
Abstract
The obligate intracellular bacterium Mycobacterium leprae is the causative agent of leprosy and primarily infects macrophages, leading to irreversible nerve damage and deformities. So far, the underlying reasons allowing M. leprae to persist and propagate in macrophages, despite the presence of cellular immunity, are still a mystery. Here, we investigated the role of autophagy, a cellular process that degrades cytosolic materials and intracellular pathogens, in M. leprae infection. We found that live M. leprae infection of macrophages resulted in significantly elevated autophagy level. However, macrophages with high autophagy levels preferentially expressed lower levels of proinflammatory cytokines, including interleukin (IL)-1β, IL-6, IL-12, and tumor necrosis factor-α, and preferentially primed anti-inflammatory T cells responses, characterized by high IL-10 and low interferon-γ, granzyme B, and perforin responses. These anti-inflammatory T cells could suppress further induction of autophagy, leading to improved survival of intracellular M. leprae in infected macrophages. Therefore, these data demonstrated that although autophagy had a role in eliminating intracellular pathogens, the induction of autophagy resulted in anti-inflammatory immune responses, which suppressed autophagy in a negative feedback loop and allowed the persistence of M. leprae.
Introduction
T
Patients with TT leprosy presented strong T helper type 1 (Th1) responses, with strong interleukin 2 (IL-2) and interferon-gamma (IFN-γ) secretion, while patients with LL leprosy presented strong Th2 and Treg responses, with IL-4, IL-5, and IL-10 dominating the cytokine secretion, and with enriched presence of Foxp3+ T cells at the lesion site (Dockrell et al., 1996; Joosten et al., 2007; Modlin, 2010; Palermo et al., 2012; Bobosha et al., 2014). Studies have found that macrophages infected with M. leprae preferentially differentiated into M2-type (low IL-10 high IL-12) macrophages, and primed regulatory T cell responses instead of Th1 or cytotoxic T cell responses (Sasiain et al., 1992; Montoya et al., 2009; Mège et al., 2011; Palermo et al., 2012; Yang et al., 2016). In addition, macrophages in TT and LL leprosy patients presented significant phenotypical and functional differences. In TT patients, macrophages under the influence of tumor necrosis factor (TNF)-α formed granulomas with CD4+ and CD8+ T cells, while in LL patients, poor granuloma formation was observed (Walker and Lockwood, 2006). Also, the anti-inflammatory M2 macrophages in LL patients presented significantly higher levels of costimulatory molecules CD68 and CD163, arginase 1, and Th2/Treg cytokines IL-10, IL-13, TGF-β, and FGF-b, than those in TT patients (de Sousa et al., 2016). The specific mechanisms underlying these differences are still unclear.
Autophagy refers to the enclosure and delivery of aged or damaged cytosolic constituents to the lysosomes for subsequent degradation or recycling (Mizushima, 2007). During periods of starvation, autophagy is induced to metabolize cytosolic macromolecules to prolong cell survival (Kuma et al., 2004). Numerous recent studies have established that autophagy actively participated in antitumor and antiviral immunity, by engulfing and eliminating intracellular pathogens, delivering pathogen-associated molecular patterns to Toll-like receptors in the endosomes, and breaking down pathogenic proteins to peptides for antigen presentation (Cooney et al., 2010; Choi and Ryter, 2011; Tang et al., 2012; Yuk et al., 2012; Deretic et al., 2015). Autophagy also likely plays multiple roles in the development and polarization of T cell responses. Autophagy counteracted the FAS/FASL-signaling and prolonged the survival of activated T cells (He and He, 2013; Gump et al., 2014). The autophagy protein Beclin-1 was found to facilitate the suppression of effector T cells by Treg cells (Verghese et al., 2014). Attenuation of autophagy in antigen-presenting macrophages and dendritic cells resulted in elevates Th17 responses (Castillo et al., 2012; Wildenberg et al., 2012). The role autophagy plays in cellular response against leprosy is not clear.
Multiple lines of evidence suggest that autophagy plays an immunological role in M. leprae infection. First, M. leprae is an obligate intracellular bacteria and within the control of the autophagosomes. Second, we previously discovered that polymorphisms in the IRGM gene, which encodes an essential protein in the assembly and regulation of the core autophagy machinery (Chauhan et al., 2015), is associated with susceptibility to leprosy (Yang et al., 2014a). Third, M. leprae infection resulted in altered expression patterns in IRGM, which occurred concurrently with shifts in macrophage function (Yang et al., 2014b, 2016). In addition, autophagy is involved in the innate and adaptive immunity against M. tuberculosis infection, another intracellular pathogen related to M. leprae (Deretic et al., 2009; Castillo et al., 2012). Therefore, we examined the potential involvement of autophagy in modulating immune responses in M. leprae infection.
Materials and Methods
Isolation and culturing of human primary cells
All procedures in this study were approved by the Ethics Board of Shanghai Skin Disease Hospital. All participants provided written informed consent. Leukapheresis samples were obtained from six healthy male individuals. The peripheral blood mononuclear cells (PBMC) were harvested by centrifuging on top of a layer of Ficoll-Paque PLUS (GE Healthcare Life Sciences) at 400 g for 30 min at 22°C. Monocytes were isolated from PBMCs using Human Monocyte Isolation Kit II (Miltenyi), and plated on a six-well plate (Corning) at 2 × 105 cells/mL. These pure monocytes were cultured in RPMI 1640 supplemented with 10% autologous serum (harvested from the blood of each individual), L-glutamine, and antibiotics for 6 days at 37°C in a humidified CO2 incubator to mature into macrophages. Medium was replaced every 2 days. Naive T cells were isolated from PBMCs using Human Naive Pan T Cell Isolation Kit (Miltenyi).
