
Abstract
Diffuse large B cell lymphoma (DLBCL) is a common B cell malignancy. Complete remission can be achieved in most patients by conventional treatment with rituximab and chemotherapy. However, a subset of remission individuals will develop a relapsed disease for obscure reasons. CD4+Foxp3−IL-10+ cell (Tr1) is a novel cell subtype with the capacity to suppress pro-inflammatory responses, but has not been extensively studied in most tumors. In this study, we investigated the potential role of Tr1 cells in DLBCL. We found that compared to that in healthy controls, the frequency of Tr1 cells was significantly increased in DLBCL patients, even during complete remission. Further study showed that these Tr1 cells were enriched in the CD25low/−Foxp3−CD49b+LAG-3+ fraction and could be developed in vitro from naive CD45RA+ CD4+ T cells. To examine the effect of Tr1 upregulation, we cocultured the enriched in vitro-induced Tr1 cells (iTr1) with autologous primary DLBCL cells and CD3+ T cells and found that iTr1 cells both enhanced the survival of CD20+ DLBCL tumor cells and suppressed the antitumor response of CD3+ T cells through the production of IL-10. Furthermore, the frequency of CD4+Foxp3−IL-10+ Tr1 cells in DLBCL patients during complete remission is directly associated with the risk of relapse. Together, these results suggested that Tr1 cells contributed to tumor cell maintenance and may serve as a prognostic marker and therapeutic target.
Introduction
D
Adaptive T cell response is an integral part of antitumor immunity. Increased presence and elevated activation status of CD4+ T cells in pretreatment tumor biopsies confer better survival in large B cell NHL (Ansell et al., 2001). Genetically modified T cells expressing anti-CD19 chimeric antigen receptors (CAR) are showing promise in treating refractory or relapsed DLBCL patients (Kochenderfer and Rosenberg, 2013; Kochenderfer et al., 2015). Interestingly, in many solid tumors, the CD4+CD25hiFoxp3+ regulatory T (Treg) cells suppress pro-inflammatory antigen presentation and antitumor effector T cell responses (Chen et al., 2005; Boissonnas et al., 2010), but in DLBCL, higher circulating and tumor-infiltrating Treg frequency was associated with better drug response and prognosis (Lee et al., 2008; Tzankov et al., 2008; Głowala-Kosińska et al., 2013). Another CD4+ T cell subset with characteristic interleukin 10 (IL-10), but not Foxp3 expression, termed T regulatory type 1 (Tr1) cell, also possesses the capacity to suppress pro-inflammatory responses, but has not been extensively studied in most tumors (Adeegbe and Nishikawa, 2013).
We hypothesized that this Tr1 population might contribute to the development and progression of DLBCL due to several reasons. First, Tr1 cells are elevated in associated lymph nodes in Hodgkin's lymphoma and could suppress cytotoxic T cell responses in PBMCs (Marshall et al., 2004). Second, Tr1 cells are induced in other tumors, such as glioma and head and neck squamous cell carcinoma, and are thought to mediate immune suppression through IL-10 production in the tumor microenvironment (Akasaki et al., 2004; Bergmann et al., 2007). And third, the Tr1 characteristic cytokine IL-10 is critical for the activation, expansion, and differentiation of B cells (Itoh and Hirohata, 1995; Ouyang et al., 2011). IL-10 not only promotes the proliferation of normal B cells after BCR and CD40 activation (Rousset et al., 1992) but also is required for the development and maintenance of spontaneous B cell lymphomas in New Zealand black mice (Czarneski et al., 2004). High IL-10 serum levels predict worse outcomes in patients with primary DLBCL (Gupta et al., 2012). Moreover, blocking IL-10 signaling induces tumor cell death in primary DLBCLs (Beguelin et al., 2015). Together, these results demonstrate that the role of Tr1 cells in the development of DLBCL needs to be explored.
In this study, we discovered an upregulation of circulating CD4+Foxp3−IL-10+ Tr1 cells in patients with DLBCL, even during complete remission. These CD4+Foxp3−IL-10+ Tr1 cells directly contributed to improved survival of primary DLBCL cells and suppressed the antitumor response by CD4+ and CD8+ T cells through IL-10 secretion. Interestingly, the frequency of CD4+Foxp3−IL-10+ Tr1 cells in DLBCL patients during complete remission was positively associated with the risk of relapse. These results suggested that Tr1 cells promoted the development and progression of DLBCL and might serve as a prognostic marker and therapeutic target.
