
Abstract
Bacterial homologous chloroplast-targeted Obg GTPases (ObgCs) belong to the plant-typical Obg group, which is involved in diverse physiological processes during chloroplast development. However, the evolutionarily conserved function of ObgC in plants remains elusive and requires further investigation. In this study, we identified DoObgC from an epiphytic plant Dendrobium officinale and demonstrated the characteristics of DoObgC. Sequence analysis indicated that DoObgC is highly conserved with other plant ObgCs, which contain the chloroplast transit peptide (cTP), Obg fold, G domain, and OCT regions. The C terminus of DoObgC lacking the chloroplast-targeting cTP region, DoObgCΔ1–160, showed strong similarity to ObgE and other bacterial Obgs. Overexpression of DoObgCΔ1–160 in Escherichia coli caused slow cell growth and an increased number of elongated cells. This phenotype was consistent with the phenotype of cells overexpressing ObgE. Furthermore, the expression of recombinant DoObgCΔ1–160 enhanced the cell persistence of E. coli to streptomycin. Results of transient expression assays revealed that DoObgC was localized to chloroplasts. Moreover, we demonstrated that DoObgC could rescue the embryotic lethal phenotype of the Arabidopsis obgc-t mutant, suggesting that DoObgC is a functional homolog to Arabidopsis AtObgC in D. officinale. Gene expression profiles showed that DoObgC was expressed in leaf-specific and light-dependent patterns and that DoObgC responded to wounding treatments. Our previous and present studies reveal that ObgC has an evolutionarily conserved role in ribosome biogenesis to adapt chloroplast development to the environment.
Introduction
T
For the first time, we identified an essential gene AtObgC, which encodes a chloroplast-localized Obg GTPase (Bang et al., 2009). Garcia's group proposed that AtObgC/CPSAR1 is a prerequisite for the formation of normal thylakoid membranes, implying its role in the initiation of vesicles from the inner envelope membrane for the transfer of lipids to the thylakoids (Garcia et al., 2010). Recently, we found that loss-of-function mutation in ObgC, by affecting plastid rRNA processing, leads to severe suppression of protein biosynthesis. These results clarify the vital role of plant ObgC in the biogenesis of plastid ribosome (Bang et al., 2012). Moreover, AtObgC interacts with AtRSH1 (a plant homolog to the bacterial RelA/SpoT), suggesting that AtObgC may be involved in the response to ppGpp-mediated stress signaling (Chen et al., 2014). Previous research provides the significant insight that plant ObgC may regulate diverse physiological processes during chloroplast development, but the evolutionarily conserved function of ObgC in plants remains elusive.
Dendrobium officinale, which belongs to Dendrobium, one of the largest genera in Orchidaceae, is famous for its unparalleled medicinal value and is ranked as “the top one of the nine Chinese supernatural medicinal herbs” (Xu et al., 2011). The major medicinal components of D. officinale include alkaloids, polysaccharides, amino acids, and several trace mineral elements; and its medical benefits include the stimulation of saliva, inhibition of tumor growth, enhancement of immunity, and improvement in eyesight and stomach functions (Guo et al., 2013; Meng et al., 2013; Pan et al., 2014). Habitat destruction and human overexploitation have gradually destroyed the natural populations of D. officinale, and in 1992, it was classified as an endangered species in the Chinese Plant Red Book (Ding et al., 2009). As an epiphytic or lithophytic plant, poor growth environment, low germination percentage, and slow growth cycle are the major obstacles that D. officinale farming faces (Tan et al., 2014).
To explore the evolutionarily conserved functions of diverse plant ObgCs other than Arabidopsis and rice ObgCs, in this study, we report on the functional analyses of D. officinale ObgC (DoObgC), a chloroplast-localized protein capable of rescuing defects in the Arabidopsis obgc-t mutant. The DoObgCΔ1–160 region showing strong similarity to bacterial Obg was expressed in E. coli cells to investigate the evolutionarily conserved functions of DoObgC. The susceptibility assays of ribosomal antibiotics supported the functional role of DoObgC in ribosome biogenesis. The expression of the DoObg gene showed a leaf-specific and light-dependent pattern similar to the expression pattern of AtObgC in response to wounding stress. Our results suggest that DoObgC is a functional homolog of Arabidopsis AtObgC, implying that DoObgC may be involved in ribosome biogenesis and chloroplast development in response to environmental cues.
Materials and Methods
Plant materials
We grew D. officinale plants in media composed of barks and rubble (1:1) with a 30°C day/20°C night temperature, a 16-h light/8-h dark cycle, and 75% humidity. Leaves, stems, roots, and flowers were collected from 2-year-old plants and frozen in liquid nitrogen. Protocorms were cultured on MS agar media containing 0.5 mg/L of 6-BA, 0.1 mg/L of NAA, 0.2 g/L of activated carbon, and 100 mg/L of inositol at 25°C under 16-h light/8-h dark condition (Lin et al., 2011). Aseptic seedlings were harvested after a 6-month subculture.
Prof. Jeong Dong Bahk (Gyeongsang National University, Jinju, South Korea) generously provided the Arabidopsis T-DNA mutant (obgc1-t) (Bang et al., 2009).
Cloning of DoObgC gene from D. officinale
In accordance with the cDNA sequence of AtObgC, we searched for the fragment containing DoObgC gene in the D. officinale genomic DNA database (Chinese Herbal Plant Genome Database,
In accordance with the manufacturer's (TransGen Biotech,
Expression of GST-DoObgC in E. coli and cell growth analysis
We amplified the fragment of DoObgC encoding DoObgCΔ1–160 protein using gene-specific primers flanked by restriction sites of BamH I and Xho I in the forward and reverse primers, respectively. After PCR purification and digestion, we cloned the digested PCR product into the pGEX-6p-1 vector. We used the E. coli BL21 strain, transformed with the resulting plasmid, to produce the GST-DoObgCΔ1–160 fusion protein. We performed protein induction by adding isopropyl β-
We grew the BL21 cells with or without pGEX-DoObgC Δ1–160 at 37°C until OD600 reached 0.5. Cell cultures were serially diluted (10−3, 10−4, 10−5, and 10−6). Two microliters of each concentration of cells were spotted on an LB plate with IPTG at varying concentrations (0, 0.025, 0.05, 0.075, and 0.1 mM) and incubated at 37°C for 12 h (Feng et al., 2014).
