
Abstract
Tuberculosis, an infectious disease caused by Mycobacterium tuberculosis (MTB), is one of the global public health catastrophes. Wnt signaling has recently been identified to exert immunoregulatory functions in a variety of inflammatory and infectious diseases, including tuberculosis. The opposite expression of Wnt5a in human and mice during MTB infection drives us to explore the roles and biological significances of reduced Wnt5a for MTB-treated mice. In our study, the reduction of WNT5A in MTB-treated mice lung tissues or MTB-infected mice bone marrow-derived macrophages (BM-Mø) was in a dose- and time-dependent manner. Then, WNT5A-silenced mice, secreted frizzled-related protein 1 (SFRP1)-overexpressed or -silenced mice BM-Mø, were constructed to regulate Wnt5a levels. When Wnt5a is deficient, MTB-induced increases of pro-inflammatory cytokines (TNF-α, IL-1β, IL-12, and IL-6) can be markedly attenuated in mice lung tissues or BM-Mø. Besides, external disturbance triggered that Wnt5a lower expression can induce Mø to be M2 phenotype and enhance cell apoptosis of MTB-infected mice BM-Mø. Hence, the reduction of Wnt5a is a tactful strategy adopted by Mø to resistant MTB-induced immune responses and to enhance MTB-induced Mø apoptosis in mice. Our study revealed a new style for Mø to manipulate themselves against MTB infection. Our research identifies that Wnt5a deficiency can regulate inflammatory cytokine secretion, polarization, and apoptosis in MTB-infected Mø.
Introduction
T
It is well demonstrated that Mø play a pivotal role in inflammation by releasing of and responding to numerous inflammatory cytokines and chemokines (Pereira et al., 2008). Within Mø, MTB is able to stably multiply and can efficiently induce Mø apoptosis, which is associated with killing of intracellular MTB (Molloy et al., 1994; Oddo et al., 1998). Hence, it seems that inhibiting apoptosis of Mø might be an effective survival strategy for virulent MTB (Kremer et al., 1997). The immune response of Mø to MTB infections has been sufficiently studied in great detail. On the contrary, still little is known about how Mø manipulate themselves to resistant MTB-induced immune responses and apoptosis.
Wnt signaling pathways are well known as a group of highly conserved principles among metazoan species, which are responsible for essential biological processes in embryogenesis and tissue homeostasis (Angers and Moon, 2009; van Amerongen and Nusse, 2009). Wnt family proteins regulate multiple cellular functions, including proliferation, apoptosis, migration, survival, differentiation, and polarity, by activating various intracellular signaling cascades (Kikuchi et al., 2009). In essence, Wnts have also been identified to exert immunoregulatory functions in a variety of inflammatory and infectious disease, including tuberculosis (Schaale et al., 2013).
Currently, there are at least 19 Wnt members that have been revealed to be present in human and mice (Kikuchi et al., 2009). Among which, Wnt5a, negatively regulated by secreted frizzled-related protein 1 (Sfrp1) (Surinkaew et al., 2015), was reported to be the first member of Wnt family (Schaale et al., 2013). Generally, Wnt5a was induced in multiple inflammatory diseases, such as sepsis (Pereira et al., 2008), atherosclerosis (Christman et al., 2008), and tuberculosis (Blumenthal et al., 2006), and commonly exert pro-inflammatory effects on MTB-infected human Mø (Blumenthal et al., 2006; Schaale et al., 2011). While in mice, Reiling and colleagues discovered that Wnt5a was markedly reduced in the lung tissue of MTB-treated mice, as well as in MTB-infected mice bone marrow-derived macrophages (BM-Mø) (Schaale et al., 2013), which aroused our interest in the roles and biological significances of reduced Wnt5a in MTB-infected mice BM-Mø.
In the present study, the reducing of WNT5A in MTB-treated mice or MTB-infected mice BM-Mø was verified first. Through silencing or inhibiting of WNT5A gene factitiously, we found that MTB infection, which induced increases of inflammatory cytokine secretion, can be attenuated in Wnt5a lacked mice or mice BM-Mø and enhanced in Wnt5a overexpressed mice BM-Mø. Besides, when Wnt5a was insufficient, Mø phenotype trended to be M2 and cell apoptosis was significantly enhanced in MTB-infected BM-Mø. These findings hinted us that, in mice, Wnt5a deficiency can lead to a resistance of Mø to MTB-induced immune responses and reinforcement of MTB-induced Mø apoptosis.
