
Abstract
Selenium—as a trace element—is nutritionally essential for humans. It prevents cancerous growth by inhibiting the telomerase activity but the mechanism involved in regulation of telomerase activity in normal telomerase-positive cells remains to be elucidated. Here, we find out whether the effect of sodium selenite and selenomethionine on telomerase activity in human umbilical cord-derived mesenchymal stem cells (hUCMSCs) is associated with different levels of c-Myc and p53 expression. The use of different staining methods including ethidium bromide/acridine orange and DAPI in addition to telomeric repeat amplification protocol assay and real-time PCR indicated that different forms of selenium have opposite impacts on c-Myc and p53 expressions in both hUCMSCs and AGS, a gastric adenocarcinoma cell line, as a positive control. Our findings suggest that the signaling pathways involved in the regulation of telomerase activity in malignant and normal telomerase-positive cell types are somewhat different, at least on the c-Myc and P53 expression levels.
Introduction
S
Telomerase is composed of two main subunits: TERT and TERC. Unlike TERC, which is expressed in most cells and serves as a template for telomere replication, human telomerase reverse transcriptase (hTERT), a catalytic subunit of the human telomerase, is an effective factor in regulation of telomerase activity (Sandin and Rhodes, 2014). Recent data demonstrate a close correlation between hTERT expression and telomerase activity in human tumors (Rao et al., 2011; Lue and Autexier, 2012; Wong et al., 2014). Thus, the hTERT may be an excellent candidate for cancer therapy (Holysz et al., 2013), and it was found that the promoter of hTERT has binding sites for c-Myc and other transcription factors (Kyo et al., 2008; Calado, et al., 2009; Zhao et al., 2014; Lewis and Tollefsbol, 2016). In addition, some trace elements like selenium can regulate its expression. There is evidence reporting a link between overexpression of P53 as a transcription factor involved in apoptotic pathways and c-Myc at a relatively high dose of selenium in rat hepatocyte (Yu et al., 2009). Some researchers believed that both c-Myc and p53 overexpression—mediated by oxidative stress—may represent a key role in selenium-induced apoptosis (K, 2006; Yu et al., 2006a, 2006b; Rudolf, 2008). Various investigations also suggested that the different forms of selenium have their own unique sets of abilities for telomerase activity regulation in different cell types at the upstream or downstream levels (Chen et al., 2007; Yu et al., 2009; Zarghami, et al., 2011). It has been shown that the sodium selenite activates p53 at the transcriptional level in prostatic cancer cells to induce cell death (Sarveswaran et al., 2010). The downregulation of c-Myc has also been observed in colorectal carcinoma cell line HCT116 following selenomethionine treatment (Goulet et al., 2007). Moreover, both organic and inorganic forms of selenium promoted HL-60 cell cycle progression through overexpression of some cell cycle-related genes, especially c-Myc (Zeng et al., 2002). Although some old studies have indicated the relationship between telomerase activity and c-Myc/P53, especially in selenium-exposed cancerous cells, the mechanism by which telomerase regulation is associated with c-Myc and P53 has not been fully understood yet. There is no direct report indicating the regulation of telomerase function by these transcription factors in normal telomerase-positive cells when exposed to selenium compounds. Therefore, in this study, we aimed to use the human umbilical cord-derived mesenchymal stem cell (hUCMSC) as a telomerase-positive fetal stem cell model to determine the impact of different selenium forms on c-Myc and p53 expression profiles and their association with telomerase activity. We found that sodium selenite upregulates c-Myc/p53 expression and induces the higher telomerase activity in hUCMSCs, but selenomethionine shows an opposite effect and decreases c-Myc/p53 expression and telomerase activity.