Antibiotic susceptibility assays
For the resistance assay, we inoculated the BL21 cells with pGEX-DoObgC Δ1–160 and pGEX-6p-1 empty vector, respectively, in test tubes with 5 mL LB and grew them overnight at 37°C with shaking at 200 rpm. After overnight growth, we transferred 50 μL of the culture to fresh 5 mL LB and grew it at 37°C until OD600 reached ∼0.5. For the efficient expression of GST-DoObgCΔ1–160 and GST proteins, we transferred the cultures to a 30°C incubator and induced them with 0.1 mM IPTG for 3 h, and then treated them by adding a stock solution of streptomycin in test tubes to achieve a final concentration of 25 μg/mL. We also performed the control treatment with sterile water on the cultures. We performed all streptomycin and control treatments at 37°C with shaking at 200 rpm during 5 h. To determine the colony-forming units (CFUs), we harvested 1 mL aliquots, resuspended them in a fresh medium, serially diluted them, and plated them on a solid LB medium. We calculated the survival ratio by dividing CFU/mL of the cells treated with streptomycin by CFU/mL of cells without streptomycin.
To monitor the cell growth of E. coli and the inhibition by streptomycin, we measured the absorbance of the resulting solution at 600 nm with a Multiskan spectrum microplate spectrophotometer (Thermo Scientific, Vantaa, Finland). We grew the cells with pGEX-DoObgC Δ1–160 or pGEX-6p-1 empty vector overnight in the LB medium at 37°C, shaking at 200 rpm. For treatment, we diluted the overnight cultures 100-fold in the LB medium and used them to inoculate 200 μL of LB medium containing 0.1 mM IPTG in a 96-well microtiter plate at an initial absorbance OD600 of 0.08. All strains were grown in triplicate with and without 25 μg/mL streptomycin. We incubated samples at 37°C with shaking at 120 rpm and recorded the OD600 by spectrophotometer for growth curves. For the confirmation of the specific streptomycin resistance phenotype, we also performed the treatment with chloramphenicol at various concentrations (0, 1, 5, 25, and 125 μg/mL). We incubated the samples at 37°C with shaking at 120 rpm and recorded the OD600 after a 4-h incubation. The growth levels of the chloramphenicol-treated cells were normalized to those of untreated cells, which we used as control.
Subcellular localization of DoObgC-YFP by Agrobacterium-mediated transient assay
Stop codon was eliminated from DoObgC open reading frame (ORF) during amplification using gene-specific primers for cloning into donor vector pENTR/D-TOPO (Invitrogen). Following the manufacturer's instructions, using Gateway LR reactions (Invitrogen), we transferred the resulting clone into destination vector pEarlygate101 to produce an ORF containing DoObgC and YFP. After sequence verification, the plasmid was transformed into Agrobacterium tumefaciens LBA4404. We used 6-week-old Nicotiana benthamiana plants for Agrobacterium-mediated transformation as described (Erickson et al., 2014). We observed expression of the fusion proteins 2 and 3 days after transformation using a Nikon A1R/A1 confocal laser scanning microscope (Nikon Corporation, Tokyo, Japan). Excitation/emission wavelengths were 488/485–520 nm for YFP and 561/600–700 nm for intrinsic fluorescence of chloroplasts. The images shown in results are single focal sections that we acquired using Nikon Elements software.
Complementation of obgc-t mutant with DoObgC
Following the manufacturer's instructions, using Gateway LR reactions (Invitrogen), we transferred the pENTR/D-DoObgC construct into destination vector pMDC83. We introduced the resulting construct into A. tumefaciens and used it to transform the heterozygous obgc-t Arabidopsis line by flower dipping. We selected Hygromycin-resistant T2 transformants and further identified the homozygous/heterozygous/wild-type obgc-t lines carrying DoObgC-GFP by PCR analysis of the T-DNA/AtObgC junction, as described previously (Bang et al., 2009).
Quantitative real-time polymerase chain reaction analysis of DoObgC expression
Using TRIzol reagent (Invitrogen), we extracted total RNA from D. officinale. We used the ReverTra Ace-α-® kit (TOYOBO,
Results
Molecular cloning of the full-length DoObgC cDNA
To identify potential D. officinale sequences homologous to ObgC with the conserved domains, we used ObgC homologous sequence from Arabidopsis (AtObgC) as the query sequence and searched the D. officinale genome database. We retrieved a unique 7305-bp-length genome sequence homologous to AtObgC. However, this genome sequence covered only partial CDSs of ObgC. We expanded this 7 kb fragment by 5 kb both up- and downstream. By comparing the protein sequence of AtObgC with the obtained 17-kb-length genomic DNA sequence, we used GeneWise to predict six exons of the putative DoObgC gene. However, no start codon or stop codon was found in the six-exon constituent CDS region.
To isolate the full-length cDNA of DoObgC, we carried out 5′-RACE and 3′-RACE experiments based on the partial CDS. Using the cDNA synthesized from RNAs extracted from the leaves of 6-month-old D. officinale seedlings, we designed primers from the predicted DoObgC CDS and successfully amplified a 1563-bp potential DoObgC fragment. Next, based on this fragment, we obtained the 5′ and 3′ sequences of DoObgC using the RACE strategy and confirmed that the full-length DoObgC cDNA was 1993 bp in length (Supplementary Fig. S1). We searched the ORF of DoObgC using ORF Finder (
Bioinformatics analysis of the DoObgC protein
To analyze the evolutionary relationship of DoObgC to plant ObgCs, we searched the NCBI databases using DoObgC protein as the query sequence and found many Obg homologs from prokaryota to plantae. From among these proteins, we selected three well-known bacterial Obgs and 18 typical ObgC homologs of other plant species to construct the phylogenetic tree of Obg proteins (Fig. 1). The phylogenetic analysis showed that these plant ObgCs could be divided into three families from Chlorophyta to Mesangiospermae, which consists of two groups, Monocotyledonae and Dicotyledonae. As shown in Figure 1, the DoObgC protein belongs to the Monocotyledonae group of the Mesangiospermae ObgC family.
