
Abstract
Atherosclerosis is a common pathological basis of cardiovascular disease, which remains the leading cause of mortality. Long noncoding RNAs (lncRNAs) are newly studied non-protein-coding RNAs involved in gene regulation, but how lncRNAs exert regulatory effect on atherosclerosis remains unclear. In this study, we found that lncRNA HOXC cluster antisense RNA 1 (HOXC-AS1) and homeobox C6 (HOXC6) were downregulated in carotid atherosclerosis by performing microarray analysis. The results were verified in atherosclerotic plaques and normal arterial intima tissues by quantitative reverse transcription PCR and western blot analysis. Lentivirus-mediated overexpression of HOXC-AS1 induced HOXC6 expression at mRNA and protein levels in THP-1 macrophages. Besides, oxidized low-density lipoprotein (Ox-LDL) decreased expression of HOXC-AS1 and HOXC6 in a time-dependent manner. Induction of cholesterol accumulation by Ox-LDL could be partly suppressed by overexpression of HOXC-AS1.
Introduction
A
In recent years, a great deal of interest has focused on understanding biologic function of long non-protein-coding RNAs (long noncoding RNAs [lncRNAs]). lncRNAs usually contains more than 200 nucleotides and consist of a variety of RNAs including long intergenic ncRNAs (lincRNAs) and so on (Kung et al., 2013). lncRNAs were originally regarded as meaningless noise during genome transcription. Recently, massive researches provide us new insight into the fact that lncRNAs can regulate gene expression by executing as signals, decoys, guides, and scaffolds and acting as inhibitors or activators to modulate the transcription and translation of genes (Wang and Chang, 2011). A tremendous amount of studies have already shown the involvement of lncRNAs in cancer development (Fang and Fullwood, 2016), however, there have been limited publications reporting direct evidence that lncRNAs are closely associated with atherosclerosis. A large prospective study by Wu et al. (2014) found that the expression of lincRNA-p21 was dramatically downregulated in atherosclerotic plaques of ApoE−/− mice, and lincRNA-p21 promoted apoptosis and repressed cell proliferation in vascular smooth muscle cells (VSMCs) by feeding back to increase p53 transcriptional activity, while the proliferation of VSMCs was crucial during atherosclerotic lesion development. Our group has found that lincRNA -DYNLRB2-2 increased the expression of G protein-coupled receptor 119 (GPR119), which markedly promoted ATP-binding cassette transporter A1 (ABCA1)-induced cholesterol efflux and reduced lesion area in ApoE−/− mice (Hu et al., 2014). In addition, we also found that lncRNA RP5-833A20.1 was involved in regulating cholesterol homeostasis by inducing hsa-miR-382-5p expression and decreasing expression of nuclear factor IA (NFIA), which increased reverse cholesterol transport and suppressed formation of atherosclerotic plaques in ApoE−/− mice (Hu et al., 2015). Therefore, to gain novel insights into the mechanism by which lncRNAs exert regulatory effect on CVD, especially atherosclerosis, has been a significant area of research.
Homeobox genes are a family comprising at least 200 transcription factors that shared a common highly conserved 61 amino-acid homeodomain that can bind to DNA (Stein et al., 1996). Human genome possesses 39 HOX genes, which are organized into four different clusters, HOXA, HOXB, HOXC, and HOXD (Visconti and Awgulewitsch, 2015). HOX genes regulated the identity of different regions along the mammalian body axis, and the rules mainly complied with temporal and spatial colinearity (Dekker et al., 1992). Accumulating evidence indicated that some members of HOX genes exert significant regulatory effect on cardiovascular system, for instance, HOXA5, HOXA11, HOXB1, HOXB7, and HOXC9 participated in modulating vasculature functions (Miano et al., 1996). HOXD3 was reported to regulate the conversion from resting endothelial cells (ECs) state to invasive state, and HOXD3 was highly expressed in active proliferating EC forming tubes (Boudreau et al., 1997). Substantial studies paid close attention to the association between homeobox C6 (HOXC6) and mammary gland development and found that its expression is upregulated in a variety of cancers including breast and prostate cancers (Hamid et al., 2015; Hussain et al., 2015). Moreover, a study demonstrated that the expression of HOXC6 was quite different in human vascular wall-resident CD44 (+) multipotent stem cells (VW-MPSCs), terminal differentiated human aortic smooth muscle cell (SMC), and undifferentiated pluripotent embryonic stem cells, and thus, HOXC6 may regulate the differentiation of CD44 (+) VW-MPSCs to SMCs (Klein et al., 2013). Nevertheless, no publication has revealed association between HOXC6 and atherosclerosis.