M. leprae growth, viability determination, and treatment
Live Thai53-strain M. leprae was isolated from the footpad of athymic BALB/c-nu/nu mice (McDermott-Lancaster et al., 1987; Levy and Ji, 2006). Viability and bacterial number was determined using the Live/Dead BacLight bacterial Viability and Counting Kit (Thermo Fisher Scientific) in flow cytometry, following manufacturer's instructions. Killed M. leprae was prepared by heating the bacteria in 80°C for 30 min. For the infection/stimulation of macrophages, macrophages were treated with live or killed M. leprae at 20:1 MOI-equivalent for 6 h at 33°C. Excess bacteria were removed by washing the plate twice. The macrophages were incubated for an additional 18 h at 37°C, before further assays. For the stimulation of T cells, both live and killed M. leprae were added to naive T cells at an MOI of 20:1 each.
Autophagy induction and detection
In tissue culture, cells were placed in RPMI 1640 supplemented with 10% FBS, L-glutamine, antibiotics, and HEPES (Full medium). To induce autophagy, the medium was replaced by Eagle's balanced salt solution (EBSS, Starve medium) for 4 h. Alternatively, 25 μg/mL rapamycin in Full medium was used. Staining of autophagosomes in live cells was conducted using the Cyto-ID Autophagy Detection Kit (Enzo Life Sciences) (Chan et al., 2012). Briefly, the Cyto-ID Green dye was prepared by mixing 8 μL of the cationic amphiphilic tracer (CAT) dye from the kit with 4 mL of included assay buffer. The macrophages were first incubated in phosphate-buffered saline (PBS) with 10 mM EDTA until the cells were rounded, and then lifted from the plate by pipetting. The macrophages were then washed in Full medium and mixed with the Cyto-ID Green dye for 30 min at 37°C, followed by washing and flow cytometry in FACSCalibur (BD) in the FL1 channel (530 nm) with a blue laser (488 nm).
Intracellular M. leprae survival assay
The initial M. leprae viability was determined before adding to macrophages. M. leprae was also incubated in cell-free Full medium alongside the experiments with macrophages. The macrophages were placed in lysis buffer (10% glycerol, 1% Triton X-100, and 1% Tween 20 in PBS) for 5 min and centrifuged for 1 min at 300 g to pellet cell debris. The supernatant containing internal M. leprae was centrifuged at 13,500 g in PBS for three times to pellet the bacteria and remove lysis buffer. Viability was determined using the Live/Dead BacLight bacterial Viability and Counting Kit (Thermo Fisher Scientific), and expressed as the percentage of the initial viability before incubation.
Luminex assay
For luminex assay, the cells were placed at the bottom of a 3.0-μm Transwell-24 plate (Corning) at 2 × 105 cells/mL, while the capture beads were placed at the top. At the end of indubation, the capture beads were harvested by lifting the top layer and gently aspirating the supernatant. The Milliplex Human Cytokine and CD8+ T Cell Magnetic Bead Panels (EMD Millipore) were used following manufacturer's instructions.
Statistics
One-way or two-way analysis of variance (ANOVA) with Sidak's or Dunnett's post-tests were used to compare multiple groups. Pearson's correlation test was used to examine correlation. Data analysis was conducted in Prism 6 software (GraphPad). Analysis of flow cytometry data was performed in FlowJo 10 software (Tree Star).
Results
M. leprae infection and starvation-induced autophagy in macrophages
Infection with Shigella and Salmonella was shown to result in amino acid (AA) depletion, which triggers autophagy to restore the normal AA level (Tattoli et al., 2012). Starvation-induced autophagy has a role in eliminating M. tuberculosis infection in macrophages (Gutierrez et al., 2004). We first investigated the induction of autophagy in M. leprae infection. Macrophages derived from peripheral blood monocytes were incubated in full nutrient medium (Full), Full with 50 μg/mL rapamycin (Full+Rapa), or AA starvation medium (Starve) for 4 h. Some of the Starve-treated macrophages were then recovered in Full nutrient medium for 2 h (Recover). The level of autophagosome formation was detected using a commercial CAT dye (Cyto-ID Green autophagosome dye) (Chan et al., 2012). We observed that Full+Rapa- and Starve-treated macrophages exhibited significantly elevated autophagy than the Full-treated macrophages, while Recover-treated macrophages presented intermediate level of autophagy (Fig. 1A, B). These results were consistent with previous discoveries, which validated our experimental procedure for autophagy detection (Chan et al., 2012). We then treated macrophages with live M. leprae or heat-killed M. leprae in Full medium. Macrophages in full nutrient medium treated with live M. leprae (Full+Live) demonstrated significantly elevated autophagy level than macrophages without live M. leprae (Full) (Fig. 1C). Macrophages in full nutrient medium treated with killed M. leprae (Full+Killed) presented intermediate autophagy level between Full-treated macrophages and Full+Live-treated macrophages, but the differences were not statistically significant.