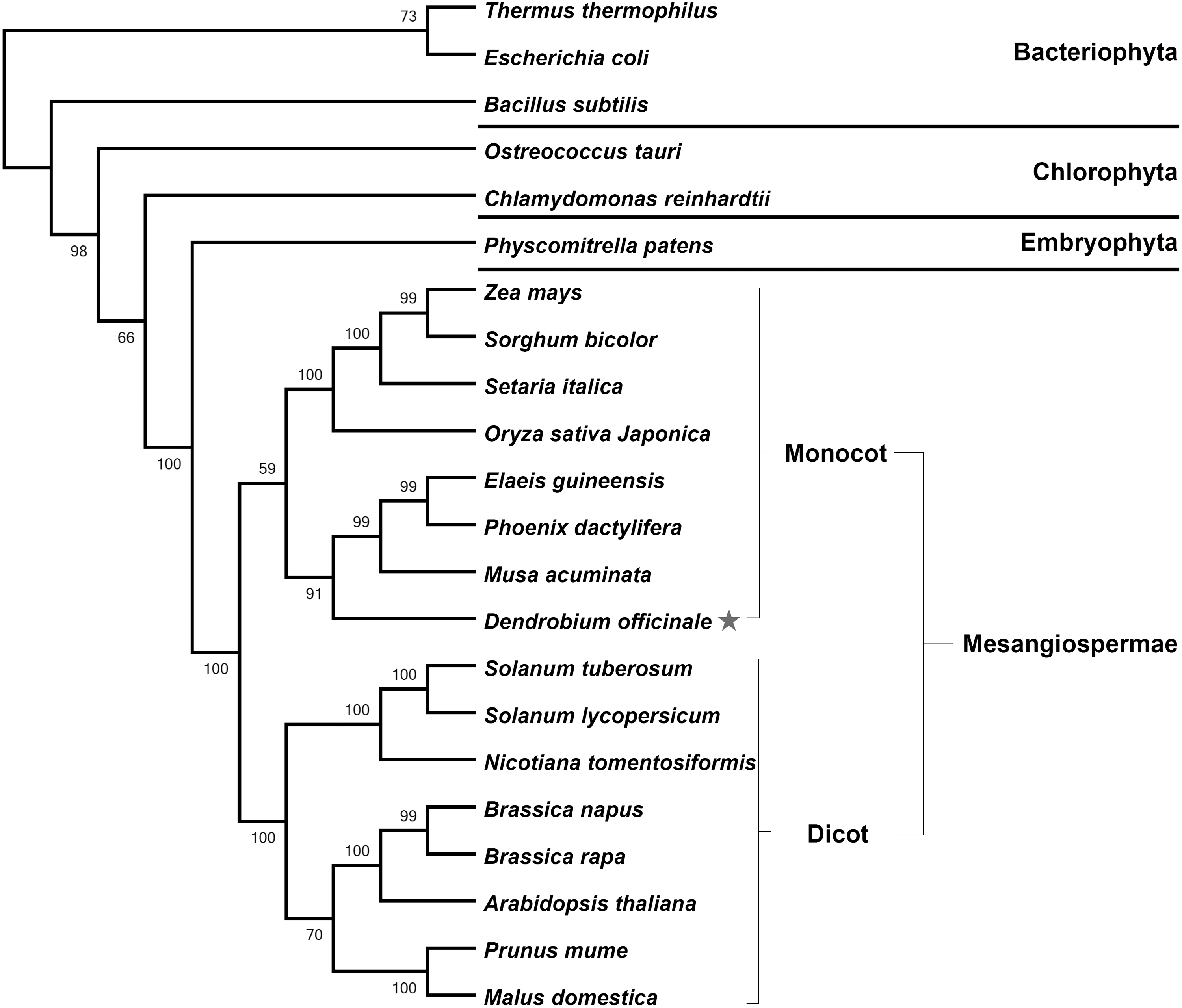
Phylogenetic tree of Obg proteins from different organisms. GenBank accession numbers: Thermus thermophilus (YP_145047); Escherichia coli (NC_000913); Bacillus subtilis (NP_390670); Ostreococcus tauri (XP_003078221); Chlamydomonas reinhardtii (XP_001702482); Physcomitrella patens (XP_001751788); Zea mays (DAA41697); Sorghum bicolor (XP_002461174); Setaria italica (XP_004958653); Oryza sativa Japonica (NP_001060586); Elaeis guineensis (XP_010921754); Phoenix dactylifera (XP_008778460); Musa acuminata (XP_009417935); Solanum tuberosum (XP_006342758); Solanum lycopersicum (XP_010321129); Nicotiana tomentosiformis (XP_009588280); Brassica napus (CDY58104); Brassica rapa (XP_009126267); Arabidopsis thaliana (NP_197358); Prunus mume (XP_008223125); Malus domestica (XP_008340828). Asterisk indicates ObgC protein from Dendrobium officinale. The phylogenetic tree was constructed using the neighbor-joining method and was tested by 1000 replications of the bootstrap analysis, which was visualized using MEGA5 software.
To predict physical and chemical characteristics of the DoObgC protein, we used ExPASy Bioinformatics-Proteomics Server online software (

Amino acid sequence alignment of DoObgC with other Obgs. Amino acids shaded in black indicate identical residues, while those shaded in gray are similar residues. The sequences compared are from two Obg groups, plant-specific ObgCs, including A. thaliana ObgC (AtObgC, NP_197358), D. officinale ObgC (DoObgC, KT359612), and Oryza sativa Japonica ObgC1 (OsObgC1, NP_001060586), and bacterial Obg proteins, including T. thermophilus Obg (TtObg, YP_145047), B. subtilis Obg (BsObg, NP_390670), and E. coli Obg (ObgE, NC_000913). The N-terminal chloroplast transit peptides are underlined with gray solid line, and the conserved Obg domains, which include Obg fold, GTPase, and OCT, are indicated using dashed line, black line, and dotted line, respectively.
The effect of DoObgCΔ1–160 expression on cell growth
Bacterial Obg proteins are universally proposed with molecular function in ribosome assembly (Feng et al., 2014). Moreover, the region of DoObgC (161–623 residues) shows strong similarity to ObgE and other bacterial Obgs (Fig. 2). We therefore hypothesized that DoObgC could have a similar function involved in ribosome assembly with ObgE. To assess the role of DoObgC protein in ribosome biogenesis, we cloned the fragment of DoObgC encoding DoObgCΔ1–160 protein (50 kDa) into the pGEX-6p-1 vector and then obtained the E. coli BL21 strains allowing controlled expression of the recombinant GST-tagged DoObgCΔ1–160. Using SDS-PAGE, we evaluated the size of DoObgCΔ1–160 and GST proteins, respectively, as ∼75 and 25 kDa (Supplementary Fig. S2). In addition, overexpression of DoObgCΔ1–160 seemed to slow down the growth of E. coli cells. We further tested the effect of DoObgCΔ1–160 expression in cell growth, performing the serial dilution-spotting assay as previously described (Feng et al., 2014). As Figure 3 illustrates, the growth of the GST-DoObgCΔ1–160-expressed cells was slower when compared with that of GST expressed under the 0.05–0.1 mM IPTG induction. Interestingly, 0.025 mM (Fig. 3) or less IPTG did not inhibit cell growth (data not shown).