In our present study, microarray analysis revealed that both lncRNA HOXC cluster antisense RNA 1 (HOXC-AS1) and HOXC6 were downregulated in carotid atherosclerosis when compared to normal renal arterial intima. Next, Bioinformatics analysis revealed that lncRNA HOXC-AS1 and HOXC6 were both located on chromosome 12 and their transcription positions were nearby. In addition, overexpression of lncRNA HOXC-AS1 promoted expression of HOXC6 at mRNA and protein levels. We also found that overexpression of lncRNA HOXC-AS1 could partly inhibit Ox-LDL-induced accumulation of cholesterol in human acute monocytic leukemia (THP-1) macrophages.
Materials and Methods
Ethics and consent
Informed consent was obtained from all patients and ethical approval was obtained from the Institutional Review Board of Nanfang Hospital, Southern Medical University (Guangzhou, China). All of the patients had not received medication before surgery.
Sources of tissue samples
All of the specimens in this research were obtained between April 2013 and April 2014 in the Department of Vascular Surgery, Nanfang Hospital, Southern Medical University. Eight renal arteries without atherosclerotic plaques were obtained from patients undergoing renal transplantation surgery as control. Eight human atherosclerotic plaques were collected from patients undergoing carotid endarterectomy. Fresh, unfixed material was used for extracting protein and RNA.
Materials
Ox-LDL was obtained from Biomedical Technologies, Inc. (Stoughton, MA). The PrimeScript RT Reagent Kit (Perfect Real Time) (DRR037A) and SYBR® Premix Ex TaqTM II (Tli RNaseH Plus) (DRR820A) were purchased from TaKaRa Bio, Inc. (Shiga, Japan). All other chemicals of pharmaceutical grade were purchased from commercial suppliers.
Cell culture
Human monocytic THP-1 cells were obtained from American Type Culture Collection (ATCC, Manassas, VA). THP-1 cells were cultured with Roswell Park Memorial Institute (RPMI) 1640 medium containing 10% fetal calf serum and incubated at 37° in an atmosphere of 5% CO2. THP-1 cells differentiated into macrophages after being incubated with phorbol 12-myristate 13-acetate (PMA, 160 nM) for 24 h.
RNA isolation and real-time quantitative PCR analysis
Total RNA from human tissues and cultured cells was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA) in accordance with the manufacturer's instructions. The mRNA levels were evaluated by quantitative reverse transcription PCR (qRT-PCR) with SYBR-Green Detection chemistry (TaKaRa Bio, Inc.) by ABI 7500 Fast Real-Time PCR system. The expression of GAPDH was used as the internal control for analysis of mRNA level of HOXC6. The expression of U6 RNA was used as the endogenous control for analysis of lncRNA HOXC-AS1 from tissues and cells. ΔΔCt method was used for determining quantitative measurements. All samples were measured in triplicate and the mean value was considered for comparative analysis. Human HOXC-AS1 primers: forward 5′-CTGCGACACTTCCCCACC-3′ and reverse 5′-CTACTT GCCCACGACCGA-3′. Human U6 primers: forward 5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAA TTTGCGT-3′. Human HOXC6 primers: forward 5′-ACAGACCTCAATCGCTCAGGA-3′ and reverse 5′-AGGGGTAAATCTGGATACTGGC-3′. Human GAPDH primers: forward 5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse 5′-TGGTGAAGACGCCAGTGGA-3′.
Western blot analysis
Cells and human tissues were harvested and protein extracts were prepared according to established methods. Proteins were extracted from cultured cells and tissues using radio-immunoprecipitation assay buffer (Biocolor Ltd., Belfast, Northern Ireland, United Kingdom), quantified using the BCA protein assay kit (KeyGen Biotechnologies, Nanjing, China), and then separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and subjected to western blot analysis using rabbit polyclonal anti-HOXC6 antibodies and rabbit polyclonal anti-β-actin-specific antibodies (Abcam, Inc., Cambridge, MA). The proteins were visualized using a chemiluminescence method (ECL Plus Western Blot Detection System; Amersham Biosciences, Foster City, CA).