Materials and Methods
Cell culture
Human Wharton's jelly-derived mesenchymal stem cells (the hUCMSCs) were isolated and cultured as previously described (Saraee et al., 2014). In brief, after obtaining fresh umbilical cord from full-term neonates (in accordance with the ARUMS ethical committee's considerations), they were dissected into small pieces and transferred to a T-25 flask containing DMEM medium (ATOCEL; ATCDL-891) supplement with 20% fetal bovine serum (FBS, Gibco;10270-098) and 2% Pen/Strep/AmphoB (ATOCEL, ATRB-010) and finally incubated at 37°C and 5% CO2. By 20 days postculturing, the remaining explants were removed and the growing hUCMSCs were subcultured in DMEMLG with 10% FBS. This stage was denoted as Passage 0. Subsequent passages of the hUCMSCs were obtained by sequential subculturing. The AGS, a gastric adenocarcinoma cell line as a human gastric cancer cell model was expanded in RPMI1640 medium (ATOCEL, ATCR-885) supplement with 10% FBS and 1% Pen/Strep/AmphoB at 37°C under a humidified atmosphere of 5% CO2.
Experimental design
As shown in Figure 1, the hUCMSCs at a density of 1000 cells/well were seeded in 96-well tissue culture plates and treated with different concentrations of sodium selenite (NaSel, Sigma; S5261), ranging from 1 to 80 μM dissolved in phosphate-buffered saline [PBS]) and selenomethionine (SeMet, Sigma; S3132) at a range of 0.5–2000 μM. Untreated cells were considered a control group.
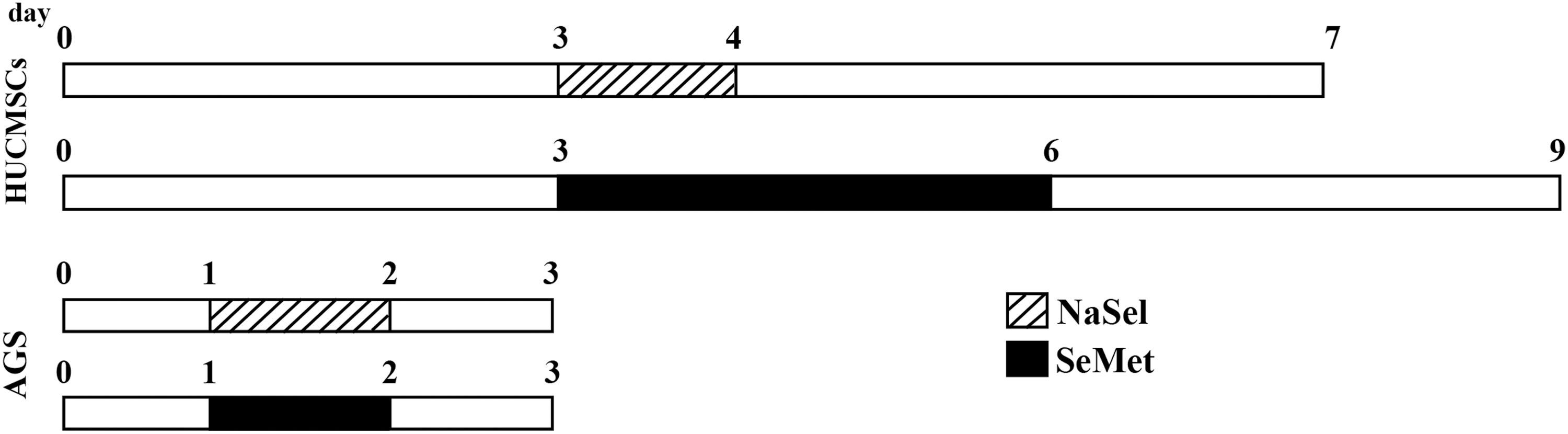
The experimental design for AGS, a gastric adenocarcinoma cell line and hUCMSCs culture and treatment with different concentrations of NaSel and SeMet. Pretreatment time for each cell type has been calculated based on the cell specific-doubling time. hUCMSC, human umbilical cord-derived mesenchymal stem cell.
Cell viability tests
To investigate the viability rate of hUMCSCs at different doses of NaSel and SeMet and to obtain the optimum dose at which the cells are less sensitive to the cytotoxicity from selenium compounds, MTT assay, DAPI, and ethidium bromide/acridine orange (EB/AO) staining methods were used as previously described (Saraee et al., 2014). For the quantification of the cell death rate, 15 microscopic fields were chosen at ×20 magnification and the photos were taken with an inverted fluorescence microscope (Olympus IX71, Japan) equipped with a digital camera (Olympus; DP71).