Materials and Methods
Mycobacteria preparation
M.tuberculosis H37Rv was purchased from American Type Culture Collection (ATCC, Manassas, VA). Experiments were performed under biosafety level 3 (BSL3) conditions. H37Rv was cultured in Middlebrook 7H9 broth (Difco, Detroit, MI) containing 10% oleic acid albumin-dextrose-catalase (OADC; Difco), 0.05% Tween 80 (Sigma-Aldrich, St. Louis, MO), and 0.2% glycerol (Sigma) at 37°C in a shaking incubator until log phase (OD600∼0.6) (Venketaraman et al., 2005; Chandolia et al., 2014). Before use, H37Rv was dispersed by aspiration through a 29-gauge needle (BD Biosciences, Franklin Lakes, NJ), vortexed, and then sonicated for 15 s in a bath sonicator (100 Watt, KQ-100DB; Kunshan Ultrasound Co., Ltd., China). After sonication, suspensions were allowed to settle for 10 min, and the upper 500 mL were removed and stored for use in following experiments (Schaale et al., 2013). For the determination of the number of colony-forming units (CFUs), 10-fold serial dilutions of the prepared H37Rv strain were plated in triplicate onto Middlebrook 7H11 agar plates (Difco) containing 10% OADC and incubated at 37°C. After incubating for 21 days, CFU was calculated (Rahman et al., 2014).
Treatment of mice with H37Rv by aerosol infection
C57BL/6 mice (8 weeks, 24 ± 0.7 g) were purchased from Shanghai Laboratory Animal Center, Chinese Academy of Sciences (Shanghai, China) and housed under specific pathogen-free conditions in accordance with institutional guidelines. All animal experiments were approved by the Animal Care and Use Committee of Shenzhen Third People's Hospital. For MTB infection, mice were infected with different doses of H37Rv (CFU: 0, 100, 500, and 1000; n = 3 for each group) through the aerosol route as described previously (Reiling et al., 2002). After infection for 7 days, lung tissues of these mice were removed to prove the success of infection by colony enumeration assay (Hölscher et al., 2001). After infection for indicated days (0, 7, 14, or 21 days, n = 3 for each group), lung tissues of these mice were removed for use in following experiments. Experiments were performed under BSL3 conditions.
Construction of Wnt5a siRNA recombinant adenovirus
Wnt5a special and nonspecial (normal-control, NC) siRNA sequences were designed and synthesized by GenePharma Co., Ltd. (Shanghai, China). Sequences were listed as follows: Special: 5′-GAA GCC CAU UGG AAU AUU ATT-3′ (sense) and 5′-UAA UAU UCC AAU GGG CUU CTT-3′ (antisense); Nonspecial: 5′-UUC UCC GAA CGU GUC ACG UUU-3′ (sense) and 5′-ACG UGA CAC GUU CGG AGA AUU-3′ (antisense). The Wnt5a siRNA recombinant adenovirus (Ad-Wnt5a-siRNA) was constructed as previously described (Yang et al., 2014). Briefly, the synthesized siRNA sequences were amplified and subcloned into pAdTrack-cytomegalovirus (CMV) plasmids (Stratagene-Agilent Technologies, Waldbronn, Germany). The obtained pAdTrack-CMV-Wnt5a-siRNA or pAdTrack-CMV-NC plasmids were homologously recombined with pAdEasy-1 (Stratagene) in Escherichia coli strain BJ5183 (Stratagene). Then, the secondarily obtained recombinant plasmids were transfected into HEK-293 cells (ATCC). After amplification and purification, virus titers were measured by the p24 ELISA Kit (Cell Biolabs, Inc., San Diego, CA).
Treatment of mice with recombinant Ad-Wnt5a-siRNA
Through caudal vein injection, mice received 200 μL phosphate buffer saline (PBS), which were used as Mock group (n = 3), 200 μL recombinant Ad-NC (1 × 1010 plaque-forming units, PFUs) as rAd-NC group (n = 3), and 200 μL recombinant Ad-Wnt5a-siRNA (1 × 1010 PFUs) as rAd-Wnt5a-siRNA group (n = 9). Mice without any treatment were used as Control group (n = 3). After 14 days, a secondary injection (200 μL PBS to mice in Mock group, 200 μL recombinant Ad-NC to mice in rAd-NC group, and 200 μL recombinant Ad-Wnt5a-siRNA to mice in rAd-Wnt5a-siRNA group) was performed. After another 14 days, three mice in each group were sacrificed to analyze Wnt5a levels in lung tissues. The rest mice in rAd-Wnt5a-siRNA group (n = 9–3) continuously received H37Rv infection or not (1000 CFU for 21 days) as described above. Meanwhile, normal mice treated with or without H37Rv (1000 CFU for 21 days) were used as appropriate controls, respectively.
Preparation of murine BM-Mø
Murine BM-Mø were generated as previously described (Reiling et al., 2001). First, the femurs of three mice were separated and flushed with HBSS (Sigma). To remove fibroblasts, the obtained cells were cultured at 37°C overnight in DMEM (Sigma) containing 10% heat-inactivated FBS (Sigma), 10 mM HEPES (Sigma), 10 mM glutamine (Sigma), 1 mM sodium pyruvate (Sigma), and 50 ng/mL macrophage colony stimulating factor (M-CSF; Sigma). Then, the adherent cells were collected and cultured in DMEM containing M-CSF (50 ng/mL) for 9 days. Cells were collected to identify the markers (F4/80 and CD11b) of Mø by Flow cytometry. Fluorescein isothiocyanate (FITC)-labeled rat monoclonal anti-mouse F4/80 antibody (Abcam, Cambridge, MA) and allophycocyanin (APC)-labeled rat monoclonal anti-mouse CD11b antibody (Abcam) were used for Mø identification.