Methods
Study participants and samples
Age- and sex-matched healthy controls (N = 13 [5 females and 8 males] aged between 30 and 70 years) and DLBCL patients (N = 100 [41 females and 59 males] aged between 28 and 68 years) provided peripheral blood samples. Surgical resection of tumor mass was performed in eight primary active DLBCL patients (Binn et al., 2003; Kim et al., 2011). Exclusion criteria included transformed follicular lymphoma, CNS involvement, history of HIV, HBV, and HCV infection, inadequate organ function, and concomitant malignancy. CHOP was administered in the standard dosage (cyclophosphamide 750 mg/m2 on day 1, doxorubicin 50 mg/m2 on day 1, vincristine 1.4 mg/m2 to a maximum of 2 mg on day 1, and prednisone 100 mg/m2 on days 1 through 5) every 21 days. Rituximab was administered at a dose of 375 mg/m2 7 and 3 days before cycle 1 and 2 days before cycles 3, 5, and, if administered, 7. Thirty-four patients achieving complete remission after four cycles were treated with a total of six cycles only, whereas all other patients received eight courses of R-CHOP. Complete remission was defined as the elimination of all lymphoma manifestations for at least 4 weeks, including from the bone marrow. Patients who otherwise fulfilled the criteria for complete remission, but without the bone marrow biopsy were not considered to have achieved complete remission and were taken off the study. Relapse was defined as new disease in complete remission patients. All participants provided written informed consent. Detailed patient information is presented in Table 1. The study was approved by the Ethics Committee of Liaocheng People's Hospital.
ABC, activated B cell-like; GCB, germinal center B cell-like; LDH, lactate dehydrogenase.
Peripheral blood was harvested by venipuncture at the upper arm and processed by Ficoll-Paque PLUS (GE Healthcare) density gradient centrifugation to obtain peripheral blood mononuclear cells (PBMCs). Resected tumors were first washed in PBS +10% penicillin-streptomycin (Thermo Fisher Scientific) and then in sterile PBS, mechanically disrupted through a metal sieve, and filtered through a 75-μm cell strainer. The resulting single cell suspension was first cultured in a T-75 flask for 7 days. Those that remained alive were used in experiments (Pulvino et al., 2012). For all cell cultures, 106 cells/mL were placed in RPMI 1640 supplemented with 10% FCS, 1% penicillin-streptomycin, and 2 mM L-glutamine (Thermo Fisher Scientific), in 37°C and 5% CO2.
Flow cytometry
PBMCs were stimulated with 1 μg/mL each of anti-CD3 (OKT3) and anti-CD28 (CD28.2), as well as 50 U/mL IL-2 for 12 h. 5 μg/mL each of brefeldin A and monensin (BD Pharmingen) were added 5 h before harvest. Cells were then washed in PBS +1% FCS, stained with Fixable Aqua Dead Cell Stain (Thermo Fisher Scientific) and combinations of anti-human CD3, CD4, CD8, CD20, CD25, CD45RA, CD45RO, CD49b, LAG-3, and CCR7 mAbs (BioLegend) for 30 min in 4°C before two washings. Fixation, permeabilization, and intracellular/nuclear staining of anti-human IL-10, Foxp3, and IFN-gamma mAbs (eBioscience) were done using the Foxp3/Transcription Factor Staining Buffer Set (eBioscience) according to manufacturer's instructions. Annexin V staining was conducted using the Annexin V Apoptosis Detection Kit eFluor 450 (eBioscience) following manufacturer's instructions. Events were acquired in BD LSR II cytometer and analyzed in FlowJo (Tree Star).
Cell isolation and coculture
CD3+ T cells, CD4+ T cells, CD45RA+ naive CD4+ T cells, and CD20+ tumor cells were purified by negative magnetic selection using EasySep Human T Cell, CD4+ T Cell, naive CD4+ T Cell, and B Cell Enrichment Kits (STEMCELL) according to manufacturer's instructions. CD49b+LAG-3+ sorting of purified CD4+ T cells was performed using the BD FACSAria cytometer. All cell types were plated at 1-to-1 ratios with each other in coculture experiments, to a final concentration of 106 cells/mL. Autologous PBMCs were irradiated at 5000 rad and used as feeder cells. Neutralizing anti-IFN-gamma (MD-1), anti-IL-12 (C8.6), and anti-IL-4 (MP4-25D2) mAbs were used at 5 μg/mL each. Neutralizing anti-human IL-10 (JES3-9D7) and IL-10R (3F9-2) mAbs and isotype controls were used at 10 μg/mL each. All mAbs were purchased from BioLegend except for 3F9-2, which was from Novus Biologicals.