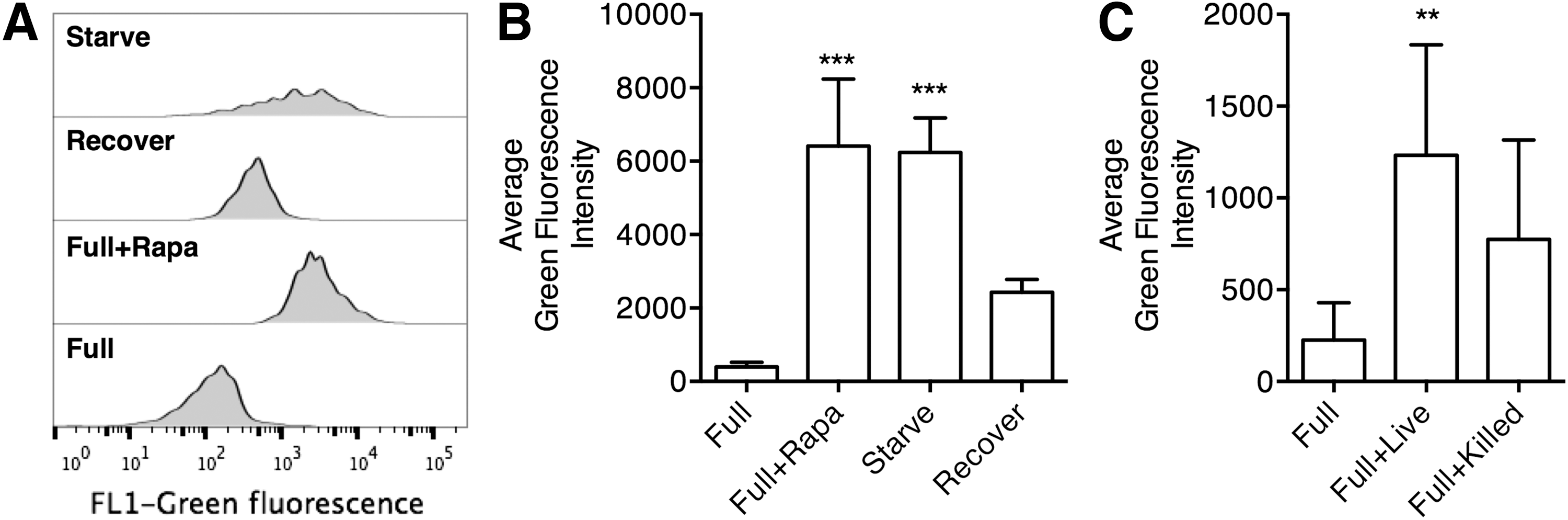
Autophagy induction in Mycobacterium leprae-treated macrophages.
The level of autophagy was associated with the mode of macrophage activation
We previously demonstrated that macrophages treated with live M. leprae presented different cytokine secretion and antigen-presentation function from macrophages treated with killed M. leprae, but the underlying mechanisms were unknown (Yang et al., 2016). Here, we investigated whether this difference was associated with autophagy. Macrophages were first incubated in full medium as a control or in starve medium for 4 h to induce autophagy, and then incubated without M. leprae (Full/Starve), with live M. leprae (Full+Live/Starve+Live), or with killed M. leprae (Full+Killed/Starve+Killed) for an additional 24 h in full nutrient medium. The cytokine secretion in the supernatant was examined first (Fig. 2). Consistent with our previous findings (Yang et al., 2016), Full+Killed macrophages presented higher IL-1β, IL-6, IL-12, and TNF-α secretion than Full macrophages, while Full+Live macrophages presented lower IL-1β and TNF-α and higher IL-10 secretion than Full macrophages.
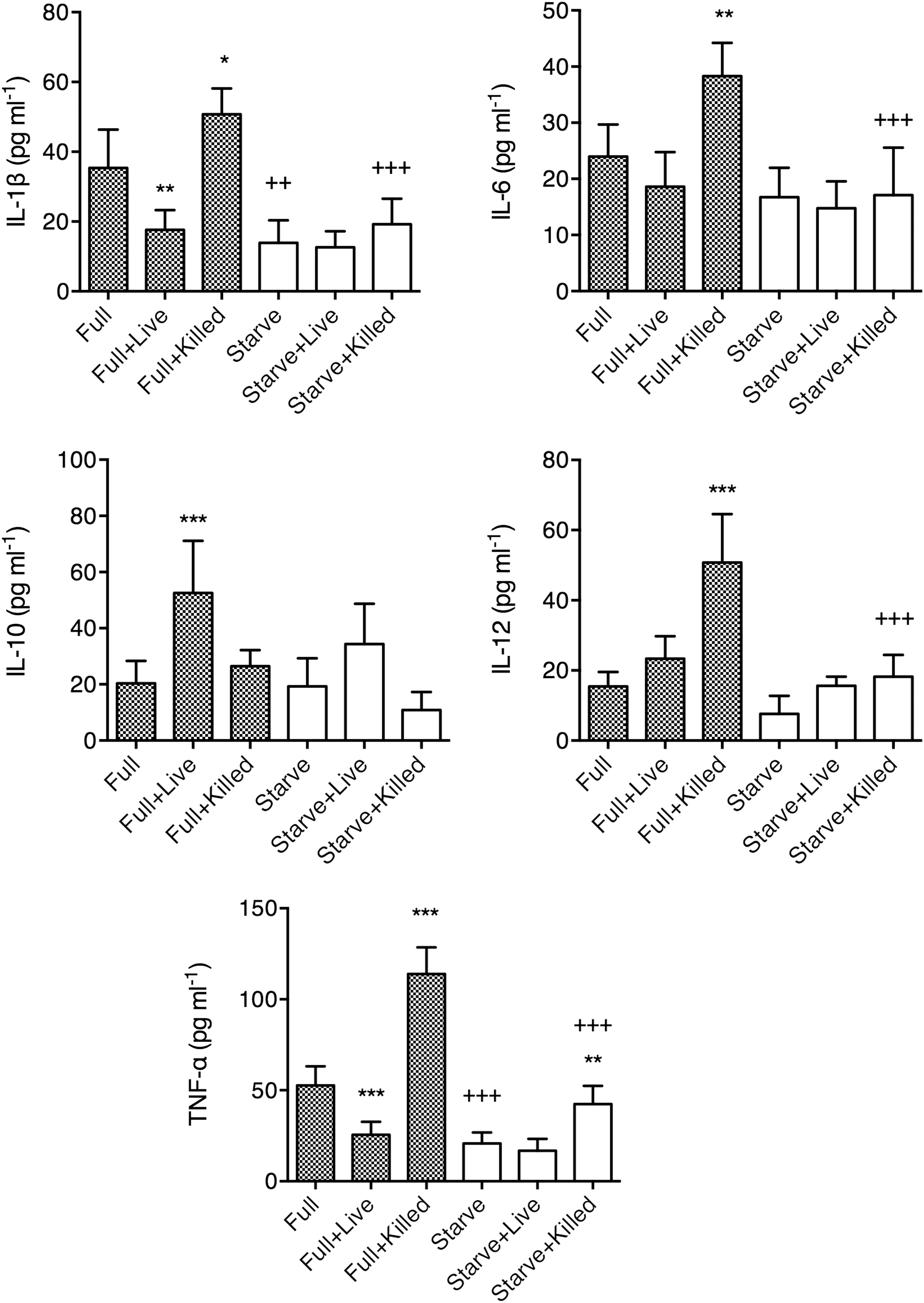
Secretion of cytokines by macrophages under Full or Starve conditions, with various M. leprae treatment. Peripheral blood monocyte-derived macrophages were cultured in Full medium without M. leprae (Full), with live M. leprae at 20:1 MOI (Full+Live), or with killed M. leprae at 20:1 MOI-equivalent (Full+Killed), or in Starve medium without M. leprae (Starve), with live M. leprae at 20:1 MOI (Starve+Live), or with killed M. leprae at 20:1 MOI-equivalent (Starve+Killed), at 5 × 105 