Treatment of BM-Mø with H37Rv
In vitro experiments, BM-Mø in 24-well plates were infected with DMEM containing different doses of H37Rv (multiplicities of infection, MOI: 0:1, 3:1, 6:1, or 9:1) for indicated times (0, 4, 8, or 12 h). Then, the plates were gently washed with HBSS (Sigma). After washing, cells were collected, and Wnt5a levels were measured by real-time quantitative polymerase chain reaction (RT-qPCR) and Western blots. For inflammatory cytokine (TNF-α, IL-1β, IL-12, IL-6, Arg-1, MRC-1, and TGF-β) level detection by RT-qPCR, cells were collected at 12 h postinfection with heat-inactivated H37Rv or H37Rv at a MOI of 6:1.
Construction and transfection of Sfrp1 overexpression vector
The cDNA of Sfrp1 was subcloned from total RNA of mice BM-Mø with Pyrobest DNA polymerase (Takara, Kyoto, Japan) and inserted into a pcDNA3.1/V5-HisA vector (Invitrogen, Carlsbad, CA). The obtained pcDNA3.1/V5-HisA-Sfrp1-vectors or pcDNA3.1/V5-HisA-empty-vectors were transfected into BM-Mø in 6-well plates using Lipofectamine 2000 (Sigma). After transfection for 48 h, cells were collected to analyze and verify Wnt5a levels. Meanwhile, pcDNA3.1/V5-HisA-Sfrp1-vector transfected cells were arranged to receive heat-inactivated H37Rv or H37Rv infection with a MOI of 6:1 for 12 h. Then, mRNA levels of inflammatory cytokine (TNF-α, IL-1β, IL-12, IL-6, Arg-1, MRC-1, and TGF-β) in the above cells were analyzed by RT-qPCR.
Silencing of Sfrp1 expression with siRNA interference
For Sfrp1 gene silence, siRNA transfection was performed as per the manufacturer's protocol. In brief, 0.2 nmol mouse Sfrp1 siRNA (Thermo Fisher Scientific, Waltham, MA) or corresponding Silencer® Select Negative Control No. 1 siRNA and 20 μL Lipofectamine 2000 were incubated in 2 mL DMEM at room temperature for 1.5 h. The above mixtures were added to BM-Mø in six-well plates and cultured at 37°C for 48 h. Then, cells were collected to verify Wnt5a levels. Besides, Sfrp1-silenced cells were continuously infected with heat-inactivated H37Rv or H37Rv with a MOI of 6:1 for 12 h. Then, cells were collected to analyze inflammatory cytokine secretion (TNF-α, IL-1β, IL-12, and IL-6).
Real-time quantitative PCR
Total RNA of mice lung tissues or BM-Mø was extracted using the High Pure RNA Kit (Roche, Mannheim, Germany). The purity and integrity of the obtained RNA were controlled by absorbance at A 260/280. For each sample, 4 μg total RNA was reverse transcribed using a High Capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA) in a reaction volume of 30 μL. The acquired cDNA was used for amplification using SYBR Green PCR Amplification Reagent Kit (Qiagen, Hilden, Germany) as per the manufacturer's instructions. RT-qPCR was performed on an Applied Biosystems PRISM 7500 Fast Sequence Detection System (Thermo Fisher Scientific). Primers were synthesized by Shanghai Sangon Biological Engineering Technology and Service Corporation Ltd. (Shanghai, China) and listed in Table 1. All reactions were performed in triplicate. Data were calculated using 2−ΔΔCt method and normalized to β-actin.
F, forward; R, reverse.
Western blot analysis
Proteins from mice lung tissues or BM-Mø were isolated using RIPA lysis buffer (Sigma). Protein concentrations were measured by BCA (Sigma). Then, 40 μg total proteins were separated by 10% SDS-PAGE gel and transferred to PVDF membranes (Sigma). After incubated with 0.5% skimmed milk power at room temperature for 1 h, target proteins in the membranes were probed with primary antibodies overnight at 4°C. After washing, the membranes were incubated with corresponding horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 1.5 h. The bands were visualized using enhanced chemiluminescence (Pierce, Rockford, IL). The levels of β-actin were used as internal reference. All antibodies (Abs) were purchased from Abcam. Primary Abs: rabbit polyclonal anti-Wnt5a Ab (ab72583; 1:2000), anti-β-actin Ab (ab8227; 1:3000); Secondary Abs: HRP-conjugated goat polyclonal anti-rabbit IgG Ab (ab6721; 1:4000).