Flowcytometry analysis
Characterization of hUCMSCs by flowcytometry (Partec, CyFlow) has been done as previously described (Saraee et al., 2014) by using different positive (CD44-FITC, CD73-PE, CD90-PE, and CD105-FITC) and negative (CD34-PE and CD45-FITC) (BD Pharmingen, England) stemness markers. To analyze apoptosis, DAPI (40,6-diamidino-2-phenylindole, Gerbu,1050) staining were used. For this purpose, the hUCMSCs were precultured for 72 h and treated with 3, 10, and 20 μM of NaSel for 24 h and 500 μM SeMet for 72 h. Then, the cells were detached from the culture dish by 0.05% trypsin/EDTA, centrifuged at 12,000 rpm for 5 min, and exposed to DAPI (1 μg/mL) for 30 min. Finally, to demonstrate the percentage of apoptotic cells, cell cycle analysis was performed.
TRAP assay
Telomeric repeat amplification protocol (TRAP assay) is a qualitative and PCR-based telomerase assay that can evaluate the telomerase activity. A pellet of 105 cultured cells from both untreated and treated groups was washed with PBS and homogenized in 200 μL of ice-cold Chaps lysis buffer. After incubating at −20°C for 30 min, the cell lysate was centrifuged at −4°C and the resultant supernatant was stored for subsequent usage or aspirated to define the protein concentration by Bradford protein assay kit (Bio-Rad, B6916). In the next step, 1 μg of protein was added to a TRAP master mix containing TS primer (in a final volume of 45 μL) and incubated at room temperature for 30 min to extend the substrate by telomerase and inactivated at 95°C for 5 min. Then, 5 μL of a mixture of 2.5 U Taq DNA polymerase, 50 ng ACX, 50 ng NT, and 1.3 × 10−9 ng TSNT primers were added to the extended product and finally subjected to PCR condition as follows: 94°C for 30 s, 52°C for 60 s, and 72°C for 60 s. Then, the PCR product was electrophoresed on 10% nondenaturing polyacrylamide gels. In this assay, the AGS cell line was used as a positive control showing high telomerase activity.
Quantitative real-time TRAP assay
For quantitative analysis of the telomerase activity by real-time PCR (qRT-TRAP Assay), 1 μL of the extracted protein was added to 24 μL solution containing 1× SYBR Green buffer, 50 ng TS, and 50 ng ACX primers. After incubation at room temperature for 30 min, the amplified product was run in a thermal cycler (Roche; 05815916001) for 40 cycles at 95°C for 10 min (to activate the hot-start Taq polymerase in the master mix), 95°C for 30 min, and 60°C for 60 s.
RNA extraction and complementary DNA synthesis
Total RNA was isolated from the cells by Biozol (BioFlux; BSC51 M1) reagent in accordance with the manufacturer's protocol. To eliminate any DNA contamination, the extracted RNA was treated with the DNase-I enzyme (Cinnaclon, PR891627C). Then, the complementary DNA (cDNA) from 1 μg DNase-I digested RNA was synthesized by using RevertAid™ H Minus First Strand cDNA Synthesis kit (Fermentase, K1622). The samples were stored at −20°C until analysis via quantitative real-time polymerase chain reaction (qRT-PCR).
Quantitative real-time PCR
Quantitative real-time PCR was performed by light cycler real-time PCR detection system (Roche; 05815916001). Briefly, 1 μL of each cDNA template was added to a mixture containing 1× SYBR Green buffer (EURx; E0402-01) and 400 nM of each forward and reverse primer. The primer sequences and the PCR product sizes were shown in Table 1. Real-time PCR reactions were carried out as follows: 10 min at 95°C for one cycle; 30 s at 95°C, 60 s at 60°C for 40 cycles. Then, the melting curve for each PCR product was analyzed. Fold changes in gene expression for treated and untreated groups were determined by using the 2−ΔΔCT method and the housekeeping gene, GAPDH as an internal control. The real-time PCR reactions were done in triplicate and again, AGS cell line was used as a positive control group expressing the desired genes.
hTERT, human telomerase reverse transcriptase; qRT-PCR, quantitative real-time polymerase chain reaction.
