
Abstract
The underlying pathogenic lesions of glycogen storage disease type II (GSD II) and the diversity of this disease among different species are still under exploration. Thus, we created an acid alpha-glucosidase (gaa) gene-mutated zebrafish model of GSD II and examined the sequential pathogenic changes. gaa mRNA and protein expression, enzymatic activity, and lysosomal glycogen accumulation were assessed, and the phenotypic changes were compared between wild-type (WT) and gaa-mutated zebrafish. The presence of a Δ13 frameshift mutation in the gaa gene was confirmed at both the DNA and transcribed mRNA levels by Sanger sequencing. The relative amount of gaa mRNA was decreased before 2 days postfertilization (dpf), after which it unexpectedly increased in the mutant compared with the WT zebrafish. Consistent with the mRNA expression, the Gaa enzymatic activity in the mutant was downregulated before 3 dpf, while the Gaa protein level was slightly decreased at 4 dpf and was maintained at a consistent level in the adult gaa mutant muscle tissue. However, more than half of the adult mutant zebrafish exhibited excessive glycogen accumulation in the liver and muscles, along with the presence of autophagosomes, as determined by transmission electron microscopy. Thus, we have successfully generated a frameshift mutation in the gaa gene in zebrafish. The unique gaa gene expression changes and mild GSD II features during the adult stage strongly indicate the existence of species-specific differences, as well as an underlying compensatory network, which may warrant further examination.
Introduction
G
GAA knockout (GAA-KO) mice (Pompe model mice) have been widely used to study the pathology of and evaluate different therapies for GSD II since the establishment of this model by Raben et al. (1998). While the pathogenesis is not completely consistent between patients with Pompe disease and GAA-KO mice (Lim et al., 2014), studies of Pompe models in other species are still necessary. In this study, we aimed to mutate the gaa gene (XM_001921922) in zebrafish (Danio rerio) (Nüsslein-Volhard, 2012) and to characterize this novel mutant's phenotype.
Materials and Methods
Zebrafish maintenance and experimental procedures
Zebrafish of the AB strain were maintained at 28°C ± 0.5°C under a 14:10-h light:dark cycle according to the standard protocol. Feeding was initiated at 5 days postfertilization (5 dpf) and was performed twice a day. Invasive procedures were performed after anesthetization of the fish using tricaine methanesulfonate as previously described (Kaufmann et al., 2012). This study was approved by the faculty of the Ethics Committee of Children's Hospital of Fudan University.
Generation and synthesis of TALEN mRNA
Targeted gene disruption of the first exon of gaa in zebrafish was performed using the following transcription activator-like effector nucleases (TALEN) DNA binding sites: left: 5′-TGTGTACTTCTTATCAT-3′ and right: 5′-GCAACACAAATAGTGT-3′. TALEN plasmids were constructed with a pCS2-eTALEN-T backbone using a TALE Toolkit (ViewSolid, China). After in vitro transcription of TALENs using an mMESSAGE mMACHINE® SP6 Transcription Kit (Ambion, USA), conducted according to the manufacturer's instructions, TALEN mRNAs were purified using the phenol–chloroform method, resuspended at a concentration of 100 μg/μL in nuclease-free H2O, and stored at −80°C before microinjection into zebrafish embryos.
Microinjection of zebrafish embryos
TALEN mRNAs were injected into zebrafish embryos at the one- or two-cell stage (50 pg of each TALEN mRNA per embryo) using a microinjector (Picospritzer III, Parker, NJ). The embryos were raised in E3 egg water (containing sea salts in distilled water), which was changed every 24 h before 6 dpf.