Cell apoptosis assay
Mice BM-Mø prepared above were allowed to receive heat-inactivated H37Rv or H37Rv infection at a MOI of 6: 1 for 12 h. Then, cell apoptosis was analyzed by flow cytometry. Cells were stained by reagents in Annexin V-FITC Apoptosis Detection Kits (Sigma) as per the manufacturer's proposal. In brief, cells were collected postinfection and washed twice with PBS and then resuspended in 1× binding buffer (1 × 106 cells/mL) in a test tube. Next, 5 μL Annexin V-FITC conjugate and 10 μL propidium iodide solution were added into the tube sequentially. After being incubated in the dark for 10 min, cells were analyzed using a FACS analyzer (BD Biosciences, San Jose, CA).
Statistical analyses
All experiments were performed in triplicate in three independent assays. Data were analyzed by Student's t-test, expressed as mean ± standard deviation, and normalized to β-actin. *p < 0.05 or **p < 0.01 indicates a statistically significant difference. All the statistical analyses were performed by SPSS 19.0 software. For Western blots, data were analyzed using Image-Pro 6.0 software.
Results
Wnt5a is reduced in MTB-infected mice lung tissues or BM-Mø
As shown in Figure 1A, data from colony enumeration assays proved that H37Rv aerosol infection procedures were successful, which can satisfy the following research. Through flow cytometry, we confirmed that BM-Mø were successfully induced by M-CSF (Fig. 1D). To determine the proper dose and time of MTB infection, mice were treated with H37Rv by different CFU, and BM-Mø were infected with H37Rv by different MOI as described in methods. In vivo assay, results shown in Figure 1B indicate that Wnt5a mRNA level is significantly reduced only at 1000 CFU (*p < 0.05) posttreatment for 7 days with a slight change. Based on the above result, the rest of mice in 0 or 1000 CFU group was continuously raised to 14 or 21 days. As shown in Figure 1C, we observed that Wnt5a mRNA levels were markedly reduced with infection time in 1000 CFU group (*p < 0.05). In vitro assay, the result (Fig. 1E) suggests that Wnt5a mRNA level is significantly reduced at each MOI (*p < 0.05) postinfection for 4 h. Based on the results, BM-Mø in 0:1 or 6: 1 MOI group were continuously cultured for 8 or 12 h. Similarly, Wnt5a mRNA levels were dramatically reduced with time in 6:1 MOI group (*p < 0.05, Fig. 1F). Together, these results reveal that Wnt5a ubiquitously reduced temporality and spatiality in H37Rv-treated mice lung tissues or in H37Rv-infected mice BM-Mø. And also, 1000 CFU for 21 days in vivo and 6: 1 MOI for 12 h in vitro were chosen for following experiments.
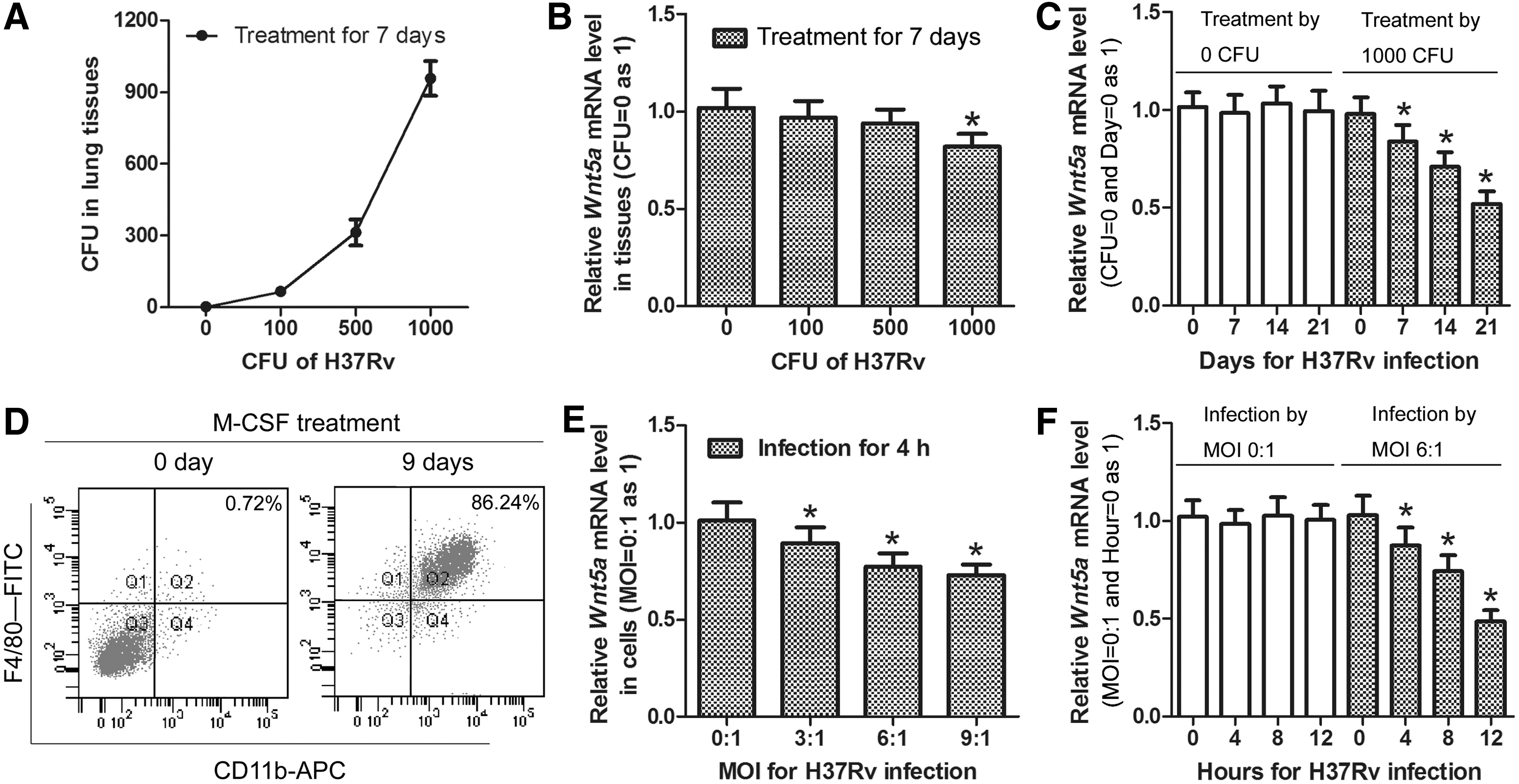