Statistical analysis
The experimental assays for quantification were carried out at least three independent repeats and the resulted data were expressed as mean ± standard error of mean. Then, based on the collected data, two-independent Student's T-test or One-way ANOVA followed by Tukey's post-hoc multiple group comparison test were used to statistically analyze different groups. A difference between groups was considered statistically significant if p < 0.05.
Results
Effect of NaSel and SeMet on hUCMSCs morphology
Following culture and expansion of the hUCMSCs up to P2 and characterizing them by several mesenchymal stem cell markers, they were exposed to the selenium compounds, that is, NaSel and SeMet at different doses to find whether these selenium agents have any effects on cell morphology and viability. As depicted in Figure 2, our results clearly illustrated that the hUCMSCs expressed the mesenchymal stem cell's markers CD44 (69.22%), CD73 (99.86%), CD90 (98.35%), and CD105 (74.05%) but very few cells expressed the hematopoietic stem cell's markers CD34 (0.44%) and CD45 (7.51%). Moreover, NaSel at 3 μM altered the cell morphology to a slender shape but most of them were alive. However, treatment of hUCMSCs with NaSel at the higher concentration (10 μM) widely induced cell death, so that many cells represented a round and marginal bright configuration, suggesting many cells had undergone to e apoptosis. In contrast, hUCMSCs exposed to SeMet at different doses (0.5 and 1 mM) showed no obvious changes in cell morphology and represented a resistance even at the higher doses.

Treatment of the hUCMSCs with different concentrations of selenium compounds.
Effect of NaSel and SeMet on cell viability
To investigate the viability rate of hUCMSCs when treated with different doses of selenium compounds and to find the optimum concentrations at which the cells are more stable and less sensitive to NaSel and SeMet cytotoxicity, MTT assay was used. Our results indicated that mean viability rate of the cells—when exposed to the different concentrations of NaSel for 24 h—significantly decreased from 78.3% ± 2.95% and 55.93% ± 5.08% at 3 and 10 μM to 6.9% ± 1.81% and 1.48% ± 0.3% at 20 and 80 μM, respectively (Fig. 3A, p < 0.01). The hUCMSCs treated with SeMet for 72 h from 0.5 to 2000 μM showed a different story in which no significant changes in the cell viability rate was obtained. According to our data, we selected 3 μM NaSel and 500 μM SeMet as the optimum amounts at which the cells were resistant to cell death and about 80% of them were alive.

Effect of selenium supplementation on the cell viability rate.
To check the cytoplasmic and nuclear changes in hUCMSCs when exposed to the aforementioned optimum quantities of the selenium agents, the treated cells were stained with EB/AO. As shown in Figure 4, tiny blebbed cytoplasmic changes and cell shrinkage along with nuclear fragmentations were seen in the cells treated with 3 μM NaSel, but these morphological alterations enhanced in the cells exposed to 10 μM, suggesting early and late apoptosis and necrosis injury occurred at the higher quantity (Fig. A1–D2). These observations were confirmed by DAPI staining quantification through which the cell death frequency significantly increased from 20.38% ± 3.79% at 3 μM to 34.38% ± 4.46% at 10 μM NaSel (p < 0.01, Fig. 4E). Furthermore, when the cells treated with 500 μM SeMet, only% 3.32 ± 0.66 of them showed apoptosis compared to 3.04% ± 0.98% of cells in control group (p > 0.05) and EB/AO staining demonstrated no apparent difference between the treated and untreated cells. Analysis of DAPI- stained apoptotic cells and cell cycle by flowcytometry clearly showed that NaSel at the higher doses (10 and 20 μM) induced apoptosis in 55.23% and 62.45% of cells in comparison with 3 μM concentration at which 39.15% of cells underwent apoptosis. As expected, just 8.48% of hUCMSCs indicated cell death in the SeMet (500 μM)-treated group (Fig. 4F).