Statistics
The paired t test, one-way ANOVA, Tukey's post test, and Gehan–Breslow–Wilcoxon test were performed in Prism software (GraphPad). Cox regression was performed in SPSS 13.0 software (IBM). Two-tailed p values less than 0.05 were considered statistically significant.
Results
Upregulation of CD4±Foxp3−IL-10±Tr1 cells started during active DLBCL and continued during remission and relapse
Tr1 cell is distinct from the conventional CD4+CD25hiFoxp3+ Treg cell in its extrathymic origin, lack of Foxp3 or other exclusive transcription factors, and high IL-10 expression (Roncarolo et al., 2014; Zeng et al., 2015). In this study, we first identified circulating Tr1 cells as CD4+Foxp3−IL-10+ T cells (Fig. 1A) and examined their frequency in healthy individuals (healthy), untreated active primary DLBCL patients (primary), treated DLBCL during complete remission (remission), and relapsed DLBCL patients (relapse). CD4+Foxp3−IL-10+ T cells could be detected at very low level in healthy individuals, but were significantly increased in active DLBCL patients, regardless of primary or relapsed status. The frequency of CD4+Foxp3−IL-10+ T cells in remission subjects was lower than that in primary and relapse patients, but was still significantly elevated compared to that in healthy subjects (Fig. 1B). Together, these results demonstrated a dysregulation of CD4+Foxp3−IL-10+ T cells (hereafter Tr1 cells) in DLBCL, during both the active disease and remission.
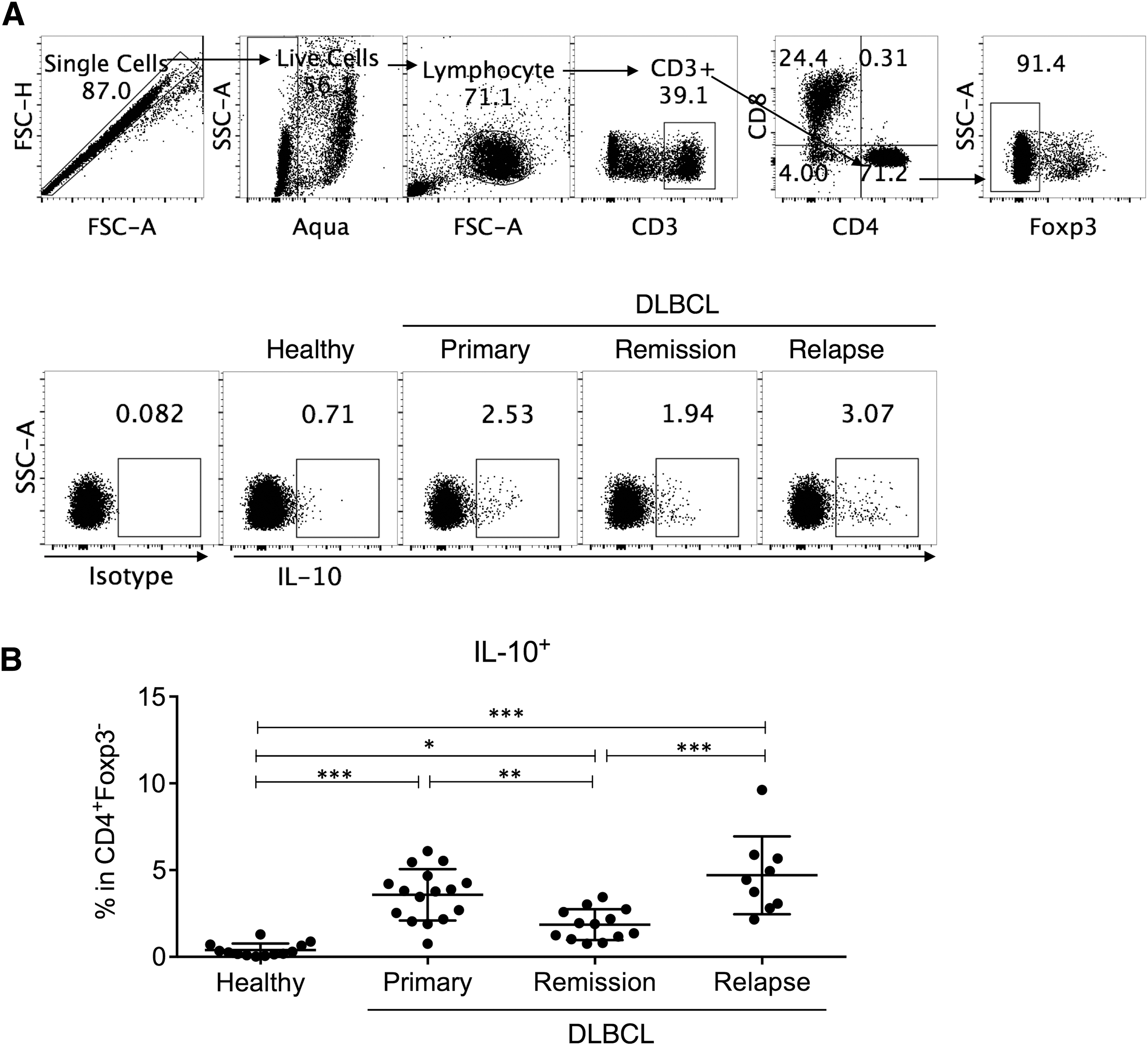
DLBCL patients had higher circulating Tr1 cells than healthy controls.