Wnt5a mRNA levels in H37Rv-treated mice lung tissues or H37Rv-infected mice BM-Mø. Mice were allowed to receive H37Rv treatment by different CFU (0, 100, 500, or 1000) for indicated days (0, 7, 14, or 21). The success of H37Rv aerosol infection procedure by different CFU for 7 days was verified by colony enumeration assays.
Wnt5a deficiency attenuates MTB-induced inflammatory cytokine secretion in mice lung tissues
To explore the biological significance of reduced Wnt5a for MTB-treated mice, expression of Wnt5a was further silenced artificially by siRNA interference in vivo, which was verified by RT-qPCR (Fig. 2A) and Western blot (Fig. 2B). Then, mRNA levels of inflammatory cytokines were detected in mice lung tissues. The results shown in Figure 2C suggest that TNF-α mRNA level was significantly higher in H37Rv-treated group than that in untreated group (**p < 0.01), which is due to multifactor and can be partly reversed in H37Rv-treated Wnt5a-silenced group (*p < 0.05). And also, similar results can be observed for IL-1β (Fig. 2D), IL-12 (Fig. 2E), and IL-6 (Fig. 2F) detection. These results indicate that Wnt5a with almost entire deficiency can partially attenuate MTB-induced inflammatory responses in mice.

Wnt5a and inflammatory cytokine levels in mice lung tissues. Mice were allowed to receive H37Rv infection or not by 1000 CFU for 21 days. Wnt5a levels were verified by RT-qPCR and Western blot. Inflammatory cytokine levels were detected by RT-qPCR.
Inhibition of Wnt5a by Sfrp1 overexpression attenuates MTB-induced inflammatory cytokine secretion in mice BM-Mø
Continuously, in vitro, we further explored the effects of up- or downregulated Wnt5a on inflammatory cytokine secretion in mice BM-Mø. First, the up- or downregulation of Wnt5a was fulfilled in Sfrp1-silenced or -overexpressed mice BM-Mø, which were verified by RT-qPCR (Fig. 3A) and Western blot (Fig. 3B). Following, mRNA levels of inflammatory cytokines were detected. The results shown in Figure 3C suggest that TNF-α mRNA level was markedly higher in H37Rv-infected cells than that in uninfected or heat-inactivated H37Rv-infected cells (**p < 0.01), which can be attenuated in H37Rv-infected Sfrp1-overexpressed cells (*p < 0.05), but enhanced in H37Rv-infected Sfrp1-silenced cells (*p < 0.05). Similar results can also be observed for IL-1β (Fig. 3D), IL-12 (Fig. 3E), and IL-6 (Fig. 3F) detection. For these inflammatory cytokines, there was no difference between uninfected and heat-inactivated H37Rv-infected cells. These results reveal that inhibition of Wnt5a by Sfrp1 can attenuate MTB-induced inflammatory cytokine secretion in mice BM-Mø, which is in accordance with the results in vivo.
