisms of the effects of miR-214 in lung cancer, several genes have been examined previously as potential targets of miR-214. For example, PTEN was reported to be negatively regulated by miR-214, which was found to be responsible for cisplatin resistance in human ovarian cancer (Yang et al., 2008). Plexin-B1, which promotes the growth and invasion of HeLa cells, was also reported as a target of miR-214 in cervical cancer (Qiang et al., 2011). Other genes, such as p53/Nanog and Twist, were also reported as target of miR-214 (Li et al., 2012; Xu et al., 2012). In this study, we demonstrated that CPD is another target of miR-214. CPD has been unveiled recently as a significant regulator of cancer progression. Its upregulation was documented in a number of cancers, including prostate cancer (Thomas et al., 2012), breast cancer (Abdelmagid and Too, 2008), etc. However, studies of CPD expression on lung cancer are rare. We, therefore, first characterized the CPD expression in normal lung and lung cancer tissues and confirmed that CPD expression is higher in malignant tissues. In addition, CPD expression was positively correlated with the cancer grade. Then we focused on characterizing the correlation between CPD and miR-214. A seed matching site was found between CPD gene and miR-214 on the 3′ UTR site. Their interaction was confirmed by luciferase reporter assay. Interestingly, miR-214 expression and CPD expression seem to counteract each other, that is, upregulation of miR-214 would decrease CPD expression and CPD expression could reverse inhibitory effects of miR-214 in lung cancer. To our knowledge, this is the first report that correlated miR-214 and CPD expression in lung cancer. We established CPD as another target of miR-214, which could potentially help us unravel the mechanism of miR-214 in regulating lung cancer.
Diagnosis and treatment of lung cancer still remain challenges in clinics, due to the high metastatic potential and frequent recurrence of lung cancer. With our increasing knowledge on the role of miRNAs in regulating cancer progression, miRNAs have become a new class of diagnostic and therapeutic targets. In this study, we established miR-214 as a significant biomarker in lung cancer. This could inspire further studies to counteract the deregulation of miR-214, for example, by taking advantage of in vivo delivery systems to tumor sites (Rai et al., 2011; Wu et al., 2016). This could potentially provide a viable option for preventing lung cancer metastasis. Besides, new diagnostic methods could be developed, using miR-214 as a molecular target (Schwarzenbach et al., 2012; Han et al., 2015; Zhou et al., 2015). Clearly, the differential expression of miR-214 could yield a more accurate diagnosis to differentiate high-grade lung cancer from low-risk ones. Furthermore, it could potentially open doors to accurate prognosis for lung cancer patients. In addition, we also identified CPD as a hallmark of malignant and metastatic lung cancer, and CPD may also serve as a diagnostic and therapeutic target for lung cancer as well. Further investigations are necessary in a larger cohort of patients to consolidate these findings and potentially use miR-214 and CPD as diagnostic and therapeutic targets in combination with routine clinical tests.
Conclusion
We have identified miR-214 as a hallmark of malignant lung cancer. miR-214 regulates lung cancer progression using CPD as a target. New diagnostic or therapeutic strategies could be developed by targeting the miR-214-CPD axis.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.