CD4±Foxp3−IL-10±Tr1 cells were enriched in the CD49b±LAG-3±fraction
To examine the activation status and allow further analysis of the function of Tr1 cells, we examined the surface marker expression of CD4+Foxp3−IL-10+ Tr1 cells. Both naive (CD45RA+) and memory (CD45RO+) cells were found in Tr1 cells from DLBCL patients. Most Tr1 cells also had low or negative CD25 and CCR7 expression (Fig. 2A, B). CD49b and LAG-3 expression was previously associated with Tr1 cells in human and mouse (Gagliani et al., 2013). In this study, we found that the CD4+Foxp3−IL-10+ Tr1 cells were enriched in the CD49b+LAG-3+ T cell fraction (Fig. 2A, B).
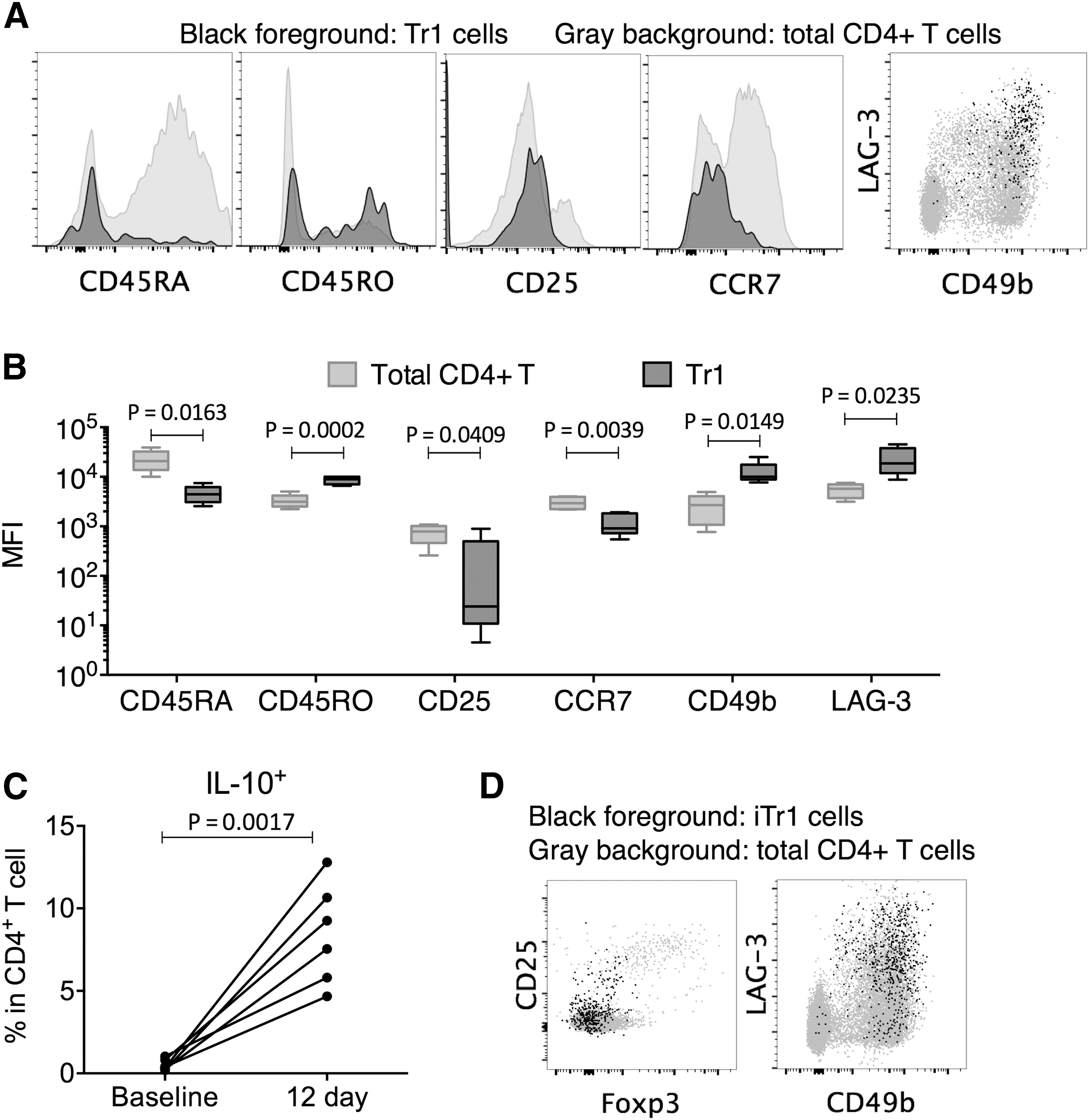
Tr1 cells and iTr1 cells were enriched in the CD25low/−Foxp3−CD49b±LAG-3±CD4± T cell fraction.