Lentivirus production and cells infection
Packed empty lentivirus vectors (LVs) with green fluorescent protein (GFP; LV-Mock), LV-mediated lncRNA HOXC-AS1 overexpression vector (LV-HOXC-AS1) with GFP were prepared. The cells were infected with the LV stock at a multiplicity of infection of 100 (THP-1 cells) transducing units per cell in the presence of 8 μg/mL of polybrene. The cells were washed with fresh complete media after 24 h of incubation. The GFP-positive cells were counted 96 h post-transduction.
HPLC analysis of cellular cholesterol levels
HPLC analysis was conducted as described previously (Hu et al., 2009). The sterol analysis used a HPLC system (model 2790, controlled with Empower Pro software; Waters Corp., Milford, MA). Absorbance at 216 nm was monitored. Data were analyzed with TotalChrom software from PerkinElmer (Waltham, MA).
Statistical analysis
Results were expressed as the mean ± standard deviation. The results were analyzed by one-way analysis of variance and Student's t-test, followed by the Student-Newman-Keuls (SNK) test using SPSS v13.0 statistical software (SPSS, Inc., Chicago, IL). A two-tailed probability (p)-value <0.01 was considered statistically significant.
Results
The expression levels of lncRNA HOXC-AS1 and HOXC6 were downregulated in atherosclerotic tissues
Microarray analysis of carotid atherosclerosis and normal renal arterial intima tissues was carried out to explore possible changes in RNA levels during atherosclerosis development with the Arraystar probe dataset. Information of patients was as described previously (Lu et al., 2016). Among the differentially expressed RNAs, expression of HOXC-AS1 was decreased 31.77 times (n = 3, p = 1.12E-07) while the expression of HOXC6 was decreased 4.64 times (n = 3, p = 0.00013) in carotid atherosclerosis. To further verify these findings in microarray we performed qRT-PCR to determine the expression of HOXC-AS1 and HOXC6 in five advanced atherosclerotic plaques (Grade V or Grade VI) and five normal renal arterial intima tissues. The mRNA level of HOXC-AS1 was decreased to 7.82% while the mRNA level