macrophages/mL at the bottom of a 3.0-μm Transwell-24 plate. Cytokine capture beads were placed at the top and the cytokine secretion was measured by Luminex. Macrophages from six different individuals were tested, in triplicates. Results were presented as mean ± standard deviation, and tested by one-way ANOVA followed by Sidak's test. Statistical significance between groups with the same medium (i.e., Full or Starve) was denoted by asterisks, while statistical significance between groups with the same M. leprae treatment (i.e., none, Live, or Killed) was denoted by plus signs. *p < 0.05. **/++ p < 0.01. ***/+++ p < 0.001.
The results between Full medium and Starve medium treatments were also compared. Starve-treated macrophages presented lower IL-1β and TNF-α secretion than Full-treated macrophages. Also, Starve+Killed-treated macrophages presented lower IL-1β, IL-6, IL-12, and TNF-α secretion than Full-Killed-treated macrophages. In contrast, no significant difference in IL-10 secretion by macrophages was found between Full and Starved treatments with matching types of M. leprae stimulation. To examine whether the downregulation of proinflammatory cytokines was associated with autophagy, we calculated the difference in proinflammatory cytokine secretion and the difference in autophagy between the corresponding Full and Starve conditions, and examined whether there was any association between the two measures. Indeed, we observed that higher autophagy levels predicted further reductions in all proinflammatory cytokine secretions (Fig. 3), which demonstrated that increase in autophagy could result in macrophages with reduced capacity to secrete proinflammatory cytokines.