We next investigated the mechanism of Tr1 induction. We stimulated purified CD45RA+ naive CD4+ T cells by anti-CD3 and anti-CD28 mAbs and IL-2, in the presence of neutralizing anti-IFN-gamma, anti-IL-12, and anti-IL-4 to prevent the induction of other CD4+ T cell subsets (Barrat et al., 2002). After 12 days in the presence of irradiated feeder cells, the level of IL-10+ T cells was significantly elevated (Fig. 2C). These IL-10+ T cells were concentrated in the Foxp3−, CD25low/−, CD49b+, and LAG-3+ compartments, demonstrating that they resembled naturally occurring Tr1 cells in DLBCL patients (Fig. 2D). We therefore referred to these in vitro stimulated CD4+Foxp3−IL-10+ T cells as induced Tr1 (iTr1) cells and used them in subsequent experiments, since the supply of Tr1 cells was limited.
Secretion of IL-10 by CD49b±LAG-3±-enriched iTr1 cells promoted the survival of DLBCL cells and suppressed antitumor T cell responses
To investigate the impact of Tr1 upregulation in DLBCL, we used CD49b and LAG-3 as surface markers to enrich iTr1 cells, which were then cocultured with autologous primary DLBCL cells and/or autologous CD3+ T cells. Primary DLBCL cell only culture was also maintained as a control. The DLBCL tumor cell apoptosis was assessed by annexin V binding to externalized phosphatidylserine (Fig. 3A). We found that the addition of iTr1 cells significantly improved the survival of DLBCL cells, as evident by the lower annexin V binding on CD20+ tumor cells in the DLBCL+iTr1 culture compared to those in the DLBCL alone culture (Fig. 3B). Interestingly, presence of CD3+ T cells suppressed the survival of DLBCL cells, but the addition of iTr1 reverted this effect. This was based on the observation that the annexin V levels on CD20+ tumor cells in the DLBCL+CD3+ T culture were higher than in the DLBCL alone culture, but were not different between the DLBCL alone culture and the DLBCL+iTr1+CD3+ T culture (Fig. 3B).