of HOXC6 was reduced to 25.76% in atherosclerotic plaques (Fig. 1A, B). Subsequently, we performed western blot analysis to determine the protein level of HOXC6, and we found the protein level of HOXC6 was reduced to 23.12% in atherosclerotic plaques (Fig. 1C). These results indicated that the expression levels of HOXC-AS1 and HOXC6 were both decreased in human atherosclerotic plaques when compared to normal intima tissues.
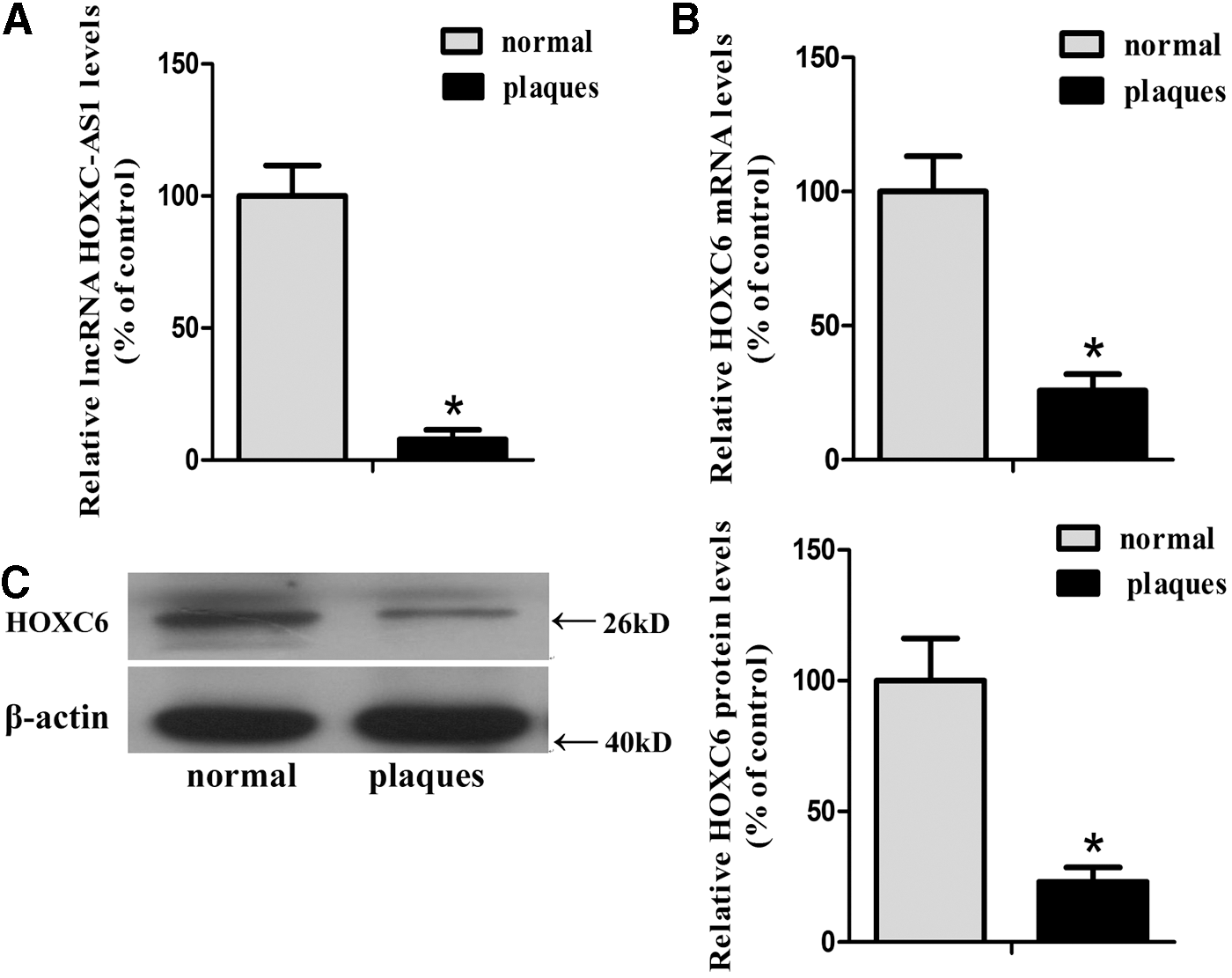
Expression of lncRNA HOXC-AS1 and HOXC6 in normal tissues and atherosclerotic plaques.
The expression of HOXC6 was promoted by lncRNA HOXC-AS1 in THP-1 macrophages
As indicated above, microarray analysis, qRT-PCR, and western blot analysis revealed that the expression levels of HOXC-AS1 and HOXC6 were both downregulated in the atherosclerotic plaques. Next, Bioinformatic analysis revealed that HOXC-AS1 (Chr 12:53,999,022-53,999,620) and HOXC6 (Chr 12:53,994,944-54,029,689) were both located on chromosome 12 and they had opposite transcription directions (Fig. 2A). Although HOXC9, HOXC8, and HOXC5 are also located near HOXC-AS1 on Chr12, the microarray analysis results showed that the p value of HOXC9, HOXC8, and HOXC5 is >0.05. Several studies indicate that lncRNAs may control expression of protein-coding genes located nearby (Lee, 2009; Mercer et al., 2009; Moran et al., 2012). Therefore, we raised the hypothesis that HOXC-AS1 promoted HOXC6 expression and then investigated the effect of HOXC-AS1 on HOXC6 in THP-1 macrophages. As shown in Figure 2B and C, after using LVs to deliver the gene of HOXC-AS1 into THP-1 macrophages, the RNA level of HOXC-AS1 was enhanced to 4275.21% while the mRNA level of HOXC6 was increased to 901.26% than that of THP-1 macrophages transfected with empty lentiviral vectors (LV-Mock). Subsequently, we evaluated the protein level of HOXC6 by western blot analysis and found that HOXC6 was increased to 428.26% by lentivirus-mediated overexpression of HOXC-AS1 (Fig. 2D). Data presented here demonstrated that HOXC-AS1 promoted HOXC6 expression in THP-1 macrophages at both mRNA and protein levels.