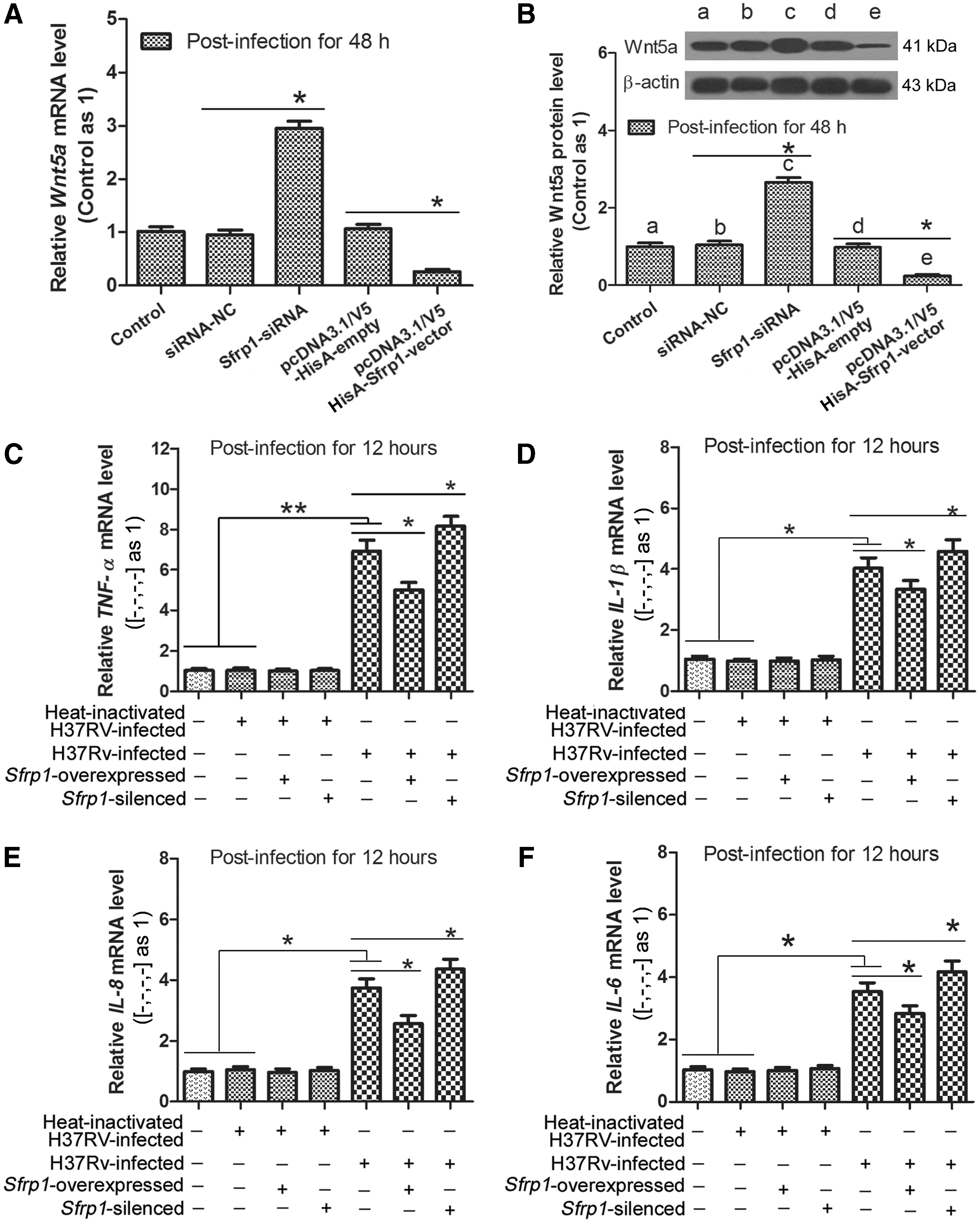
The effects of up- or downregulated Wnt5a on inflammatory cytokine secretion in mice BM-Mø. Mice BM-Mø prepared in methods were allowed to receive heat-inactivated H37Rv infection or H37Rv infection by a MOI of 6:1 for 12 h. Wnt5a levels were verified by RT-qPCR and Western blot. Inflammatory cytokine levels were detected by RT-qPCR.
Inhibition of Wnt5a by Sfrp1 induces mice BM-Mø trending to M2 phenotype in MTB-infected cells
To examine whether the reduced Wnt5a has an influence on the phenotype of mice BM-Mø, the expression of arginase-1 (Arg-1), mannose receptor (MRC-1), TGF-β, and TNF-α was measured in heat-inactivated H37Rv-infected, H37Rv-infected, or uninfected cells by RT-qPCR, and Western blot. The results shown in Figure 4A suggest that Arg-1 level is dramatically decreased in H37Rv-infected cells (*p < 0.05), which can be slightly reversed in H37Rv-infected Sfrp1-overexpressed cells (*p < 0.05). Similar results can be observed for MRC-1 (Fig. 4B) and TGF-β (Fig. 4C) detection, while opposite results for TNF-α detection (Fig. 4D). For these molecules, there was no difference between uninfected and heat-inactivated H37Rv-infected cells. The results suggest that inhibition of Wnt5a by Sfrp1 can induce mice BM-Mø trending to M2 phenotype in MTB-infected cells, which hints that the reduced Wnt5a can also have a regulation effect on BM-Mø phenotype. This finding reveals a novel way for Mø to manipulate themselves to resistant MTB-induced immune responses.