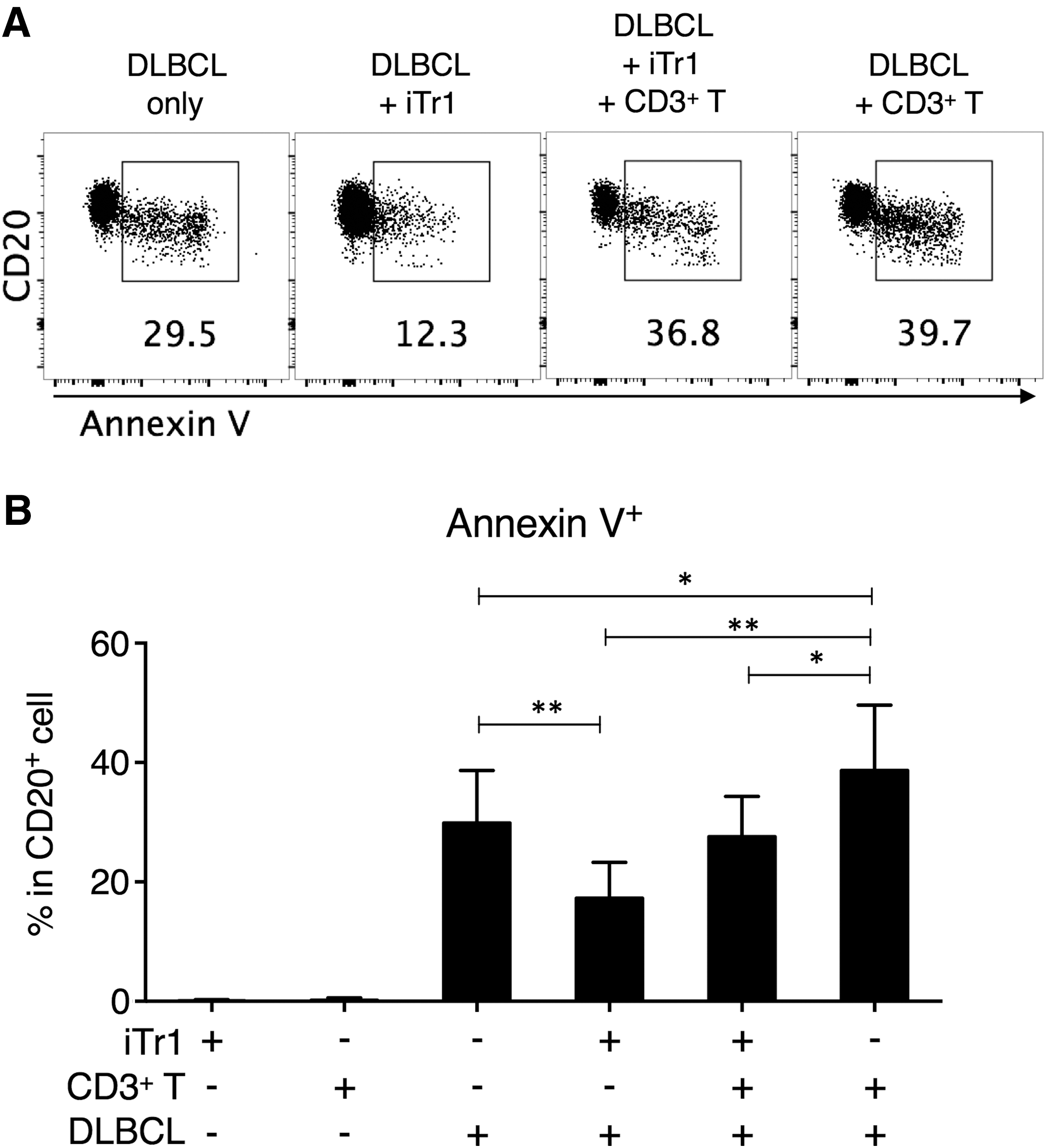
iTr1 cells improved the survival of DLBCL tumor cells.
The above results suggested that iTr1 cells could promote the survival of DLBCL tumor cells, as well as suppress antitumor activity by T cells, but the underlying mechanism of these effects was not examined. We thought that IL-10 possibly contributed to the effects of iTr1. Therefore, we repeated DLBCL+iTr1+CD3+ T coculture experiments with neutralizing anti-IL-10 and anti-IL-10R blocking antibodies or matching isotype controls. The annexin V levels on CD20+ tumor cells, as well as the IFN-gamma production by cocultured CD3+ T cells, were examined (Fig. 4A). We found that blocking IL-10 signaling had significantly elevated the annexin V level on CD20+ tumor cells, as well as increased the IFN-gamma production by cocultured CD4+ and CD8+ T cells (Fig. 4B).

iTr1 cell improved CD20± tumor cell survival and suppressed CD4± and CD8± T cell IFN-gamma response through IL-10 signaling.
High frequency of CD4±Foxp3−IL-10±Tr1 cells was associated with high relapse risk
The effect of Tr1 cells in improving DLBCL survival suggests that the presence of Tr1 cells in vivo could result in leftover tumor cell clones during remission and increase the risk of relapse. To examine this hypothesis, we recruited more DLBCL patients and examined their circulating CD4+Foxp3−IL-10+ Tr1 cell frequency during complete remission. The patients were then divided into High Tr1 (median: 5.98%, range: 4.00% −8.82%) and Low Tr1 (median: 2.18%, range: 1.03% −3.98%) groups using median Tr1 frequency (3.99% in circulating CD4+Foxp3− T cells) as the dividing line. We found that the High Tr1 group had significantly shorter remission time than the Low Tr1 group (Fig. 5). The High Tr1 group also had higher relapse rate than the Low Tr1 group (51.6% vs. 25.8%) after 2 years. We also examined the effect of various prognostic factors in these patients using Cox regression (Table 2). No statistical significance was found, possibly due to the small sample size.