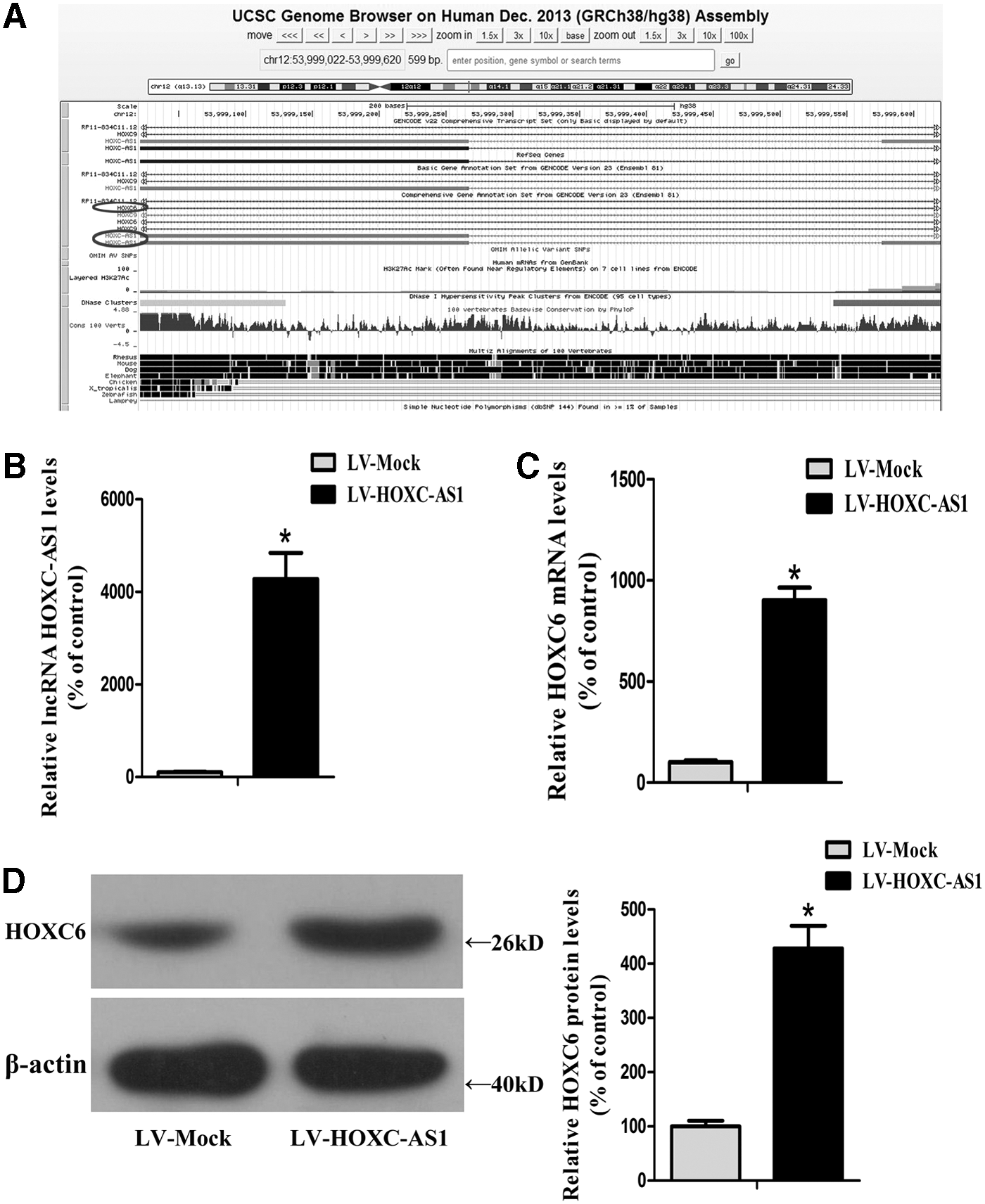
Effect of lncRNA HOXC-AS1 on HOXC6 in THP-1 macrophages.
The expression levels of lncRNA HOXC-AS1 and HOXC6 could be suppressed by Ox-LDL in THP-1 macrophages
It is long known that circulating monocytes adhere to the intima and differentiate to macrophages, which transform into foam cells after taking in Ox-LDL (Tabas, 2010). Atherosclerotic plaques gradually form along with the stock of foam cells, which become the major part of necrotic core of plaques. In addition, data presented here indicated that expression levels of HOXC-AS1 and HOXC6 were both reduced in atherosclerotic tissues. Thus, we legitimately expected to propose the hypothesis that the downregulation of HOXC-AS1 and HOXC6 were due to Ox-LDL. To test the hypothesis, we treated THP-1 macrophages with 50 μg/mL of Ox-LDL and found that Ox-LDL obviously decreased expression RNA levels of HOXC-AS1 and HOXC6 and protein level of HOXC6 in a time-dependent manner in THP-1 macrophages (Fig. 3A–C). Treatment with Ox-LDL for 48 h was the most effective at decreasing expression of HOXC-AS1 and HOXC6. In addition, Ox-LDL suppressed expression of HOXC6 in absence of HOXC-AS1. After transfecting THP-1 cells with lentivirus to overexpress HOXC-AS1, the inhibition of Ox-LDL on HOXC6 was totally compensated (Fig. 3D). Therefore, our data presented here demonstrated a causative role for the inhibition of Ox-LDL on HOXC-AS1 and HOXC6. Besides, HOXC-AS1 might mediate the regulation of Ox-LDL on HOXC6 in THP-1 cells.