DNA extraction and PCR for digestion and sequencing
We verified gaa mutagenesis by the sequencing of genomic DNA from lysates of zebrafish embryos (5–6) at 2 dpf or from fin clips of adult zebrafish. The lysates were prepared by incubation of either zebrafish embryos or fin clips in 50 μL of 50 mM NaOH at 95°C for 20 min, followed by addition of 1 M Tris–HCl (pH 8.0). Next, the lysates were centrifuged at 12,000 rpm for 5 min, and the supernatants (genomic DNA) were collected and used as templates for PCR amplification of the target region of the gaa gene (primers: F: 5′-TATGAGGCAATAAATTTCCG-3′ and R: 5′-TGATCTCTAGGAACAGTTGAT-3′). In addition, the sequences of the PCR primers used for cDNA are shown in Supplementary Table S1a (Supplementary Data are available online at
Luciferase single-strand annealing recombination assay
The luciferase activity generated during DNA repair induced by double-strand breaks was assessed to determine the efficiency of TALEN-mediated mutagenesis in vitro. For every 200 ng of TALEN expression plasmids, we cotransfected 50 ng single-strand annealing (SSA) reporter plasmid and 10 ng Renilla plasmid using Lipofectamine™ 2000 (Invitrogen, USA) into HEK293T cells in 24-well plates (Renilla luciferase driven by the β-actin promoter was used as a negative control). The cells were prepared using the Luciferase Cell Lysis Buffer (NEB, USA) at three days after transfection, and relative luciferase activity was measured using a Dual-Luciferase Assay System (Promega, USA) with a SYNERGYMx Luminescence Microplate Reader (BioTek, USA).
Whole-mount in situ hybridization
Digoxigenin antisense probes (Roche, Austria) were synthesized for the detection of gaa transcripts within cDNA at 24-h postfertilization (hpf) of the zebrafish embryos. The probe sequences were as follows: 5′-TGGAGCCTTTTACCCCTTCA-3′ (forward) and 5′-GATGGTGTCCAACGAAGCAG-3′ (reverse). In addition, the amplification conditions were as follows: 94°C for 5 min; 14 cycles at 94°C for 30 s, 64°C (with a decrease of 0.5°C every cycle) for 30 s, and 72°C for 40 s; 30 cycles at 94°C for 30 s, 57°C for 30 s, and 72°C for 40 s; and a final extension at 72°C for 7 min. Embryos and larvae collected from wild-type (WT) and gaa mutant zebrafish at different stages (24 hpf, 48 hpf, and 3 dpf) were fixed in 4% paraformaldehyde (PFA) at room temperature for 4 h, dehydrated with methanol, and stored at −20°C before in situ hybridization. Whole-mount in situ hybridization (WISH) was performed as previously described (Thisse and Thisse, 2008), and images were captured with an OLYMPUS DP73 stereomicroscope equipped with cellSens software (OLYMPUS, Japan).
Real-time PCR
Total RNA was isolated from the zebrafish samples using the TRIzol reagent (Ambion, USA), and it was then reversed transcribed using a PrimeScript cDNA Synthesis Kit (Takara, Japan). The sequences of the primers used for gaa gene amplification were as follows (5′ to 3′): F-AGTCAGCGAGCCCAGGAT and R-CCAACGAAGCAGGAAACA. In addition, the housekeeping gene β-actin was amplified using the following primers (5′ to 3′): F-ACGAACGACCAACCTAAACTCT and R-TTAGACAACTACCTCCCTTTGC. The cycling program was 95°C for 1 min, followed by 40 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s. The primers used to identify the mRNA alternative splicing isoforms before (GB) and after (GA) targeted mutagenesis sequence are listed in Supplementary Table S1b.
Western blotting
Samples (10–20 larvae or 300 mg whole tissue) were homogenized in RIPA buffer supplemented with protease inhibitor (Thermo, USA), and then, the proteins in each sample were separated by SDS-PAGE and transferred to nitrocellulose membranes. A GAA polyclonal antibody (rabbit, 1:200; Proteintech), β-actin polyclonal antibody (mouse, 1:2000; Well), goat anti-rabbit secondary antibody (1:5000; Well), and goat anti-mouse secondary antibody (1:5000; Well) were used to visualize protein bands with an LAS400 imaging system (FUJI, Japan).