Changes of Arg-1, MRC-1, TGF-β, and TNF-α levels in mice BM-Mø. Mice BM-Mø prepared in methods were allowed to receive heat-inactivated H37Rv infection or H37Rv infection by a MOI of 6:1 for 12 h. Levels of mRNA were measured in BM-Mø by RT-qPCR for Arg-1
Inhibition of Wnt5a by Sfrp1 leads to apoptosis increase of MTB-infected mice BM-Mø
To explore whether the reduced Wnt5a influences the apoptosis of MTB-infected mice BM-Mø, cell apoptosis was detected in heat-inactivated H37Rv- infected, H37Rv-infected, or uninfected cells (Fig. 5A). As shown in Figure 5B, cell apoptosis rate was significantly increased in H37Rv-infected cells (*p < 0.05), which can be further enhanced in H37Rv-infected Sfrp1-overexpressed cells (*p < 0.05). These results indicate that inhibition of Wnt5a by Sfrp1 can lead to apoptosis increase of MTB-infected mice BM-Mø, which results in the destruction of survival environment for MTB. Hence, we conclude that the reducing of Wnt5a is a tactful strategy adopted by mice BM-Mø to manipulate theirselves to resistant MTB infection.
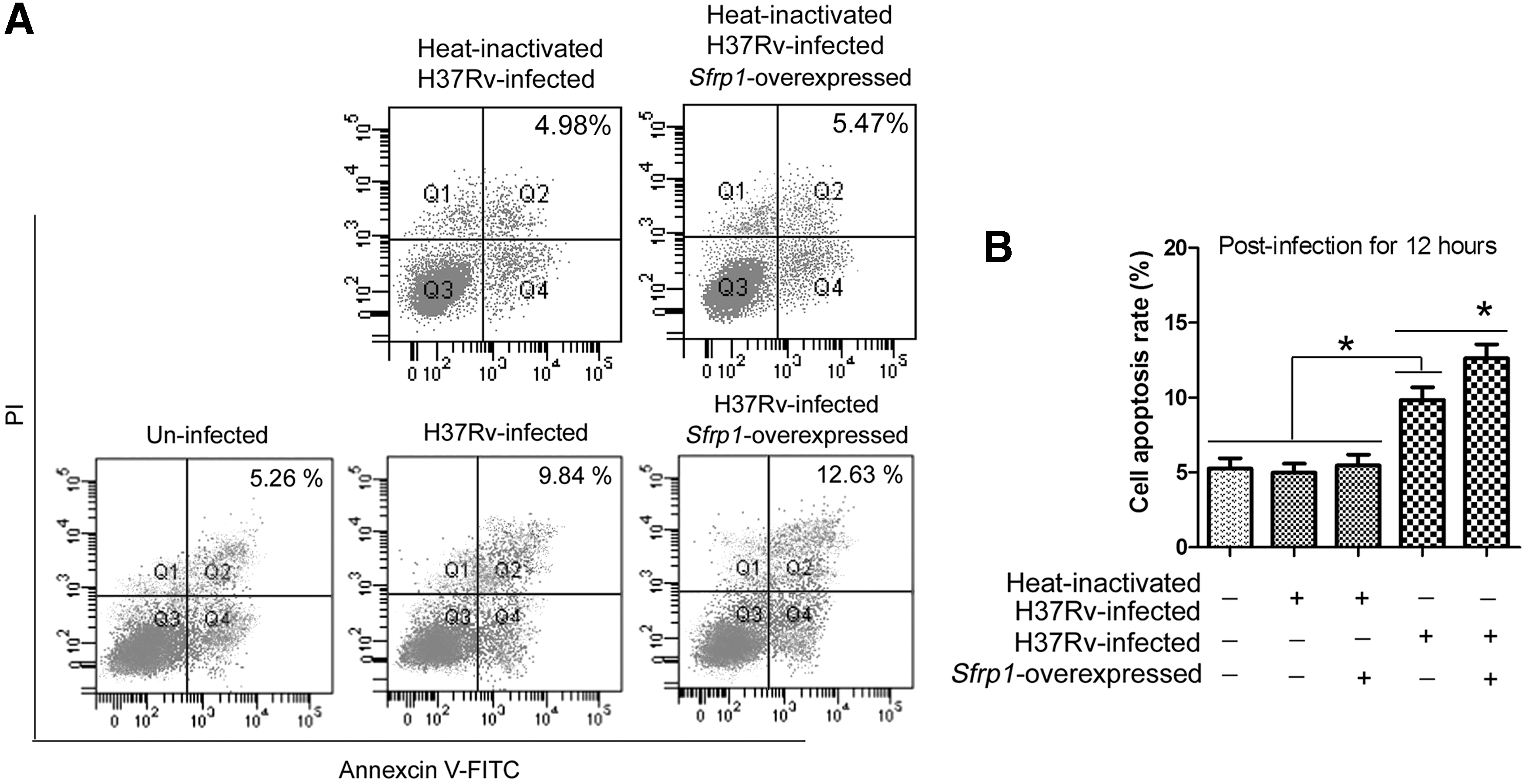
Changes of cell apoptosis in mice BM-Mø. Mice BM-Mø prepared in methods were allowed to receive heat-inactivated H37Rv infection or H37Rv infection by a MOI of 6:1 for 12 h.