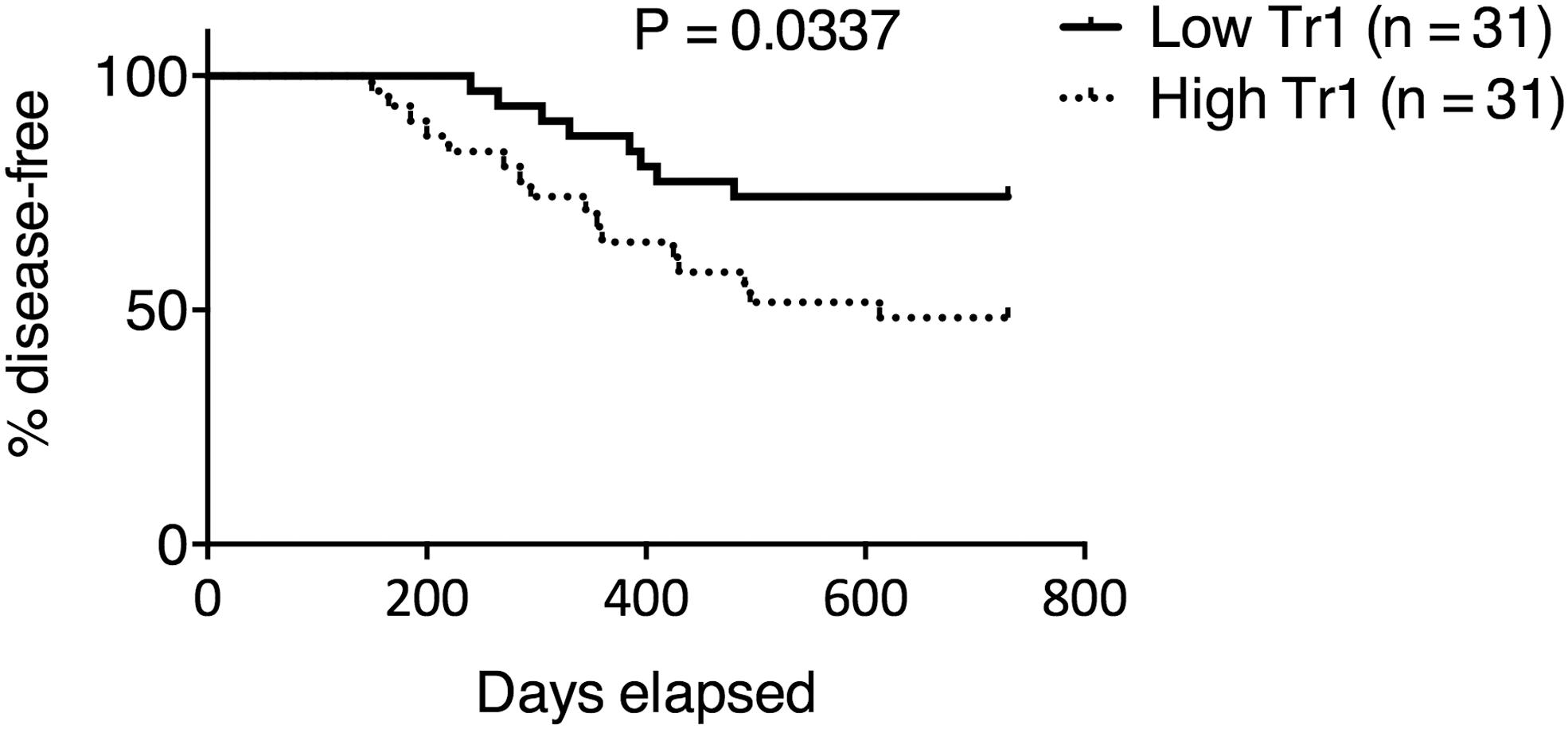
DLBCL patients with higher Tr1 cell frequency had shorter remission and higher relapse rate. CD4+Foxp3−IL-10+ Tr1 cell frequency was determined 4 weeks (28–30 days) after the last treatment course in 62 DLBCL patients achieving complete remission, who were then divided into High Tr1 and Low Tr1 groups according to the median value (3.99% in CD4+Foxp3− T cells). Events of relapse were monitored for 2 years. Gehan–Breslow–Wilcoxon test.
CI, confidence interval.