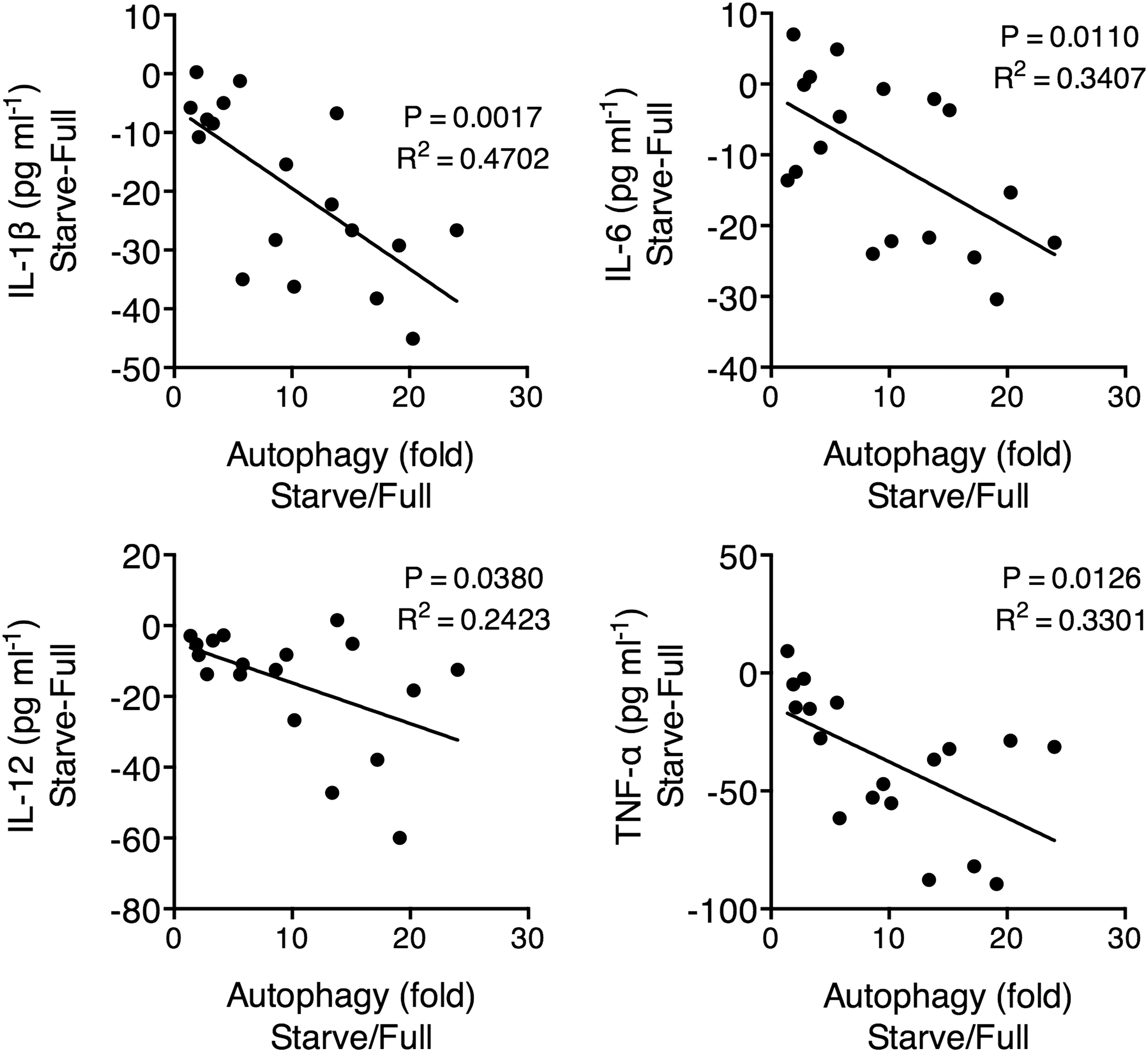
Correlation between the level of autophagy and the cytokine secretion in macrophages. After M. leprae treatment, macrophages were harvested for autophagy detection by flow cytometry using CAT dye. The increase in autophagy level was expressed as the quotient between the corresponding Full and Starve conditions, using the formula (average green fluorescence intensity in Starve conditions)/(average green fluorescence intensity in Full conditions). The change in cytokine secretion was expressed as the difference between the corresponding Full and Starve conditions, using the formula (average green fluorescence intensity in Starve conditions) − (average green fluorescence intensity in Full conditions). Macrophages were from six individuals with each examined under three M. leprae treatment conditions, resulting in 18 data points. Pearson's correlation test. A linear fit line is drawn.
Starve-treated macrophages helped the priming of anti-inflammatory T cell responses
We previously observed that live M. leprae-treated macrophages preferentially primed anti-inflammatory T cell responses, characterized by low IFN-γ and high IL-10 secretion, elevated Foxp3 expression, and reduced cytotoxicity (Yang et al., 2016). Here, we examined the effect of autophagy in the T cell priming function of macrophages. After incubating the macrophages under Full or Starve conditions and stimulating with various types of M. leprae, the CD45RA+ T cells were added in the macrophage culture. After 6 days, the T cells were negatively purified and restimulated by anti-CD3/CD28 and the secretion of IFN-γ, IL-10, granzyme B, and perforin was assessed after 12 h. We observed that the presence of macrophages was necessary for the induction of significant amount of IFN-γ, IL-10, granzyme B, and perforin from T cells, since little secreted cytokines and cytolytic molecules were found in naive T cells alone or naive T cells with Live or Killed M. leprae only cultures (Fig. 4). Compared to T cells cocultured with Full-treated macrophages, T cells cocultured with Full+Killed-treated macrophages secreted significantly higher IFN-γ (Fig. 4A), while T cells cocultured with Full+Live-treated macrophages secreted significantly higher IL-10 (Fig. 4B).