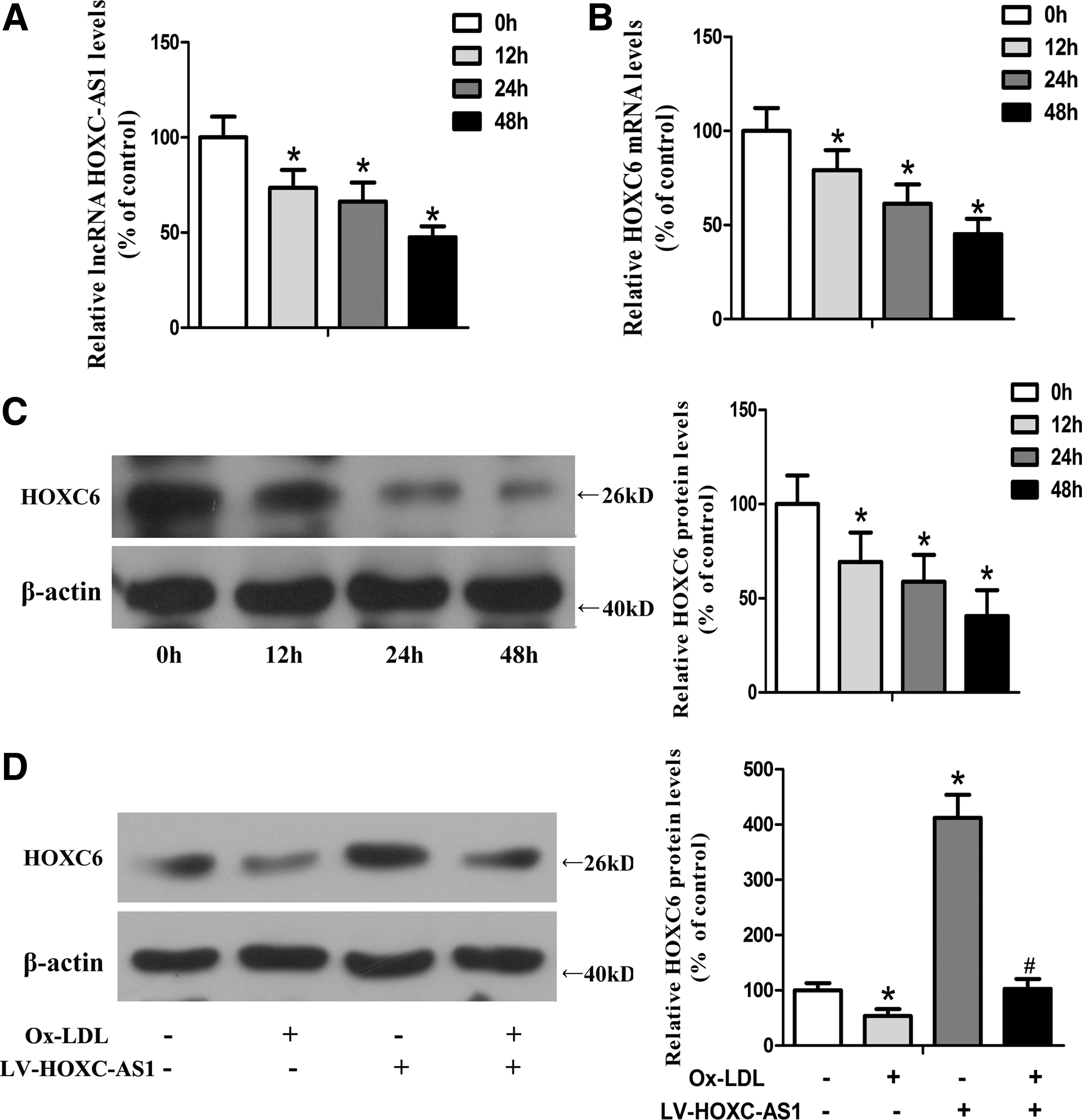
Ox-LDL suppressed the expression of lncRNA HOXC-AS1 and HOXC6 in THP-1 macrophages.