Gaa enzymatic activity assay
Gaa activity in the zebrafish was determined by measuring the production of 4-methylumbelliferone from an artificial substrate, 4-methylumbelliferyl α-
Periodic acid–Schiff staining of adult zebrafish
Adult zebrafish bodies (2/3 of the whole body) were fixed in 4% PFA for 48 h at room temperature. The fixed zebrafish were then paraffin embedded and sectioned at 5 μm for periodic acid–Schiff (PAS) staining. Then, the sections were deparaffinized and rehydrated in serially diluted alcohol and deionized water. Next, the slides were placed in a 0.5% periodic acid solution for 5 min and then rinsed in distilled water, followed by Schiff (15 min) and hematoxylin (1 min) staining. Finally, the slides were dehydrated, mounted with xylene, sealed with neutral gum, and visualized under a light microscope.
Transmission electron microscopy examination
Tissue specimens (<1 mm3) were obtained from zebrafish skeletal muscles using a razor blade. All of the fragments were fixed with 2.5% glutaraldehyde in 0.1 M phosphate-buffered saline (PBS) for 2 h or more and washed in PBS three times for 15 min each. Subsequently, the samples were postfixed in 1% osmium tetroxide for 2 or 3 h and washed again in PBS 3 times. After the samples were serially dehydrated and embedded overnight, the blocks were sectioned (70 nm) using an ultramicrotome for preparation for uranyl acetate and lead citrate staining. Finally, the sections were examined at 80 kV with an electron microscope (FEI, Czech Republic).
Statistical analyses
All data are expressed as the mean ± standard error of the mean (SEM) and analyzed by Student's t-test or one-way ANOVA, followed by Tukey's multiple-range test. For all statistical analyses, a P < 0.05 was considered statistically significant.
Results
gaa mRNA expression at embryonic and adult stages
WISH was performed on WT zebrafish at 24 hpf, 48 hpf, and 3 dpf, and widespread staining of the gaa transcript was detected, suggesting that gaa was widely expressed throughout the embryo during the early stages of the development (Fig. 1A). gaa mRNA expression increased until 3 dpf and decreased thereafter in the whole larvae harvested at 24 hpf, 48 hpf, 3 dpf, and 5 dpf, as determined by real-time PCR (Fig. 1B), and in the adult tissues, gaa was expressed at the highest level in the brain and at the lowest level in the heart (Fig. 1C).