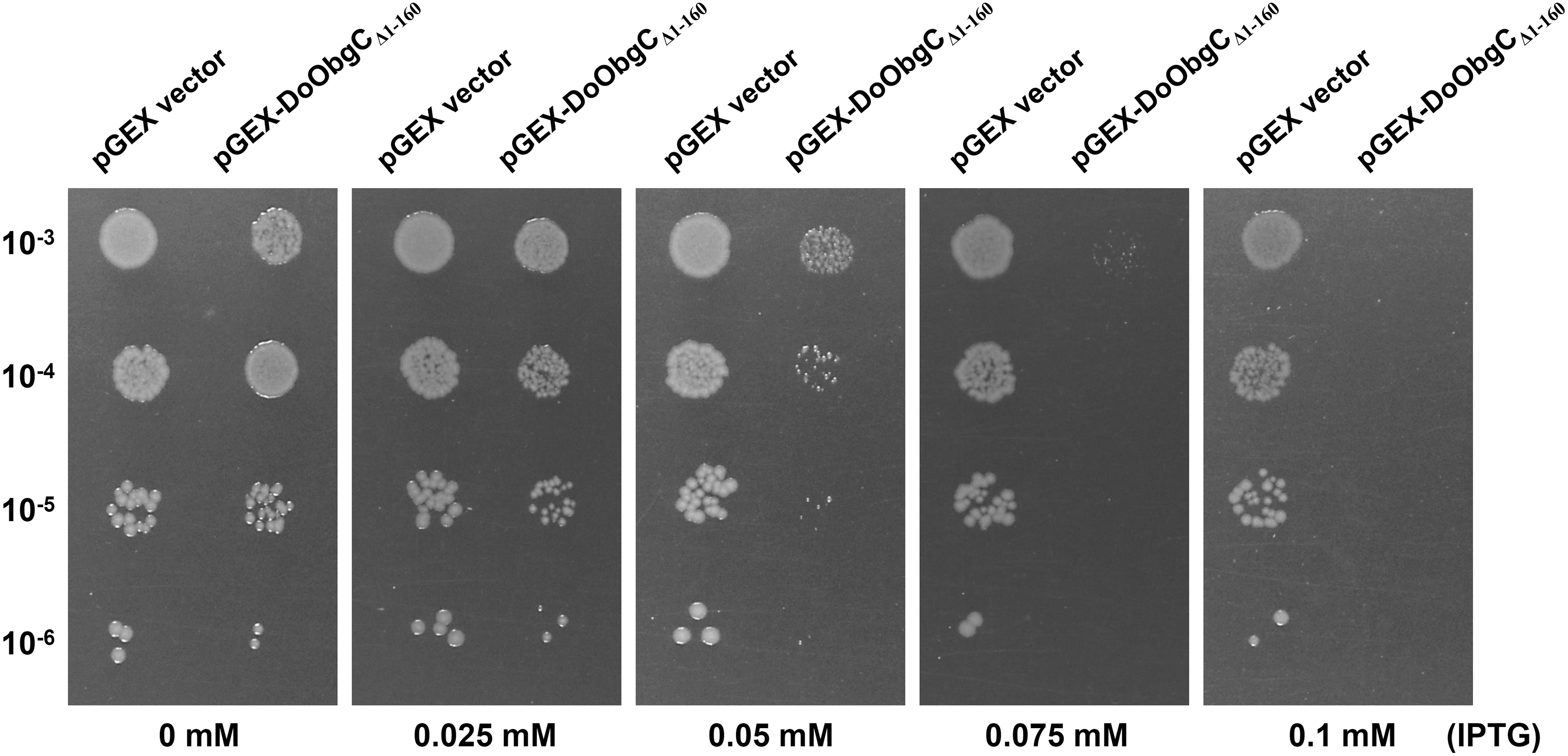
The effect of overexpression of DoObgCΔ1–160 in E. coli on the cell growth. The E. coli BL21 strains with empty pGEX-6p-1 vector or pGEX-DoObgC
Δ1–160 were serially diluted (10−3, 10−4, 10−5 and 10−6) on the LB plates with IPTG in the incremental concentrations (0, 0.025, 0.05, 0.075, and 0.1 mM). Cell cultures were incubated at 37°C for 12 h. IPTG, isopropyl β-
The common features of overexpressing bacterial Obg in E. coli are slower growth and elongated cells (Dutkiewicz et al., 2002; Polkinghorne and Vaughan, 2011). To test whether overexpression of DoObgCΔ1–160 leads to changes in cell morphology, we examined cultures with empty pGEX-6p-1 vector or pGEX-DoObgC Δ1–160 incubated with 0.1 mM IPTG for 3 h by microscopy (Supplementary Fig. S3). The result revealed a clear phenotypic effect in DoObgCΔ1–160 overexpressing bacteria with ∼40% of cells displaying an elongated phenotype (Supplementary Fig. S3B). We did not observe this phenotype in GST overexpressing controls (Supplementary Fig. S3A).
Because ObgE reportedly prevents the association of the ribosomal subunits by binding to the 50S subunit, the overexpression of ObgE in E. coli causes a slow cell growth (Feng et al., 2014). This evidence implies that the overexpression of DoObgCΔ1–160 may lead to the dissociation of ribosomes.
The effect of DoObgCΔ1–160 expression on susceptibility to ribosomal antibiotics
Streptomycin, one of the most commonly used aminoglycosides, specifically binds to the 30S ribosomal subunit and then induces breakdown of the completed 70S initiation complex on ribosomes of E. coli (Modolell and Davis, 1970; Demirci et al., 2013). To examine the effect of DoObgCΔ1–160 overexpression on the cell's susceptibility to ribosomal antibiotics, we treated the exponential phase cultures of the GST and GST-DoObgCΔ1–160 with 25 μg/mL streptomycin for 5 h, respectively. The GST-DoObgCΔ1–160-expressed cells showed much higher survival percentage in comparison with the GST-expressed cells (sixfold, Fig. 4A). Examining various concentrations of streptomycin showed that the overexpression of GST-DoObgCΔ1–160 exhibited insusceptibility to streptomycin (data not shown) in comparison with the overexpression of GST proteins. To further outline how streptomycin influences cell growth upon the expression of DoObgCΔ1–160, we measured the cell densities of untreated samples and samples treated with streptomycin with a microplate spectrophotometer. In the control cells with GST expression, the manifest lag period occurred on growth rates with streptomycin treatment (“X” and “O”, Fig. 4B). However, the treatment of streptomycin had no effect during the exponential phase of the expressed GST-DoObgCΔ1–160 cells (dark gray “−” and light gray “+,” Fig. 4B). Furthermore, GST-DoObgCΔ1–160-overexpressed cells exhibited mild resistance to streptomycin in the stationary phase in comparison to the control cells (Fig. 4B). These results suggested that the expression of DoObgCΔ1–160 antagonizes the growth-inhibitory effect of streptomycin in the early period of the growth cycle. This led us to explore other ribosomal antibiotics, such as translation inhibitor chloramphenicol (Schlunzen et al., 2001). Unexpectedly, the cell expressing GST-DoObgCΔ1–160 in the exponential phase was even more sensitive to various concentrations of chloramphenicol in comparison with GST-expressed cells (Supplementary Fig. S4). Thus, overexpression of DoObgCΔ1–160 specifically enhances cell insusceptibility to streptomycin, but not to chloramphenicol.