Cholesterol accumulation induced by Ox-LDL could be inhibited by lncRNA HOXC-AS1 in THP-1 macrophages
It is well accepted that Ox-LDL promotes formation of foam cells and disorder of cholesterol homeostasis in macrophages. And we have revealed the regulatory effect of Ox-LDL on HOXC-AS1. We then performed HPLC to investigate the effect of Ox-LDL and HOXC-AS1 on cholesterol content in THP-1 macrophages. As shown in Table 1, the levels of total cholesterol (TC), free cholesterol (FC), and cholesterol ester (CE) were markedly increased after treating THP-1 macrophages with 50 μg/mL of Ox-LDL. Besides, the induction of cholesterol content by Ox-LDL could be partly suppressed by lentivirus-mediated overexpression of HOXC-AS1.
N = 5, p < 0.01 versus LV-Mock group.
N = 5, p < 0.01 versus LV-Mock+Ox-LDL group.
CE, cholesterol ester; FC, free cholesterol; HOXC-AS1, HOXC cluster antisense RNA 1; LV, lentivirus vectors; Ox-LDL, oxidized low-density lipoprotein; TC, total cholesterol.
Discussion
Atherosclerosis is a common disease in clinical practice, and pathological basis of ischemic cardiovascular and cerebrovascular diseases (Hansson, 2005). Dyslipidemia is the foremost reason that leads to atherosclerosis and reduction of cholesterol can decrease the risk of death from CVD (Panel, 2006). Analysis of lncRNAs in balance of cholesterol metabolism is still in its infancy and their role in atherosclerosis is yet to be realized. In this study, we showed for the first time that Ox-LDL could downregulate HOXC6 by reducing HOXC-AS1, which partly suppressed Ox-LDL-induced cholesterol accumulation.
lncRNAs are a recently studied type of non-protein-coding RNAs and gradually proved to be participating in regulating a wide variety of biological processes. lncRNAs can be classified into several subgroups according to genomic location, and the groups include lincRNAs, natural antisense transcripts (NATs), transcribed ultra-conserved regions (TUCRs), and enhancer-like ncRNAs (eRNAs) (Kung et al., 2013). lncRNAs regulate gene expression by reacting with stimulus or transducing signals in cells, controlling relative proteins and microRNAs, and guiding target molecules to specific nearby or distant regions to control gene expression (Zhou et al., 2016). Evidence from several recent studies indicates that lncRNAs may regulate expression of protein-coding genes located nearby (Lee, 2009; Mercer et al., 2009; Moran et al., 2012). In our present study, microarray analysis showed that expression levels of HOXC-AS1 and HOXC6 were both downregulated in atherosclerotic plaques. We verified the results in advanced atherosclerotic plaques and normal intima tissues, and the results were in accordance with those from microarray analysis. Bioinformatics analysis revealed that HOXC-AS1 and HOXC6 were both located on chromosome 12 and their transcription positions were nearby. Furthermore, our results showed that overexpression of HOXC-AS1 markedly increased expression of HOXC6 at both mRNA and protein levels. Thereby, these findings indicated that HOXC-AS1 may promote HOXC6 expression. The detailed mechanism of induction of HOXC6 by HOXC-AS1 needs further clarification.