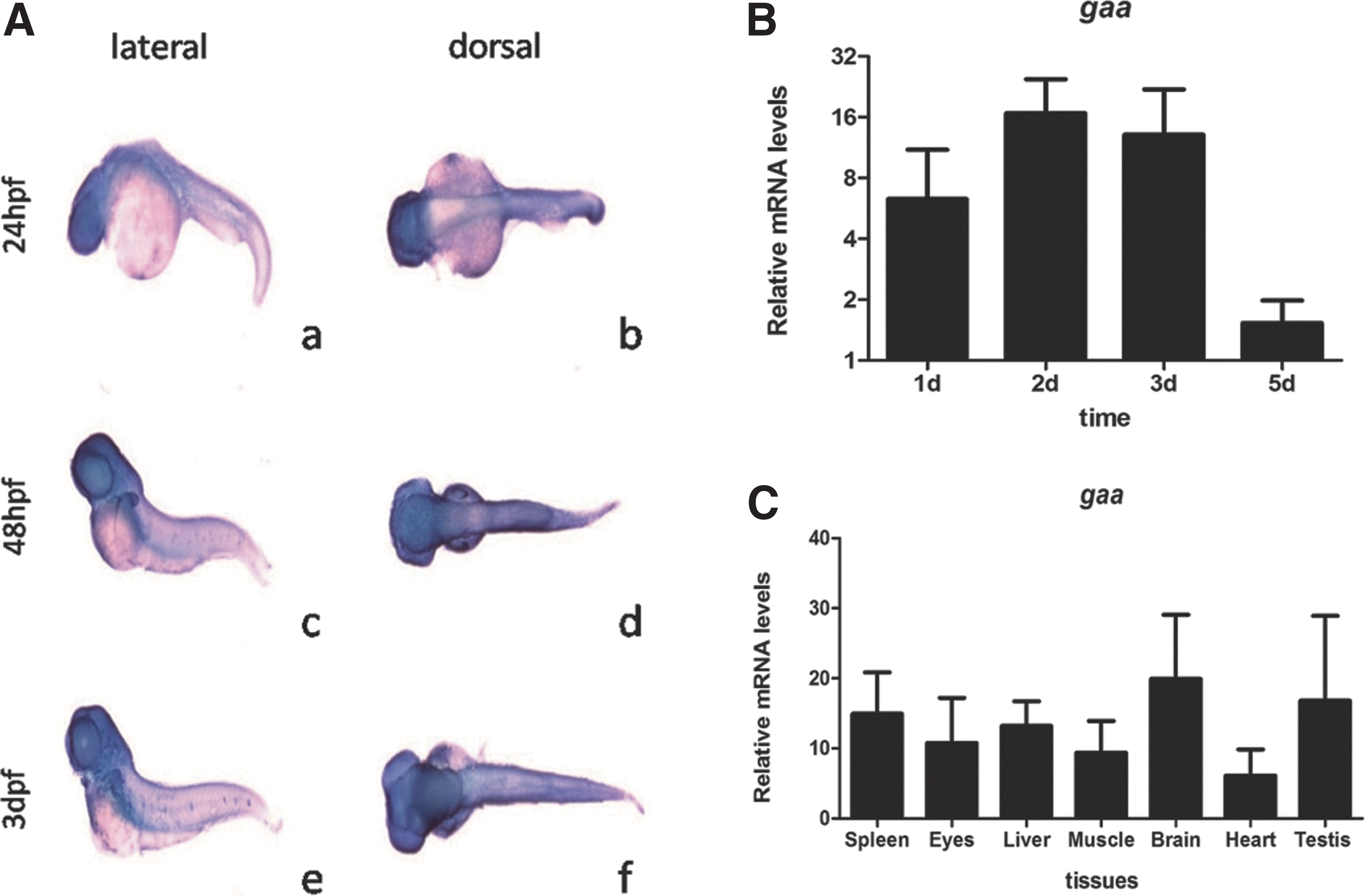
gaa expression in WT zebrafish.
Generation of gaa zebrafish mutants
TALENs targeting exon 1, located at c.60–73 nt after the start codon of the zebrafish gaa gene, were designed as illustrated in Figure 2A. The in vitro transcribed TALEN mRNAs were injected into one-cell stage embryos to generate genetic mutants (F0). The efficiency of the synthesized TALENs was double verified, first by the direct sequencing of genomic DNA extracted from every 5 F0 zebrafish embryos at 48 hpf (Supplementary Fig. S1A) and second by enzymatic Bbs I digestion. Samples with the mutation were resistant to digestion, while the WT samples were sensitive to digestion and yielded 386- and 114-bp cleavage products (Fig. 2B). Furthermore, assessment of relative luciferase activity, reflecting in vitro efficiency, revealed that the activity was 41 times higher in the gaa-TALEN-transfected cells than in the untransfected controls (Fig. 2C).

The target site and efficacy evaluation of gaa TALEN mRNAs.
After verifying the efficiency of TALEN-mediated mutagenesis, TALEN mRNAs were injected into one-cell stage embryos to obtain F0 mosaic zebrafish, which were outcrossed with WT zebrafish for the further generation of heterozygous gaa mutant offspring (F1). The gaa heterozygous mutant zebrafish were identified through fin clip genotyping, and the homozygous mutants (F3 or F4 offspring) were screened by the sequencing of offspring generated by interbreeding with F2 or F3 heterozygous offspring.
A 13-bp deletion (c.58 A+c.61-72 GTGTATGAAGAC) in the zebrafish gaa gene was successfully introduced (Fig. 2D). Compared with the reference amino acid sequence (XP_001921957.2), the 13-bp deletion (Δ13) caused a frameshift mutation, resulting in the translation of a truncated Gaa protein containing the first 19 amino acids of Gaa, followed by 43 missense amino acids and a premature stop codon. Bioinformatics analysis predicted that the structure of the mutated protein was extremely different from that of the WT protein translated from the original ATG start site (Fig. 2E).