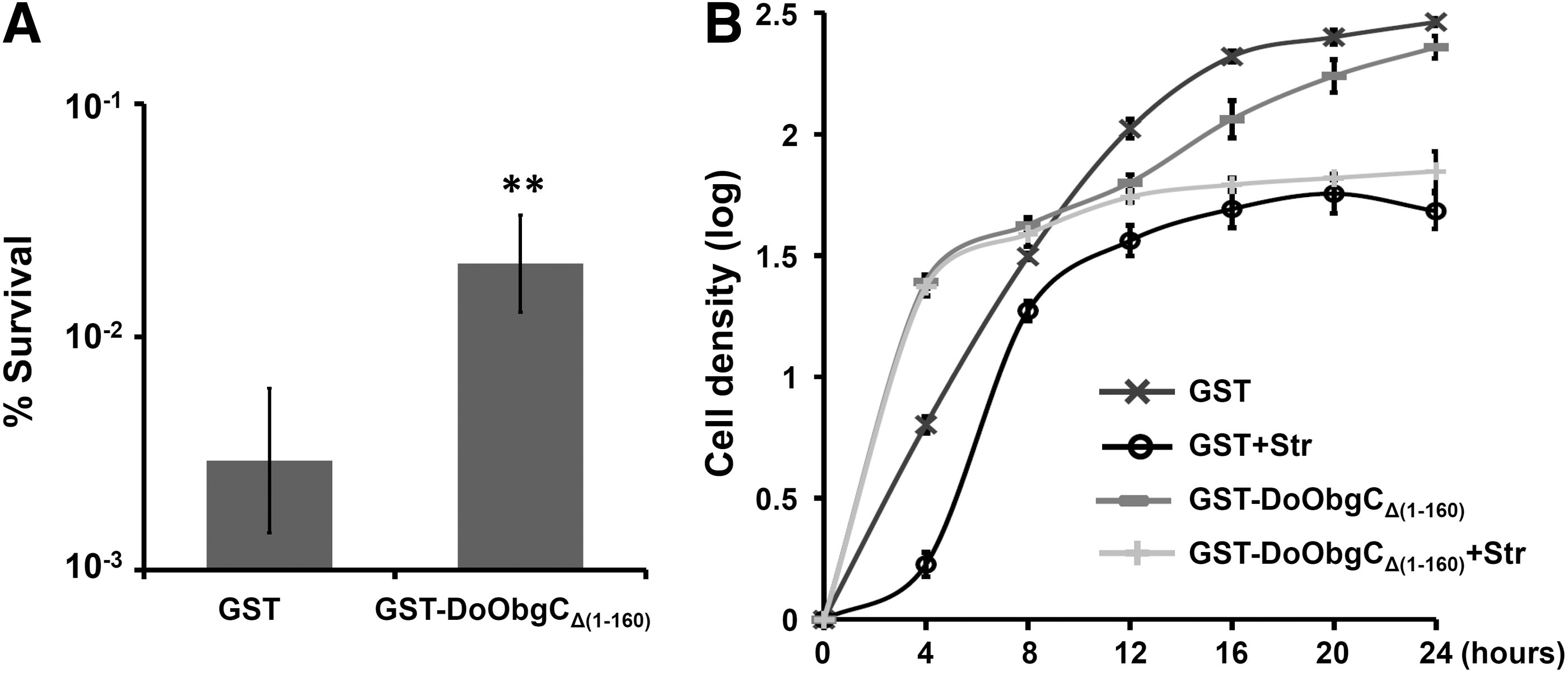
Growth assays of DoObgCΔ1–160-expressing E. coli treated with streptomycin.
Subcellular localization of DoObgC
To verify the predicted localization of DoObgC in the chloroplast, we used an agrobacterium-mediated transformation method to transform into N. benthamiana a construct containing the full-length DoObgC attached to YFP under the control of a 35S promoter (35S-DoObgC-YFP). We used confocal laser-scanning microscopy to examine the fluorescence of the DoObgC-YFP fusion protein. As Figure 5 shows, the green signals of DoObgC-YFP overlapped with the red chlorophyll autofluorescence (CHL), suggesting that DoObgC is specifically targeted to the chloroplast. Like the punctate pattern of AtObgC-GFP (Bang et al., 2009), DoObgC-YFP proteins clearly appeared as small spots in the chloroplast (Fig. 5).

Subcellular localization of DoObgC-YFP. Localization of DoObgC-YFP was examined in tobacco using Agrobacteria-mediated transient assay. Twenty-four hours after infiltration, transformed tobacco plants were observed under confocal microscope. Scale bar in the image represents 20 μm in length. “Merge” displays the overlapped images of Bright, YFP, and CHL (chlorophyll autofluorescence), which was used as a marker for chloroplast distribution.
Functional complementation of DoObgC in Arabidopsis
Disruption of the AtObgC gene, an important gene for the biogenesis of chloroplast ribosomes, leads to seed abortion at the early stage of embryogenesis (Bang et al., 2012). To further validate whether DoObgC has conserved functions with AtObgC in planta, the DoObgC gene was ectopically expressed under the control of CaMV 35S promoter in Arabidopsis obgc-t mutants (Table 1). For genotype segregation analysis, we used T2 plants that could survive hygromycin. We determined genotype segregation of heterozygote Arabidopsis obgc mutants transformed with 35S-DoObgC-GFP through PCR genotyping (Bang et al., 2009). The results revealed that the ratio of wild-type:heterozygous:homozygous plants was not significantly different from the expected Mendelian segregation ratio of 1:2:1, as assessed using the chi-squared test (χ 2 = 4.2539, df = 2, p = 0.1192) (Table 1). Subsequently, we obtained survival homozygous atobgc mutants from segregated T2 plants of transformed heterozygous obgc-t plants with 35S-DoObgC-GFP construct. The overexpression of DoObgC suppressed the embryonic lethality caused by obgc-t mutation, demonstrating that DoObgC is a functional homolog of AtObgC. The complemented plants showed normal morphology and seed viability.