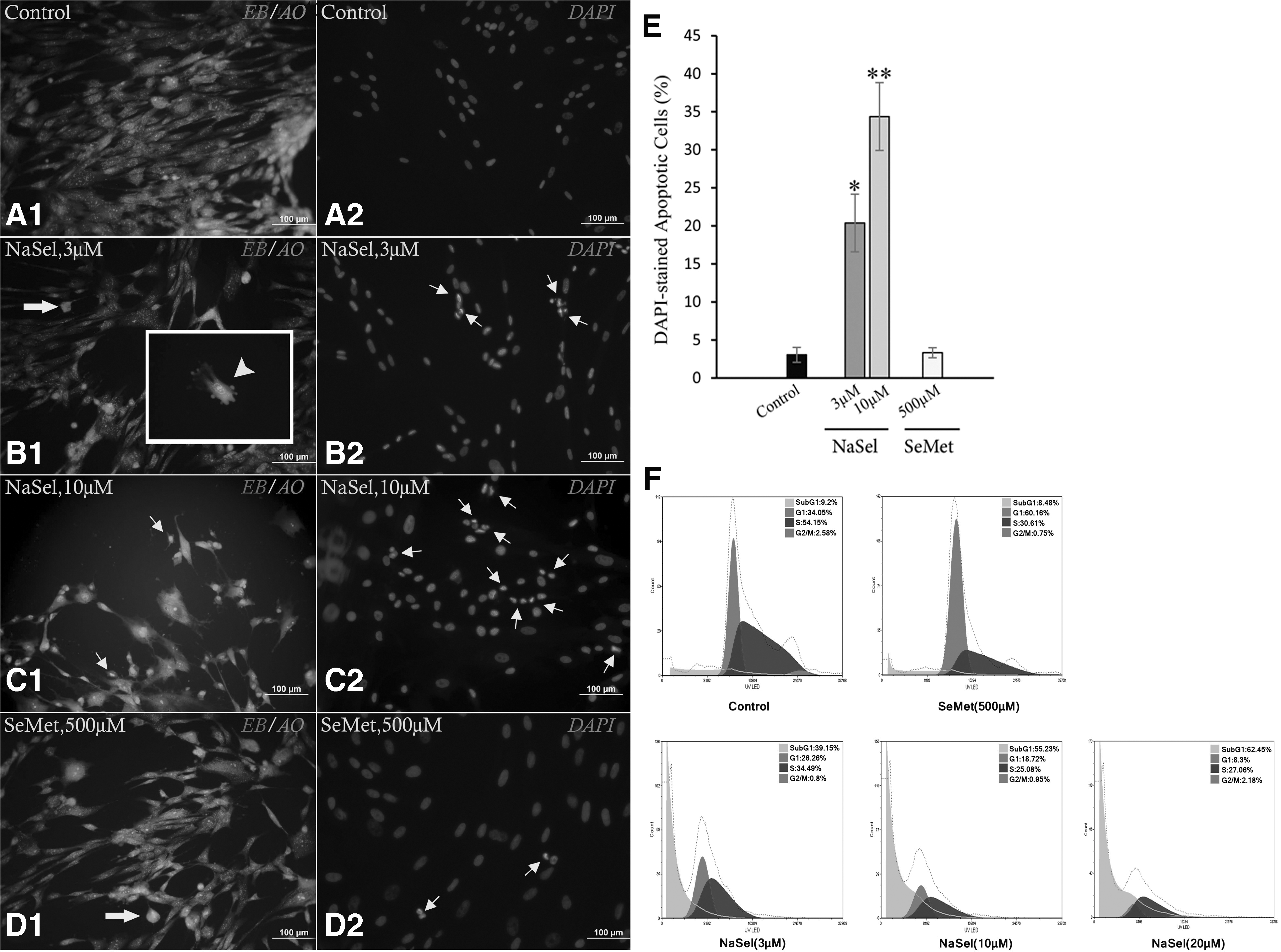
Assessment of the hUCMSCs cytoplasmic and nuclear alterations by EB/AO and DAPI staining methods.
Effect of NaSel and SeMet on telomerase activity in the hUCMSCs
To investigate the effect of NaSel and SeMet on telomerase activity, TRAP assay and quantitative real-time TRAP assay were performed (Fig. 5). Since 3 μM NaSel and 500 μM SeMet showed very low cytotoxic effects on hUCMSCc, in the remaining experiments they were considered as the optimum doses influencing the cells. As expected, by using the TRAP assay, the hUCMSCs obviously showed high telomerase activity, which upregulated upon treating with NaSel and downregulated when the cells exposed to SeMet. To quantify the Ct at which the treated cells indicate alterations in telomerase activity, qRT-TRAP assay calculated 22.25 ± 0.9 and 31.99 ± 1.01 Ct in NaSel and SeMet-exposed cells, respectively. These data were significantly different from the Ct for the untreated cells (27.05 ± 0.58, p < 0.05) (Fig. 5B).