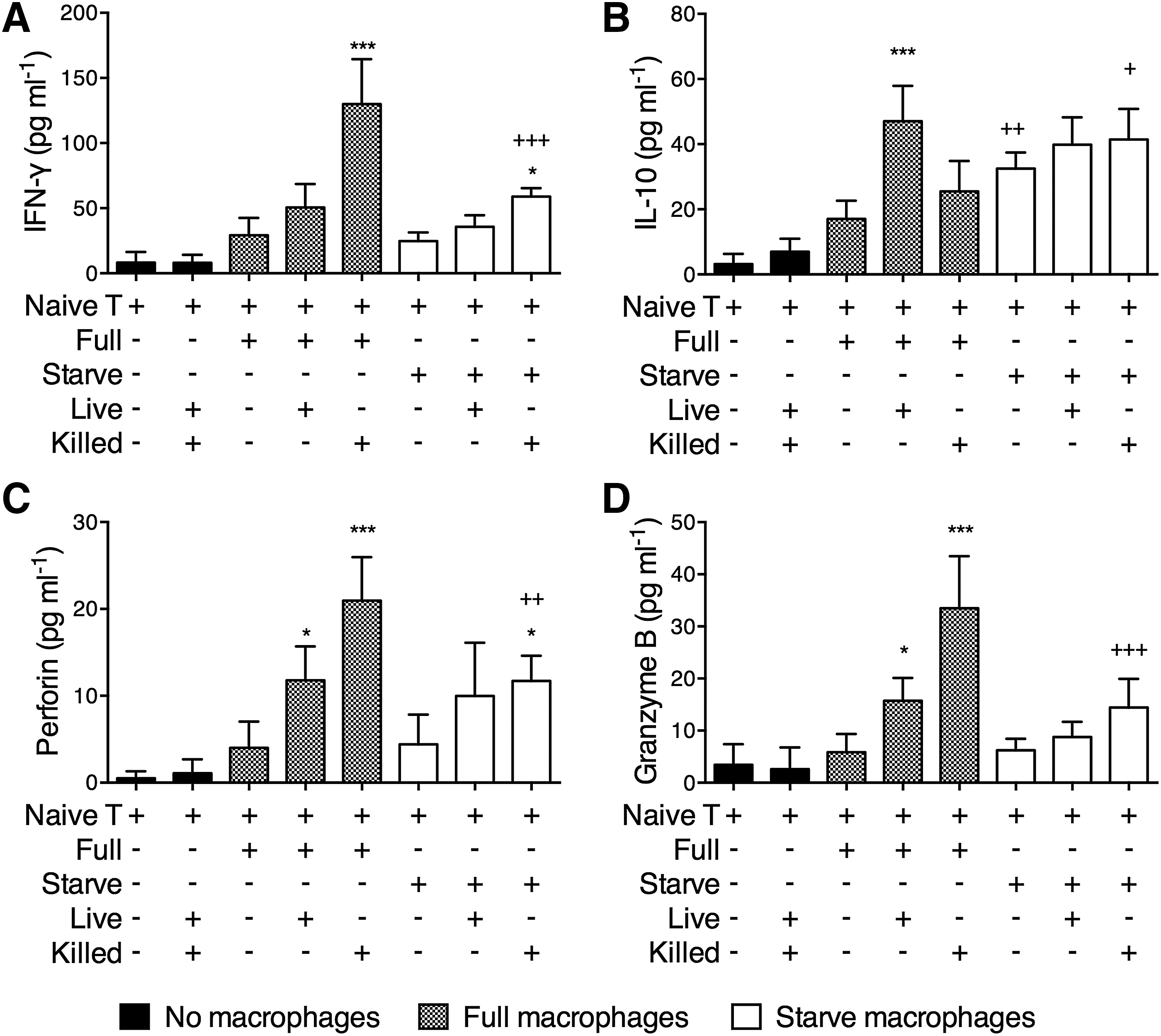
Priming of T cells by Full- or Starve-treated macrophages. Naive CD3+CD45RA+ T cells were incubated alone, with Live and Killed M. leprae, or with autologous Full-, Full+Live-, Full+Killed-, Starve-, Starve+Live-, and Starve+Killed-treated macrophages. The ratio of T cell: macrophage was 1:1. M. leprae was added at an MOI or MOI-equivalent of 20:1. After 6 days, T cells were purified and restimulated with 2 μg/mL anti-CD3 (OKT3) and anti-CD28 (CD28.2) at the bottom of a transwell-24 plate, while cytokine capture beads were placed at the top. After 12 h, the expression of
In addition, several differences in T cell cytokine secretion were observed if the macrophages used in coculture were pretreated with Starve medium instead of Full medium. Replacing Full-treated macrophages with Starve-treated macrophages resulted in significantly higher IL-10 production in T cells (Fig. 4B). Also, replacing Full+Killed macrophages with Starve+Killed macrophages resulted in significantly lower IFN-γ and significantly higher IL-10 production in T cells (Fig. 4A, B). Both Full+Live-treated macrophages and Full+Killed-treated macrophages stimulated significantly higher granzyme B and perforin expression from T cells than Full macrophages, with Full+Killed-treated macrophages being more effective (Fig. 4C, D). Replacing Full+Killed-treated macrophages with Starve+Killed macrophages significantly reduced the level of granzyme B and perforin expression from T cells.
Skewing of T cells affected the efficiency of autophagy
Autophagy could act as a defense mechanism by directly destroying intracellular mycobacterium (Gutierrez et al., 2004), but a Th2 (IL-4 and IL-13)-dominant immune response could abrogate this process (Harris et al., 2007). Here, we examined whether autophagy of live M. leprae was affected by the skewing of T cell responses. Naive T cells were incubated with Full-Killed-treated macrophages or Starve-killed-treated macrophages for 6 days, and then harvested by negative selection to obtain IFN-γ-high, granzyme B-high and perforin-high proinflammatory T cells (Full-Killed T, shown in Fig. 4A–D, column 5), or IL-10-high, granzyme B-low and perforin-low anti-inflammatory T cells (Starve-Killed T, shown in Fig. 4A–D, column 8), respectively.