T2 numbers indicate segregated wild type (WT), heterozygous (hetero), and homozygous (Homo) T2 plants identified by PCR-mediated genotyping.
Total number of T2 plants containing 35S-DoObgC-GFP that can survive hygromycin selection.
Chi-squared test was used to test the Mendelian segregation ratio of 1:2:1 (WT:Hetero:Homo).
Expression patterns of DoObgC in D. officinale
To investigate the tissue-specific expression patterns of DoObgC in D. officinale, we performed qRT-PCR analysis of the gene expressions in different tissues, such as stems, flowers, aerial roots, and leaves (Fig. 6). In our previous study, a northern-blot analysis revealed that AtObgC and OsObgC1 genes exhibited light-dependent and leaf-specific expression patterns (Bang et al., 2009). In accordance with this result, the qRT-PCR analysis indicated that the DoObgC gene was also predominantly expressed in leaves, but was expressed much less in stems, flowers, and roots (Fig. 6). To assess the light-induced expression of DoObgC genes, we used the total RNA purified from the light-grown and dark-grown D. officinale for qRT-PCR analysis. We illuminated the etiolated protocorms of dark-grown D. officinale for 1 month or continuously cultured them in dark conditions, respectively. The dark-grown protocorm-like bodies (PLBs) exhibited a severe chlorotic phenotype, while the light-grown PLBs showed almost normal green color (Fig. 7A). In addition, the DoObgC gene exhibited a light-dependent expression pattern (Fig. 7B), implying that DoObgC may play a role in the light-dependent chloroplast development.
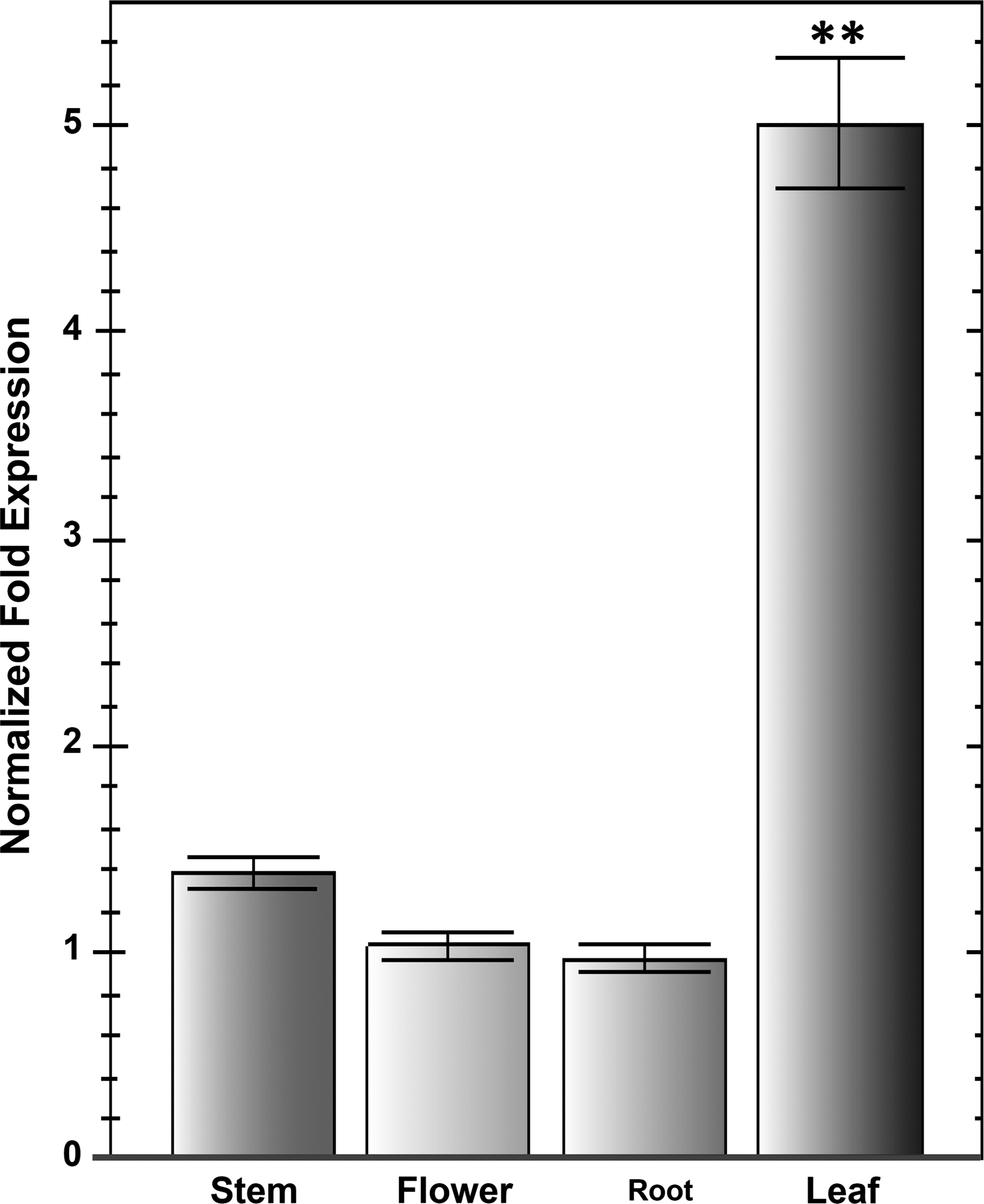
Expression patterns of DoObgC in various tissues. Total RNAs were purified from the following tissues: stems, flowers, roots, and leaves from 2-year-old D. officinale at the reproductive stage. Each transcript level was normalized to GAPDH, which was used as the internal control. Each bar represents mean ± SD values calculated from three biological repeats. Asterisks indicate significant differences of the mean analyzed by Student's t-test (**p ≤ 0.01).