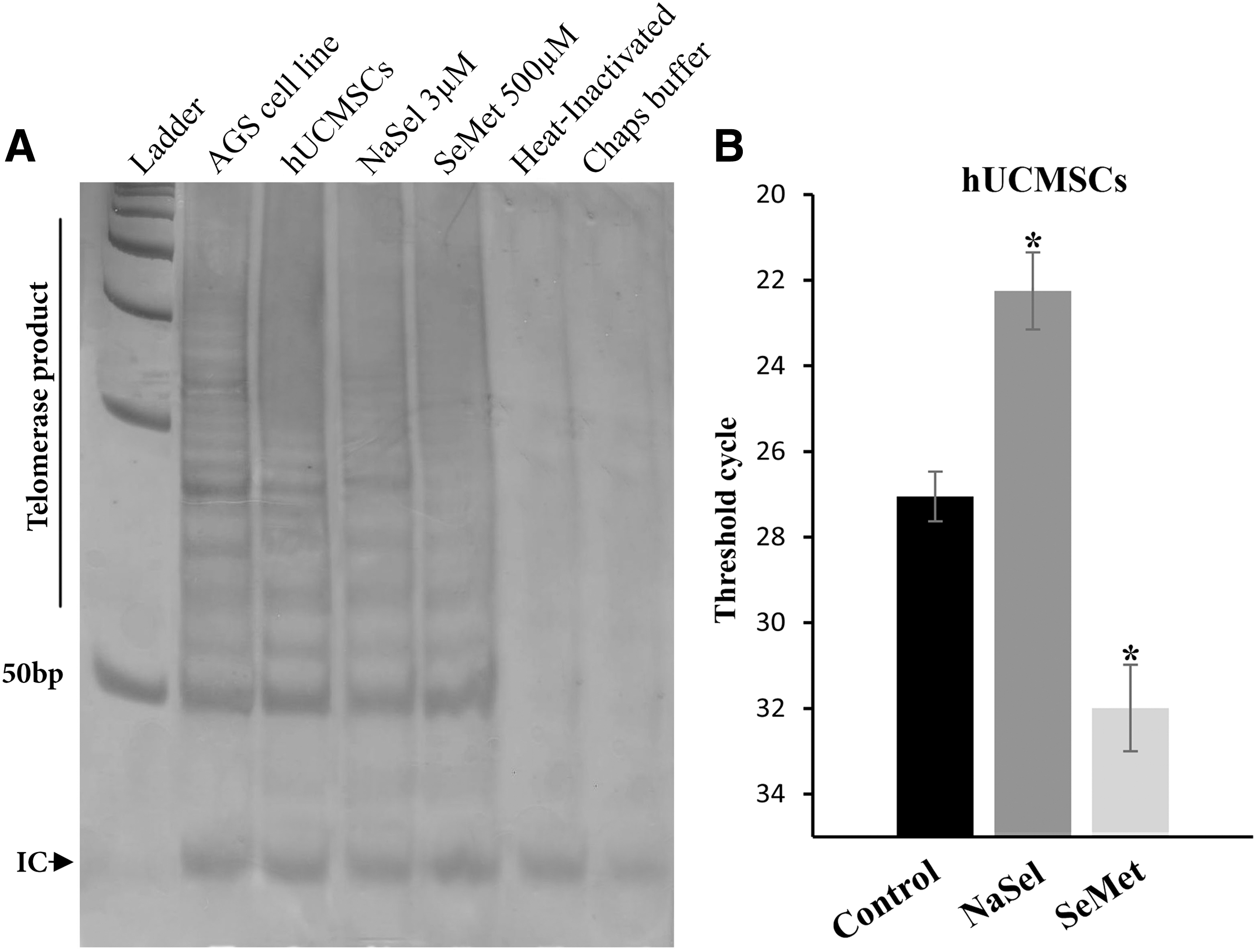
Effect of selenium compounds on the telomerase activity in hUCMSCs.