Discussion
In this study, we first showed that the frequency of circulating CD4+Foxp3−IL-10+ Tr1 cells was upregulated in DLBCL patients, regardless of the disease status. Although the frequency of Tr1 cells in remission DLBCL patients was reduced compared to that in active primary and relapsed DLBCL patients, it was still increased compared to healthy subjects. The underlying mechanism that resulted in this upregulation was not explored. Previous studies showed that CD4+ T cells under chronic antigen stimulation and in the presence of IL-10 could develop into Tr1 cells (Groux et al., 1997; 1999). In solid tumors such as glioma, overexpression IL-10 and TGF-beta by tumor dendritic cells elicited Tr1 responses (Akasaki et al., 2004; Bergmann et al., 2007). Interestingly, B NHL cells could directly secrete large amounts of IL-10 and were considered a major source of IL-10 in the serum (Voorzanger et al., 1996). It is therefore possible that DLBCL cells directly induced Tr1 development as an immune evasion strategy. Previously, Epstein–Barr virus (EBV) infection was shown to induce the gene expression of Tr1 cell-related markers, such as ITGA2, ITGB2, LAG3, and IL-10, in Hodgkin's lymphoma (Morales et al., 2014). EBV-positive DLBCL also accounted for approximately 9–11% of Asian DLBCL cases (Park et al., 2007). Whether EBV infection contributed to Tr1 upregulation in our DLBCL patients would require further studies.
To obtain sufficient Tr1 cells and allow further functional analysis of Tr1 cells, we mimicked antigen-specific stimulation by anti-CD3/CD28 mAbs and depleted the IL-4, IL-12, and IFN-gamma, which resulted in the differentiation of naive CD4+ T cells to iTr1 cells that closely resembled naturally occurring Tr1 cells in IL-10 production and surface marker expression. These iTr1 cells directly enhanced DLBCL survival, since the annexin V staining on CD20+ tumor cells was higher in DLBCL alone cultures than in DLBCL+iTr1 cultures. This effect was mediated by IL-10, since blockade of IL-10 signaling elevated annexin V binding on CD20+ tumor cells.
However, this explanation may not represent the full picture of Tr1-mediated improvement on DLBCL cell survival, since the source of IL-10 could be DLBCL cells as well. We also showed that the addition of iTr1 cells into the DLBCL+CD3+ T cell culture suppressed the antitumor effects of CD3+ T cells, while blockade of IL-10 signaling reverted this suppression. Again, Tr1 cells might have utilized inhibitory mechanisms other than direct IL-10-mediated suppression of effector T cells, some of which are difficult to study in vitro, but can be examined in animal models. For example, a longitudinal study in HIV-1 infection showed that early induction of Tr1 cells was responsible for subverting the general inflammatory status toward the regulatory type, with increased CTLA-4 expression in all regulatory T cell subsets (Chevalier et al., 2015). Tr1 cells could also inhibit dendritic cell maturation and induce dendritic cell tolerance (Manavalan et al., 2003; Gregori et al., 2010), which in turn promotes regulatory T cell differentiation and suppresses effector T cell activation. Furthermore, granzyme B and perforin release was discovered in Tr1 cells and was responsible for the elimination of antigen-presenting cells (Magnani et al., 2011). In the future, these in vivo mechanisms of Tr1 cells should be examined since our experiments demonstrated a strong upregulation of Tr1 cells in all stages of DLBCL.
The high Tr1 level during complete remission suggested that the Tr1 response could be maintained for an extended period of time, as evident by the observation that a subset of naturally occurring Tr1 cells expressed the memory marker CD45RO. Interestingly, the frequency of Tr1 cells in DLBCL patients who just achieved complete remission predicted the duration of remission and the risk of relapse. This suggested that depletion of Tr1 cells might be beneficial to the overall outcome of DLBCL, even when the patient is responding well to standard therapy. Notably, this is in striking contrast with the conventional CD4+CD25hiFoxp3+ Treg cells, which were thought to improve the prognosis of DLBCL. Thus, the possibility of incorporating Tr1-depletion, potentially by removing CD49b+LAG-3+ T cells, in the standard treatment of DLBCL should be investigated. High Tr1 frequency might simply be reflecting a generally immunosuppressive environment with low likelihood of eliminating recurrent tumor cells. Or, Tr1 cells could have directly helped tumor cells to persist and expand by secreting IL-10. The mechanism by which Tr1 possibly increased the risk of relapse should be examined as well.
Footnotes
Acknowledgment
The authors thank Dr. Smith at DICAT Biomedical Computation Centre for assisting in data analysis.
Disclosure Statement
No competing financial interests exist.