Alterations in Gaa expression and enzymatic activity in gaa mutants at early developmental stages
Real-time PCR was performed to assess gaa mutant zebrafish larvae at 22 hpf, 48 hpf, 3 dpf, and 5 dpf. Interestingly, gaa mRNA expression rapidly decreased in the mutants during the first 2 dpf and progressively increased thereafter, suggesting that the frameshift mutation may have affected the expression or stability of the gaa mRNA during early development of the gaa mutant zebrafish (Fig. 3A). Gaa protein expression was only slightly downregulated in the gaa mutant larvae at 4 dpf (P = 0.78, Fig. 3B, Supplementary Fig. S3, and the positive control can be seen in Supplementary Fig. S2), and in the normal controls, Gaa enzymatic activity increased from 2473 ± 51 (nmol/h/mg protein) to 4867 ± 129 (nmol/h/mg protein), while in the larvae with the gaa homozygous deletion, the activity increased from 762 ± 93 (nmol/h/mg protein) to 4831 ± 106 (nmol/h/mg protein) (Fig. 3C).
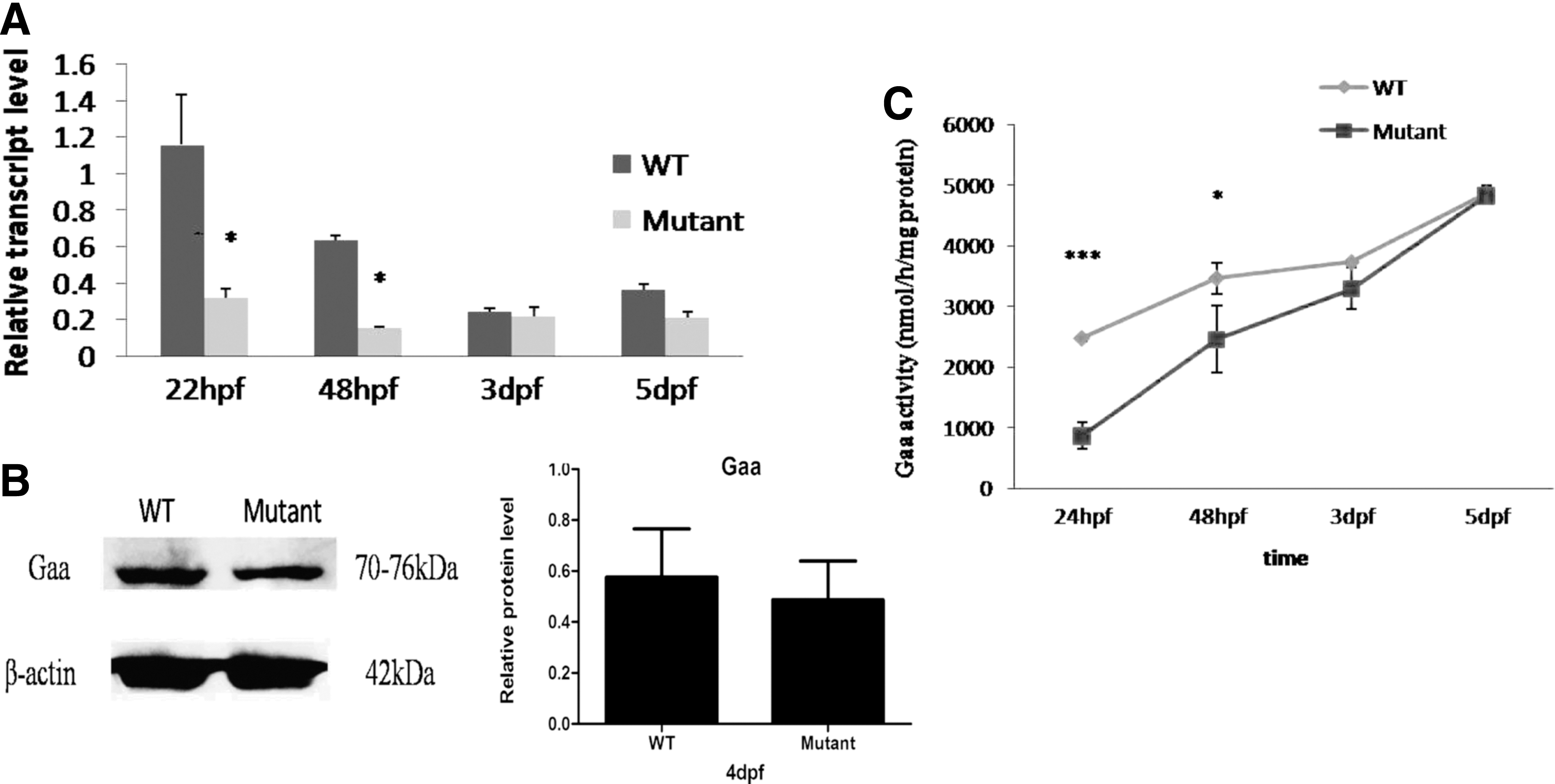
Comparison of gaa mutant zebrafish at early developmental stages between mutants and WT strain.