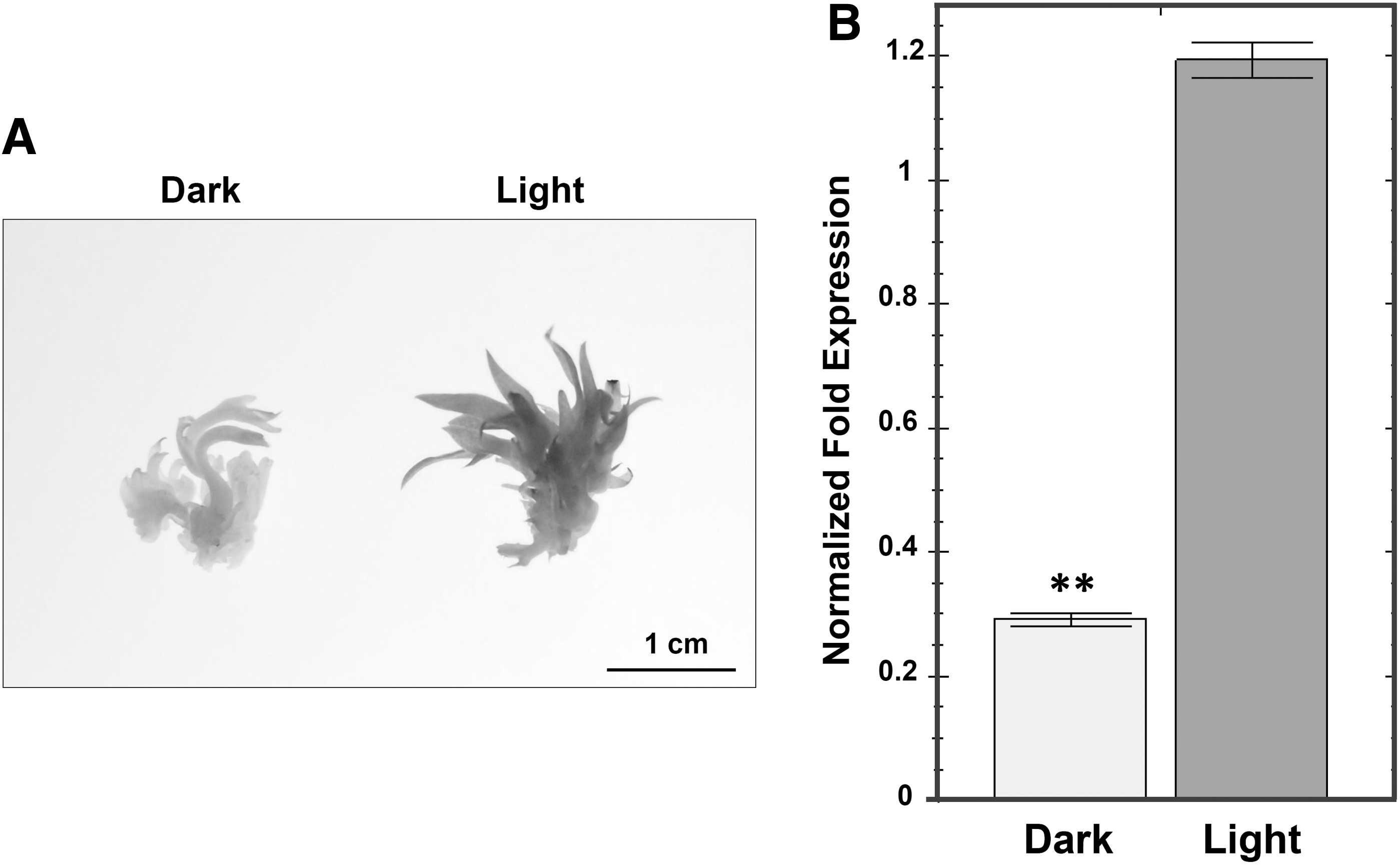
Morphology of D. officinale protocorms and expression pattern of DoObgC in light/dark condition.
Wounding induces high ppGpp accumulation in Arabidopsis chloroplasts (Takahashi et al., 2004). AtObgC has been reported to interact with the putative ppGpp hydrolase AtRSH1, and wounding treatment reduces the expression level of AtObgC, implying that AtObgC may be involved in response to ppGpp-mediated stress signal transduction (Chen et al., 2014). We further analyzed the expression pattern of DoObgC in response to wounding stress. The results showed that the DoObgC mRNA level decreased within 1 h after wounding, as Figure 8 shows. Notably, at 3 h, transcript levels of DoObgC decreased to 53% in comparison with those of untreated plants (0 h; Fig. 8). The expression of DoObgC gene then gradually increased at 6 h and further recovered to almost the transcription level before wounding at 12 h (Fig. 8). The results indicate that DoObgC transcripts exhibit an expression pattern similar to that of AtObgC in response to wounding stress.
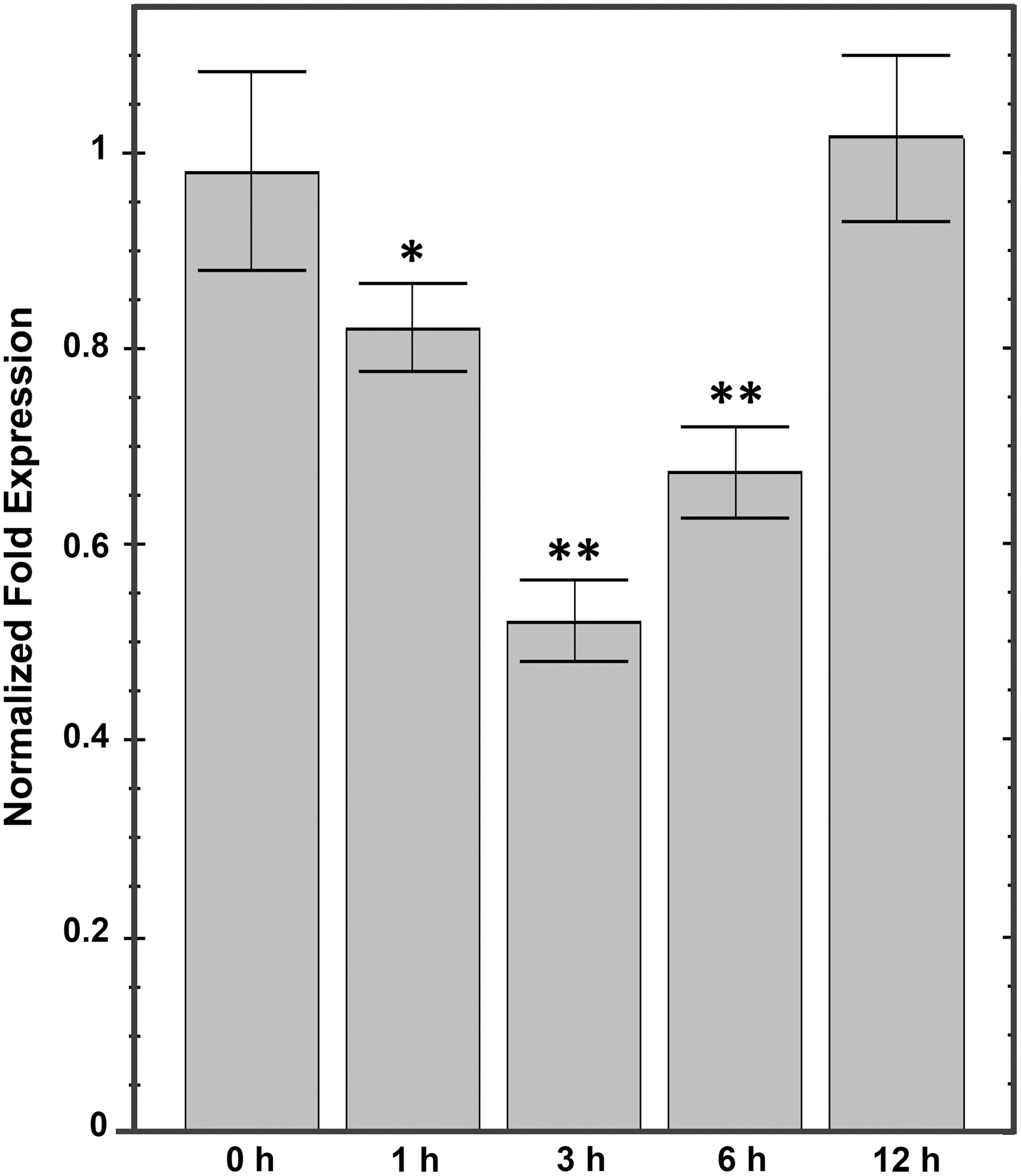
Wounding-induced expression pattern of DoObgC in leaves. Six-month-old D. officinale seedlings were wounded by crushing the leaves with forceps; and leaf samples were harvested 1, 3, 6, and 12 h after the treatment. Leaves without wounding treatment (0 h) were used as control. Each transcript level was normalized to GAPDH, which was used as the internal control. Each bar represents mean ± SD values calculated from three biological repeats. Asterisks indicate significant differences of the mean analyzed by Student's t-test (*p ≤ 0.05, **p ≤ 0.01).
Discussion
Bacterial Obg proteins are linked to multiple functions ranging from sporulation, cell cycle, ribosome biogenesis, and DNA replication to ppGpp-mediated stress response (Kint et al., 2014). Recently, structural and functional analyses of ObgE indicate that ObgE functions as an antiassociation factor, which binds to the 50S subunit to block the formation of the 70S ribosome, senses the cellular energy stress by ppGpp level, and links ribosome biogenesis to stress response machinery (Feng et al., 2014). To test the potential function of D. officinale ObgC in ribosome, the highly conserved region of DoObgC with ObgE protein was heterologously overexpressed in E. coli. Analysis of E. coli overexpressing recombinant DoObgCΔ1–160 revealed inhibition of growth in an IPTG dose-dependent manner (Fig. 3). A similar phenotype has been described for heterologous expression in E. coli of Obg homologs from Vibrio cholerae and Chlamydia abortus (Shah et al., 2008; Polkinghorne and Vaughan, 2011). This result is also consistent with the growth defects of ObgE-overexpressing E. coli cells, suggesting that DoObgCΔ1–160 has the role conserved with ObgE. Overexpression of recombinant DoObgCΔ1–160 also resulted in the enhancement of cell insusceptibility to streptomycin, an inhibitor of bacterial 70S-initiation complex (Fig. 4A). Intriguingly, the expressing DoObgCΔ1–160 strain drastically conferred resistance to streptomycin in the exponential phase, whereas it caused only mild resistance to streptomycin in the