Discussion
Wnt proteins are well known for their role in embryonic development and tissue homeostasis (Angers and Moon, 2009; van Amerongen and Nusse, 2009). Based on recently accumulated evidences, Wnt signaling was also shown to be involved in the regulation of inflammatory processes and infectious diseases (Schaale et al., 2011). In this study, we discovered that external disturbance triggered Wnt5a lower expression can attenuate inflammatory cytokine secretion, induce Mø to trend to M2 phenotype, and increase cell apoptosis of MTB-infected mice BM-Mø, which may lead to a prominent protective effect on mice from MTB infection. Our data revealed a novel strategy that was adopted by Mø to manipulate themselves to resistant MTB-induced immune responses and enhance MTB-induced Mø apoptosis in mice, which can increase our understanding of the interaction between MTB and Mø.
Currently, the pro-inflammatory effects of Wnt5a have been found in several inflammatory diseases, such as sepsis (Chen et al., 2015), atherosclerosis (Bhatt et al., 2012), and rheumatoid arthritis (de Sousa Rabelo et al., 2010). Wnt5a can act through Ca2+/calmodulin-dependent protein kinase (CaMKII) and that this pathway can contribute to the inflammatory response of Mø (Pereira et al., 2008), which is the first cell type encountered by invaded MTB (Ernst, 1998). A Blumenthal et al. (2006) previously found that Wnt5a was induced in MTB-infected human Mø and plays a pro-inflammatory effect. Similarly, our data also identified the pro-inflammatory roles of Wnt5a in MTB-infected mice BM-Mø. Differently, reduction of Wnt5a was ubiquitously observed in MTB-treated mice lung tissues or MTB-infected mice BM-Mø in our study, which was in accordance with the findings by Reiling and colleagues (Schaale et al., 2013). The different response of Wnt5a expression to MTB-infection in human and mice Mø may hint different interaction pattern between MTB and Mø.
In our study, Wnt5a was proved to be significantly reduced in MTB-treated mice lung tissues or MTB-infected mice BM-Mø in dose and time-dependent manner. In Wnt5a deficiency mice lung tissues or BM-Mø, MTB-induced increases of pro-inflammatory cytokines, including TNF-α, IL-1β, IL-12, and IL-6, can be markedly attenuated, which indirectly reflected the pro-inflammatory effect of Wnt5a during MTB infection. Among these cytokines, TNF-α is essential for lung granuloma formation of tuberculosis patients (Chensue et al., 1994).
For the past few years, the concept of Mø polarization has been widely developed and further sophisticated (Schaale et al., 2013). At present, Mø are deemed as a spectrum of phenotypic subtypes ranging from classically activated Mø (M1) to the alternatively activated Mø (M2) based on gene expression induced in response to pathogen- or cytokine-derived stimulation (Benoit et al., 2008; Mosser and Edwards, 2008). In this study, meanwhile, we discovered that Wnt5a deficiency can contribute to the tendency of mice Mø to be M2 phenotype, which can be reflected by the slightly reversed expressions of Arg-1, MRC-1, TGF-β, and TNF-α in MTB-infected Wnt5a deficiency mice BM-Mø. Among the above molecules, Arg-1 and MRC-1 are the well-known markers of M2 Mø (Yang et al., 2012).
As always, the interaction between MTB and Mø is complex and difficult to illustrate. When MTB enters into the distal airways, patrolling Mø can provide a critical intracellular niche that is required for MTB to infect alveolar Mø and establish infection in the human host (Lee et al., 2009). However, induction of Mø apoptosis is associated with the killing of intracellular MTB (Molloy et al., 1994; Oddo et al., 1998). In our study, Mø apoptosis was significantly increased in MTB-infected mice BM-Mø and further markedly enhanced in MTB-infected Wnt5a deficiency mice BM-Mø, which are unfavorable for MTB survival. Hence, the reduction of Wnt5a in MTB-infected mice BM-Mø is a tactful strategy adopted by Mø against MTB infection. Besides, we guess that Wnt5a deficiency induced BM-Mø apoptosis increase may entirely or partially contribute to the reduction of inflammatory cytokine secretion mentioned above, which will be illuminated in our further study. In summary, our study revealed that Wnt5a deficiency can lead to a resistance of Mø to MTB-induced immune responses and reinforce MTB-induced Mø apoptosis in mice.
Footnotes
Disclosure Statement
No competing financial interests exist.