To further verify that targeted mutagenesis of the gaa mRNA was successful in the gaa mutants, gaa cDNA from the mutant zebrafish was sequenced, and the results confirmed that the same Δ13 deletion was present in the gaa mRNAs expressed in the mutants (Supplementary Fig. S1B). Furthermore, real-time PCR did not reveal the presence of any alternative splicing isoforms before or after the target sites in the gaa gene (Supplementary Fig. S1C). In addition, the Gaa protein level was slightly decreased in the gaa mutant zebrafish, suggesting that the protein was unstable. After ruling out the possibility of alternative transcript isoforms, we investigated whether there is a potential alternative translation start site in gaa that allows it to retain the functional domains by translation from a second start codon, although the length of the signal peptide would be truncated (Supplementary Fig. S1D).
Glycogen accumulation in adult gaa mutants
Because gaa expression and Gaa enzymatic activity in the gaa mutant zebrafish were only partially altered during the early developmental stages, we further evaluated whether the effects of gaa disruption persist in adult zebrafish. The viabilities of the mutant zebrafish were generally normal when they were raised into adulthood. PAS staining of the heart, liver, and skeletal muscles of the adult zebrafish revealed that mutants with a defective gaa gene exhibited the increased accumulation of glycogen granules (Fig. 4A–E). In accord with the abnormal glycogenolytic capacity, increased glycogen granule accumulation (P < 0.01) was detected in the gaa mutant zebrafish (39%, 61%, and 54% in the heart, liver, and muscle, respectively, n = 7) compared with the WT zebrafish (18%, 36%, and 25% in the heart, liver, and muscle, respectively, n = 7). In addition, Gaa enzymatic activity assay confirmed that the gaa mutants with increased glycogen accumulation (Fig. 4G) had significantly reduced enzymatic activity (667 ± 94 nmol/h/mg protein) compared with the WT zebrafish (937 ± 108 nmol/h/mg protein, p = 0.02). Furthermore, the Gaa protein level in the mutants with increased glycogen accumulation was lower (P < 0.05) than that in the WT zebrafish (Fig. 4H).