Effect of NaSel and SeMet on c-Myc and P53 mRNA expression in the hUCMSCs
It is known that telomerase activity is regulated by c-Myc and P53. Moreover, exposure to different forms of selenium appears to have controversial effects on the genes noted. To address this possibility, as indicated by Figure 6, real-time PCR analysis obviously represented that c-Myc expression in the NaSel-treated hUCMSCs at the mRNA level significantly increased to 1.83 ± 0.29-fold changes compared to the untreated cells (p < 0.05). In contrast, its expression in NaSel-treated AGS was downregulated to 0.26 ± 0.21 compared to the untreated AGS cells (p < 0.05). Similar results were obtained for p53 expression level when both the cells were treated with NaSel (2.13 ± 0.21-fold changes for the hUCMSCs and 0.28 ± 0.14 for the treated AGS cells, [p < 0.05 vs. both control groups]). The organic form of the selenium revealed the opposite impacts on c-Myc and P53 expression in the hUCMSCs. The expression of c-Myc in SeMet-treated hUCMSCs was found to have changes around 0.73 ± 0.13-fold, but was significantly downregulated to 0.33 ± 0.13 in SeMet-exposed AGS cells in comparison with the untreated cells (p < 0.05). No distinct alteration in p53 expression was detected in SeMet-exposed hUCMSCs (0.96 ± 0.18), but p53 expression again decreased to 0.2 ± 0.02-fold changes in SeMet-treated AGS cell.
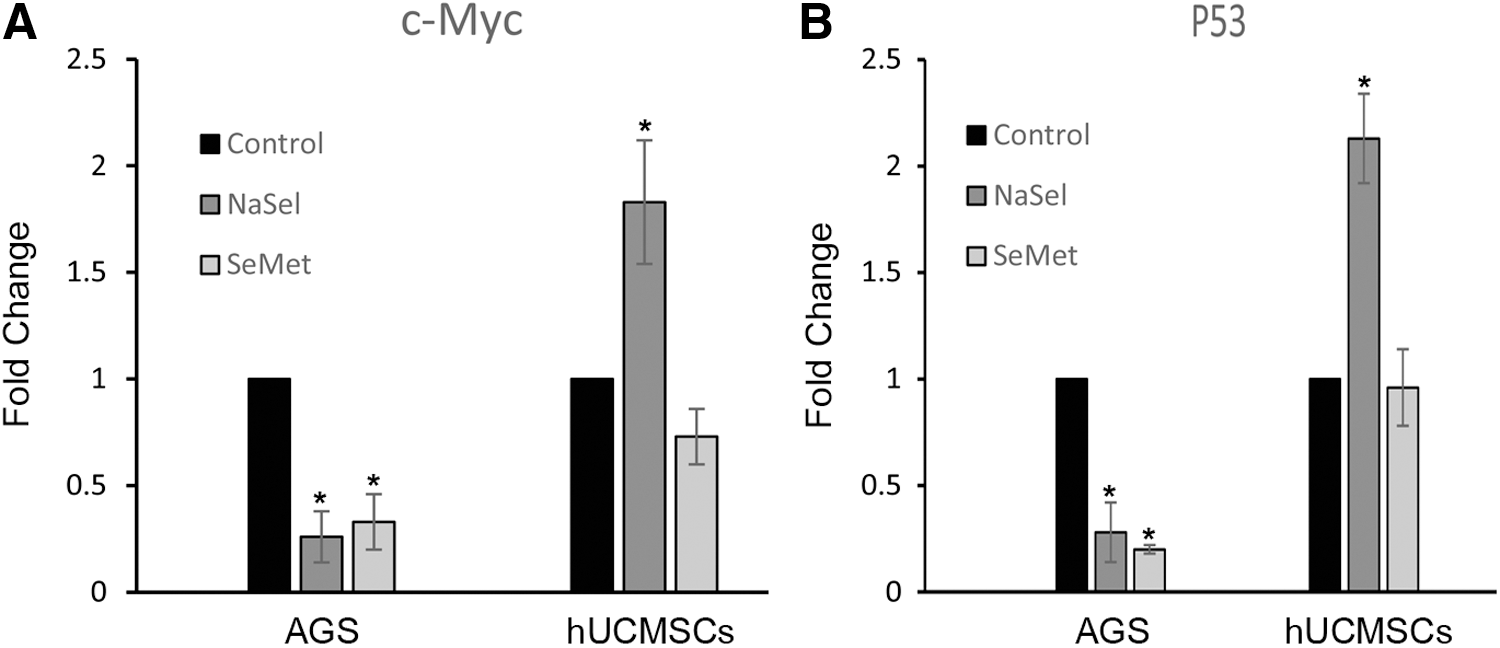
Effects of NaSel and SeMet on c-Myc
Discussion
In this study, we presented a relationship between telomerase activity and levels of c-Myc/p53 expression in the hUCMSCs as the fetal stem cells, which are similar to cancerous cells, are telomerase-positive cells. We also reported evidence that NaSel at physiological concentration has an opposite effect on hUCMSCs compared to AGS cancer cell line to enhance telomerase activity through upregulating c-Myc/p53 expression but the effect of SeMet on both cell types appeared to be the same. While some reports suggest that sodium selenite can extend the telomere length and increased telomerase activity in hepatic cell line and yeast cells (Liu et al., 2004; Yu et al., 2009) in contrast, studies on breast cancer cells and human bronchial cells clearly demonstrate that sodium selenite represents the opposite effects to diminish telomerase activity (Chen et al., 2007; Zarghami, 2011). However, what are the distinct molecular mechanisms involved in the regulation of telomerase activity in the selenium-induced cells? This remains to be understood. So, in the remaining study we tried to find whether telomerase activity is related to c-Myc and P53 expression levels.