The Full+Killed T cells and Starve+Killed T cells were then added to macrophages, without or with Live M. leprae for 24 h. The level of autophagy in the macrophages was then examined. We found that high concentration of Full+Killed T cells enhanced autophagy in Live M. leprae-treated macrophages, while high concentration of Starve+Killed T cells suppressed autophagy in Live M. leprae-treated macrophages (Fig. 5A). To confirm that the efficiency autophagy by macrophages was altered by Full+Killed T cells and Starve+Killed T cells, we examined the viability of M. leprae. At the end of incubation, Live M. leprae-treated macrophages were lysed and the intracellular M. leprae were harvested. We found that compared to the viability in extracellular M. leprae, the viability in macrophages treated with Full+Killed T cells was significantly reduced, an observation absence in macrophages treated with Starve+Killed T cells (Fig. 5B). To examine whether IL-10 inhibited autophagy in M. leprae-treated macrophages and allowed survival of intracellular M. leprae, we blocked IL-10 action with a neutralizing antibody (IL-10 Neut. mAb). Compared to the isotype control antibody, the IL-10 Neut. mAb in the Starve+Killed T cell+Live M. leprae-treated macrophage coculture resulted in significantly higher autophagy and lower M. leprae viability (Fig. 5C).
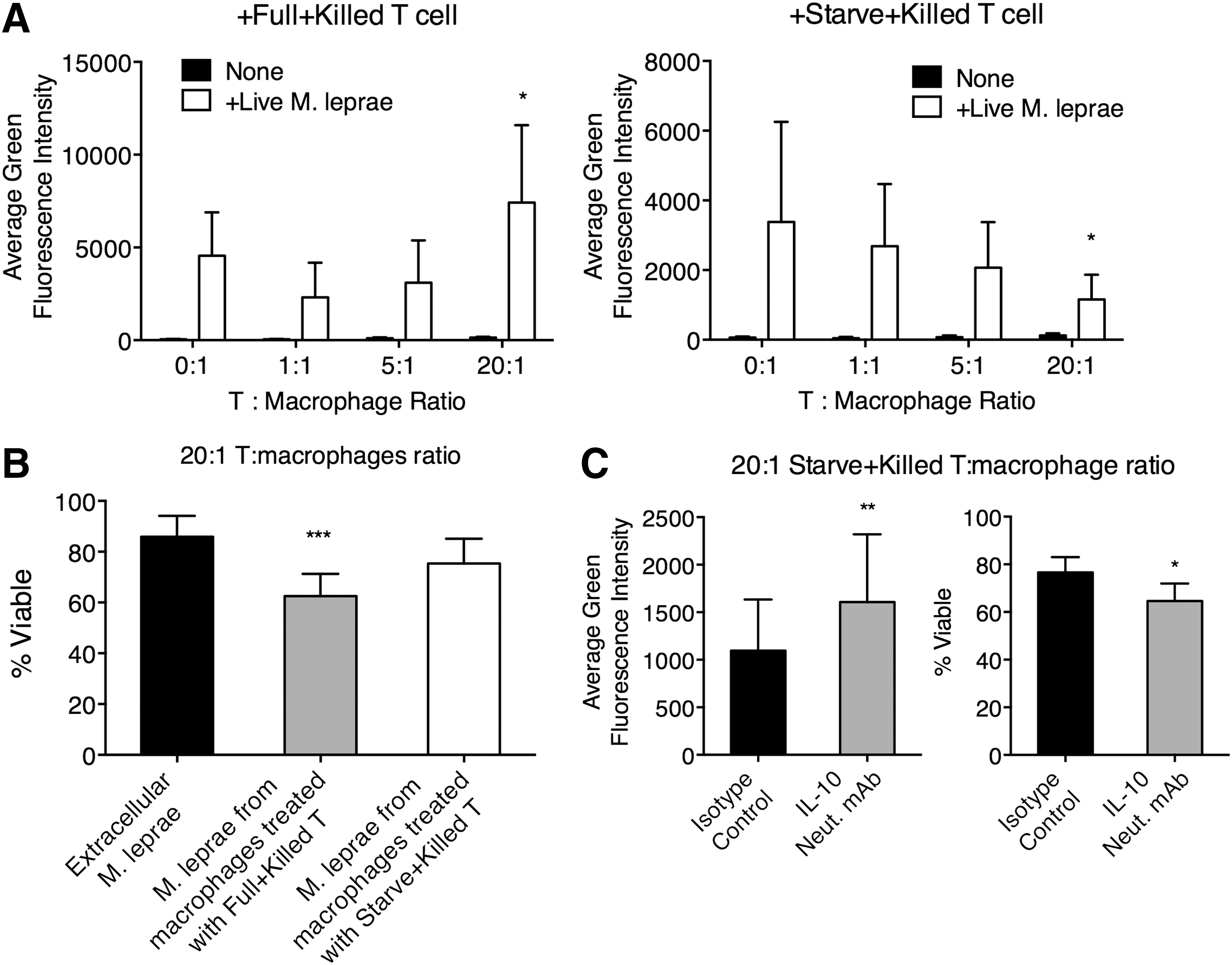
The efficiency of autophagy in Live M. leprae-treated macrophages was affected by the types of T cell responses. Naive T cells primed by Full+Killed or Starve+Killed macrophages, and then purified from the T cell-macrophage coculture so that proinflammatory Full+Killed T cells and anti-inflammatory Starve+Killed T cells were obtained. The T cells were then added at various ratios to macrophages without or with Live M. leprae (MOI 20:1) for 24 h.