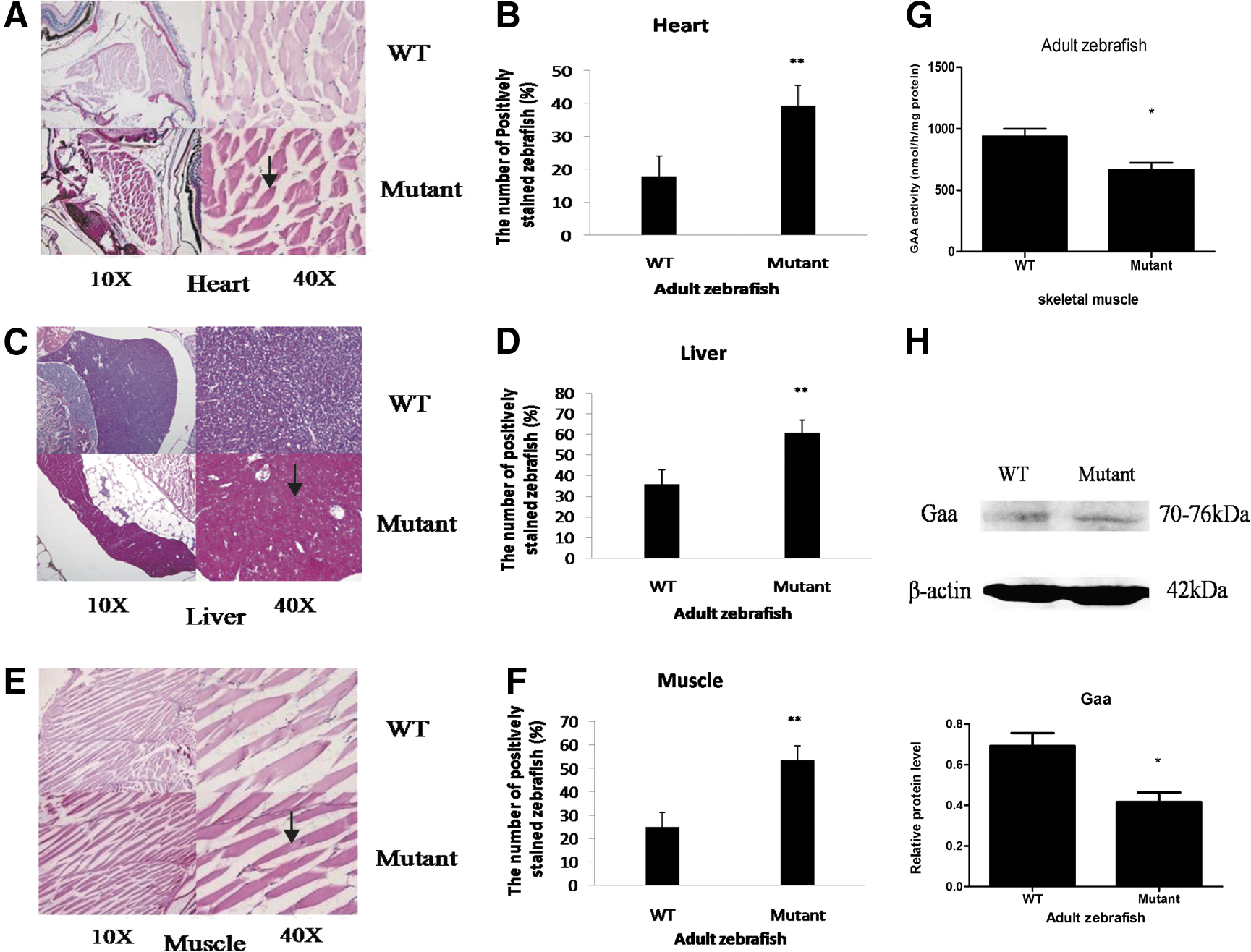
Gaa expression and enzymatic activity in adult zebrafish.
Skeletal muscle transmission electron microscopy (TEM) examination revealed that the gaa mutants possessed an increased amount of glycogen granules in the muscle tissues, particularly in the cytoplasm around the mitochondria and in autophagic vacuoles (Fig. 5A, B). In addition, autophagic multivesicular bodies and double-membrane autophagosomes filled with glycogen and other unknown debris were frequently observed in the mutants (Fig. 5A–C). In contrast, decreased glycogen accumulation and fewer autophagic vesicular structures were detected in the skeletal muscles of the WT controls (Fig. 5D–F).