Sodium selenite-exposed hUCMSCs apparently revealed the higher telomerase activity and this function was found to be associated with overexpression of c-Myc and p53 compared to the untreated cells. We also noticed that these results were completely different compared to the NaSel-induced AGS cells in which selenium downregulated telomerase activity and c-Myc and P53, suggesting NaSel inducing ability on telomerase activity is fully dependent on the cell type. SeMet-induced hUCMSCs showed a completely different story so that c-Myc and p53 were underexpressed, and this downregulation was accompanied by reduction in telomerase activity. Hence, these findings were similar to what happened in AGS cells, proposing the idea that organoselenium may show an antitelomerase function, independent of the cell type. Chi-Hwa et al., have reported that c-Myc inactivation—either in vitro or in vivo—results in cellular senescence, suggesting the inactivation of telomerase (Chi-Hwa et al., 2007). However, using an inducible c-Myc overexpression, Drissi et al. have made an opposite system of observations in human leukaemia cells, where c-Myc activation leads to apoptosis, but no alterations in TERT levels or telomerase activity were evident (Drissi et al., 2001). Another study in rat hepatocytes also confirmed our results that NaSel increases the cellular telomerase activity and hTERT/c-Myc/p53 gene expression level (Yu et al., 2009). Moreover, it has been shown that exposure of leukemic cells to antisense c-Myc also resulted in the inhibition of telomerase activity (Fujimoto and Takahashi, 1997).
The telomerase-positive cells should express hTERT as a functional subunit that can be regulated by c-Myc (Wick et al., 1999). To evaluate any relationship between hTERT and c-Myc with telomerase activity, we measured hTERT expression level in selenium-induced AGS and found that both NaSel and SeMet significantly diminished hTERT expression up to 7- and 23-fold in comparison with untreated cells. However, we could not detect any hTERT expression in hUCMSCs in untreated and selenium-exposed cells. No hTERT mRNA expression in hUCMSCs may reflect on the existence of under-detectable amounts of hTERT mRNA in these cells but this level is sufficient for initiation or acceleration of telomerase activity. Using TaqMan-based real-time PCR, Wang et al. found that hTERT was expressed in hUCMSCs at a very low level (Wang et al., 2014).
In conclusion, the telomerase activity is related to hTERT, c-Myc, and p53 expression levels and selenium compounds have different telomerase activity effects on both the hUCMSCs and gastric cancer cells, but the exact molecular signaling transduction pathway promoting these diverse impacts remains to be elucidated further.
Footnotes
Acknowledgments
This study was funded by a grant provided by the Research Council for Ardabil University of Medical Sciences (ARUMS, Iran).
Disclosure Statement
No competing financial interests exist.