Discussion
In this study, we examined the effect of macrophage autophagy in M. leprae infection. Because the CAT dye could directly stain autophagosomes in live cells without the necessity of LC3-GFP transfection, and flow cytometry could analyze higher cell numbers than fluorescent microscopy, we used CAT dye coupled with flow cytometry to examine autophagy in macrophages. We found that the level of autophagy was significantly higher in live M. leprae-treated macrophages than in resting macrophages, while the level of autophagy in killed M. leprae-treated macrophages was not significantly different from that in resting macrophages.
We previously observed that treatment of macrophages with live or killed M. leprae resulted in different macrophage differentiation (Yang et al., 2016). To examine whether the difference in autophagy induction played a role, we first investigated the macrophage differentiation under different levels of autophagy induction. AA starvation was commonly used to artificially induce autophagy in Mycobacterium infection (Gutierrez et al., 2004; Harris et al., 2007; Deretic et al., 2015), and was demonstrated here to effectively induce autophagy. Therefore, we incubated macrophages in full nutrient medium, which did not induce high autophagy levels by itself, or in AA starvation medium, which significantly elevated the formation of autophagosomes. The live or killed M. leprae were then added to the macrophages. In full nutrient medium, killed M. leprae preferentially induced proinflammatory IL-1β, IL-6, IL-12, and TNF-α, while live M. leprae preferentially induced anti-inflammatory IL-10. Interestingly, the macrophages in AA-starvation medium presented significantly altered cytokine expression profile. When no M. leprae was added, AA-starvation medium alone led to reduced IL-1β and TNF-α production from macrophages. Moreover, AA-starvation medium had reverted the proinflammatory effects of killed M. leprae. On the other hand, AA-starvation medium did not significantly change the cytokine secretion profile in live M. leprae-treated macrophages. Together, these data demonstrated that Starve treatment of macrophages resulted in significantly less proinflammatory cytokine secretion. Since in our experimental system, the macrophages were only deprived of AA for the first 4 h in a total of 4 + 24 h of incubation, this is unlikely a result of less protein synthesis due to starvation. Indeed, the secretion of IL-10 by macrophages in Starve treatments was not significantly decreased.
It was previously demonstrated that autophagy suppresses inflammasome activation by clearing endogenous NLRP3 inflammasome agonists, such as reactive oxygen species and mitochondrial DNA and degrading components of the inflammasomes, and pro-IL-1β (Harris et al., 2011; Nakahira et al., 2011; Chuang et al., 2013; Deretic et al., 2015). Several autophagy proteins were demonstrated to suppress type I IFN responses and downregulate NF-κB signaling (Deretic et al., 2015). Therefore, a possible explanation to the starvation-induced changes in the cytokine expression profile is that AA starvation before M. leprae treatment induced autophagy in macrophages, which favored the differentiation of macrophages toward the regulatory types and overrode the effects of M. leprae. Here, we observed that the increase in autophagy by switching to Starve medium predicted lower levels of secreted proinflammatory cytokines. These results, together with earlier discoveries, supported the idea that autophagy possessed an anti-inflammatory role.
We next examined the effects on autophagy on the T cell priming function of macrophages. In full medium, killed M. leprae-treated macrophages primed strong IFN-γ, perforin, and granzyme B production from T cells. However, in AA starvation medium, the T cell IFN-γ, perforin, and granzyme B production primed by killed M. leprae-treated macrophages was significantly reduced, while the T cell IL-10 production was increased, demonstrating that AA starvation had significantly changed the T cell priming function of killed M. leprae-treated macrophages. Interestingly, we observed that IL-10-high T cells primed by Starve-Killed macrophages could reduce autophagy in Live M. leprae-treated macrophages, while IFN-γ-high T cells primed by Full+Killed macrophage priming promoted autophagy in Live M. leprae-treated macrophages. This inhibition of autophagy by IL-10-high T cells was associated with the absence of killing of intracellular M. leprae in macrophages. Indeed, inhibition of IL-10 resulted in significantly elevated autophagy level and significantly reduced intracellular M. leprae survival in macrophages cocultured with IL-10-high T cells. Comparing the autophagy levels in the IFN-γ-high T cell+macrophage coculture (Fig. 5A, last column) and in the IL-10-high T cell+macrophage+IL-10 neutralization coculture (Fig. 5C, left panel, 2nd column), we found that the neutralization of IL-10 could not fully increase the autophagy level in macrophages, suggesting that T cell-mediated mechanisms other than IL-10 was involved. Overall, these results demonstrated that low macrophage autophagy and anti-inflammatory T cell responses resulted in improved intracellular M. leprae survival, which could contribute to the persistence of the pathogen and development of leprosy. Interestingly, it was demonstrated that the human hepatitis B virus (HBV) activated the host's autophagy machinery, which promoted HBV DNA replication (Sir et al., 2010; Li et al., 2011), but the underlying mechanism is unclear.
Here, our study demonstrated that although autophagy could promote the clearance of intracellular pathogens, it also induced anti-inflammatory T cell responses, which inhibited further autophagic processes and promoted the persistence of M. leprae.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (81470144), and Shanghai Municipal Training Project of Excellent Young Medical Talents (XYQ2011005).
Authors' Contributions
Conceived and designed the experiments: Y.M., Y.L., D.Y. Performed the experiments: Y.M., L.Z., J.L., T.S., J.C., J.U.Y. Analyzed the data: J.O.Y., Y.L., D.Y. Contributed reagents/materials/analysis tools: J.O.Y., Y.L., D.Y. Wrote the paper: Y.M., Y.L., D.Y.
Disclosure Statement
No competing financial interests exist.