stationary phase (Fig. 4B), implying that growth conditions would influence the cell persistence of the expressing DoObgCΔ1–160 to streptomycin. Surprisingly, the cell-expressed DoObgCΔ(1–160) was even more sensitive to another antibiotic chloramphenicol (Supplementary Fig. S4). Chloramphenicol prevents protein chain elongation by inhibiting the peptidyl transferase activity of the bacterial ribosome. In addition, chloramphenicol can reduce the GTPase activity of HflX, a ribosome splitting factor in E. coli (Coatham et al., 2016). Therefore, it is possible that a similar mechanism carries out the inhibition activity of DoObgCΔ(1–160).
The plant-unique Obg homologs, ObgCs from rice and Arabidopsis, are essential for plant vitality and chloroplast development (Bang et al., 2012). However, the evolutionarily conserved functions of diverse plant ObgCs require further investigation. D. officinale is a traditional Chinese orchid herb that has a concomitant CAM and C3 pathways for the adaption to changing environmental conditions (Zhang et al., 2014). Several expanded gene families involved in abiotic and biotic stress responses have been identified in the D. officinale genome (Yan et al., 2015). To understand the fundamental roles of DoObgC in D. officinale, we demonstrated the characteristics of DoObgC in planta. Expression of DoObgC-YFP fusion proteins in tobacco demonstrated that DoObgC-YFP is also targeted to chloroplasts and exhibits the AtObgC-like puncate pattern (Fig. 5).
In addition, DoObgC complemented the embryo lethal phenotype in the Arabidopsis obgc-t mutant (Table 1), further validating that DoObgC is a functional AtObgC homolog. Previous investigations of AtObgC expression in Arabidopsis and OsObgC1 expression in rice exhibited leaf-specific and light-dependent patterns, which are also observable in DoObgC (Figs. 6 and 7). Interestingly, DoObgC transcripts exhibit an expression pattern similar to that of AtObgC in response to wounding stress (Fig. 8), which induces the high ppGpp accumulation in chloroplasts (Takahashi et al., 2004). Based on previous studies, AtObgC plays vital roles in diverse physiological processes during chloroplast development such as plant viability, thylakoid biogenesis, and ribosome biogenesis (Bang et al., 2009, 2012; Garcia et al., 2010). The diverse functions of plant ObgC proteins indicate that this protein family might participate in certain essential processes central to various cellular functions. Thus, we propose that the evolutionarily conserved function of plant ObgC is involved both in ribosome biogenesis and in adapting chloroplast development to the environment. Further studies are required to determine the exact role of DoObgC in plastid ribosome of D. officinale under preferred growth conditions or during wounding stress.
Footnotes
Acknowledgments
We thank Prof. Wen Wang and Dr. Yang Dong (Kunming Institute of Zoology, Kunming, China) for assistance in searching for the D. officinale ObgC sequence. This work was supported by grants from the Education Department of Sichuan Province (No. 14ZB0013) and the National Science Foundation (No. 31501242).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.