The internalization of Ox-LDL by macrophages results in formation of lipid peroxides, accumulation of CE, and final formation of foam cells, which constitute the necrotic core of atherosclerotic plaques (Ross, 1999). Meanwhile, the uptake of Ox-LDL by monocyte-derived macrophages via scavenger receptors lacks negative feedback regulation and leads to uncontrolled accumulation of Ox-LDL in macrophages (Siegel-Axel et al., 2008; Badimon et al., 2011). Besides, Ox-LDL can promote endothelial dysfunction and lead to inhibition of adhesion molecules, then results in recruitment of more and more monocytes into the subintima space (Ketelhuth and Hansson, 2011). Therefore, Ox-LDL plays an important role in the formation of foam cells. Besides, our results found the expression levels of HOXC-AS1 and HOXC6 were downregulated in atherosclerotic plaques and our group previously revealed that Ox-LDL could regulate the expression of lncRNA (Hu et al., 2014, 2015). Thus, we explored the possibility that the downregulation of HOXC-AS1 and HOXC6 expression was induced by Ox-LDL. Our study represents the first endeavor for screening the downregulation of HOXC-AS1 and HOXC6 by Ox-LDL in THP-1 macrophages in a time-dependent manner. The reduction of HOXC6 by Ox-LDL could be compensated by overexpression of HOXC-AS1. These results provided strong evidence supporting the notion that HOXC-AS1 mediated the downregulation of HOXC6 by Ox-LDL. The fact that induction of cholesterol accumulation by Ox-LDL cannot be completely suppressed by HOXC-AS1 suggests that other mediators may be involved in the process. Besides, whether Ox-LDL regulated HOXC-AS1 directly or indirectly by other mechanism needs to be further explored.
The human HOX genes are a family that contains a common conserved homeodomain and plays a vital role in development and differentiation (Ramachandran et al., 2005). Several members of HOX genes are reported to be involved in regulation of vascular system. HOXB7 and HOXC9 were reported to play a role in modulating vasculature functions by participating in specifying the phonotype of fetal SMC (Miano et al., 1996). In addition, inhibition of HOXA9 could suppress formation of tube and EC migration, which suggested that HOXA9 was involved in regulating angiogenesis (Bruhl et al., 2004). HOXC6 is one of 39 human HOXC genes involved in mammary gland development and is found to be differentially expressed in a variety of cancers (Hamid et al., 2015; Hussain et al., 2015). However, there was no publication ever reporting the direct association between HOXC6 and atherosclerosis. A study has revealed that expression of HOXC6 is upregulated after fat loss due to bariatric surgery (Dankel et al., 2010). In this study, we showed that Ox-LDL promoted cholesterol accumulation via increasing the levels of TC, FC, and CE. HOXC-AS1 partly inhibited the induction of Ox-LDL on cholesterol accumulation. Thus, it is reasonable to speculate that HOXC-AS1 mediated regulation of Ox-LDL on cholesterol accumulation. Besides, Ox-LDL suppressed expression of HOXC-AS1 and HOXC6 in THP-1 cells. HOXC-AS1 promoted expression of HOXC6 in THP-1 cells. Therefore, these results indicated that HOXC6 may be a mediator involved in the regulation of Ox-LDL and HOXC-AS1 on cholesterol content and HOXC6 may exert a positive effect on lipid metabolism. The role of HOXC6 in regulation of Ox-LDL and HOXC-AS1 on cholesterol accumulation can be verified after decreasing expression of HOXC6 by using siRNA and increasing expression of HOXC6 by using virus in further study. Besides, we would investigate whether HOXC6 influences atherosclerosis formation as other members of HOX genes mentioned above via controlling the phonotype of fetal SMC, formation of tube, or EC migration. Moreover, as a transcription factor, HOXC6 may regulate transcription of genes involved in lipid metabolism via action of homeodomain or DNA binding domain with promoter or other cofactors (Michaut et al., 2011). In addition to being a transcription factor, homeodomain is revealed to be a protein–protein interaction domain and it is of great interest to explore whether HOXC6 protein interacts with proteins involved in processes regulating cholesterol metabolism (Plaza et al., 2001).
Conclusion
Our study elucidated for the first time that lncRNA HOXC-AS1 promoted expression of HOXC6 at both mRNA and protein levels. Ox-LDL decreased expression of HOXC-AS1 and HOXC6 in a time-dependent manner. Accumulation of cholesterol induced by Ox-LDL could be partly suppressed by overexpression of HOXC-AS1. Our findings provide valuable insight into the specific role of HOXC-AS1 in cholesterol homeostasis and we hope that HOXC-AS1 and HOXC6 can be promising therapeutic targets in preventing atherosclerosis.
Footnotes
Acknowledgments
This study was supported by the National Natural Sciences Foundation of China (Grant Nos. 81472009 and 81500387), Natural Science Fund of Guangdong (2014A030313287, 2014A030310135, and 2015A030313245), Medical Scientific Research Foundation of Guangdong Province (B2014245), and the President Foundation of Nanfang Hospital, Southern Medical University (2012B002, 2013B004, and 2014Z003).
Disclosure Statement
No competing financial interests exist.