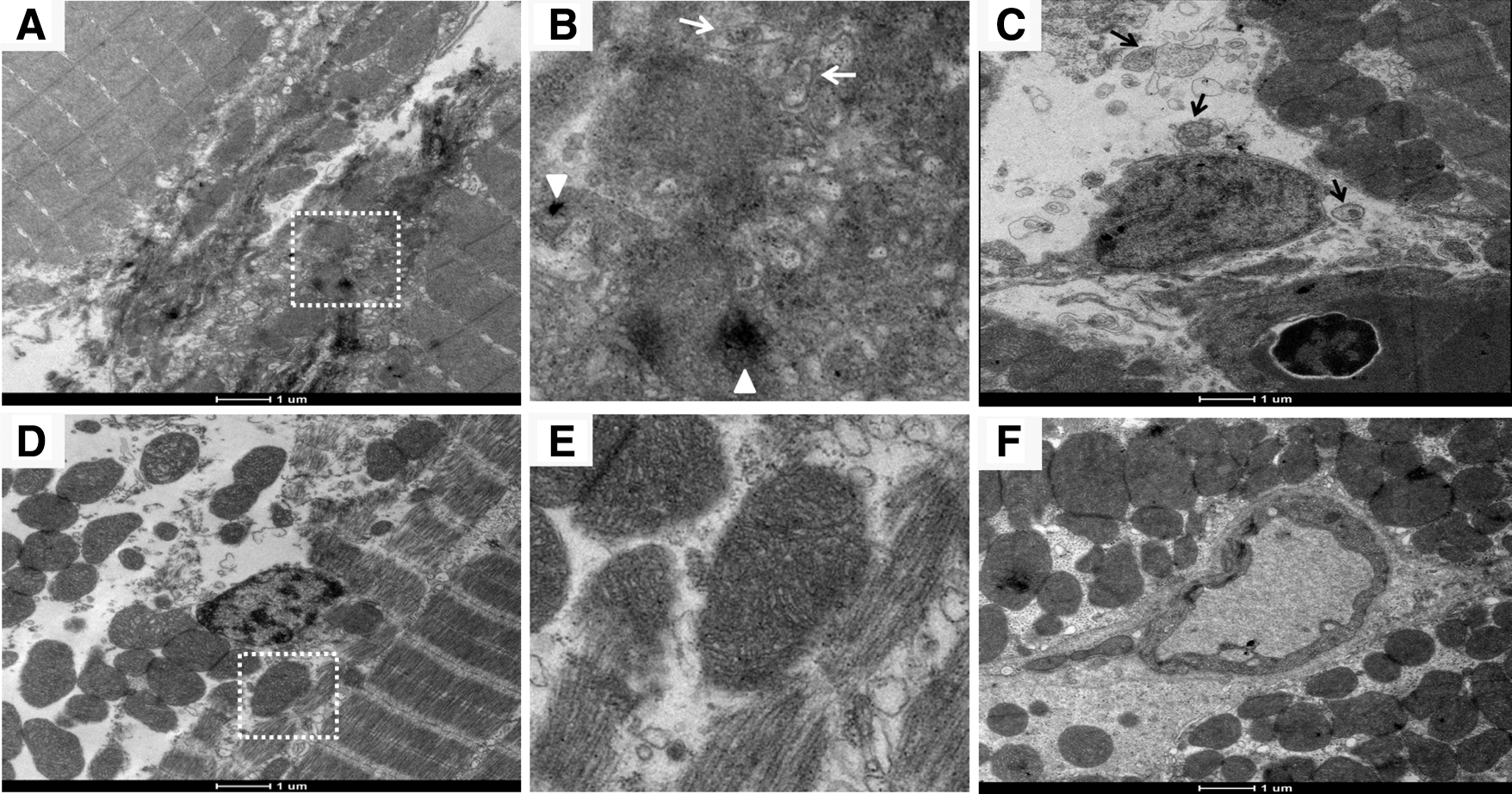
Transmission electron microcopy of skeletal muscles of adult gaa mutant and WT zebrafish.
Discussion
Recent high-resolution sequencing analysis has revealed the existence of highly similar, overlapping orthologous genes among humans, mice, and zebrafish (Howe et al., 2013). Due to its excellent reproductive ability and the optical clarity of its body, zebrafish is particularly suitable for genetic modification and massive drug screening studies (Nüsslein-Volhard, 2012). The current study showed that the gaa gene was widely expressed throughout the zebrafish embryo during the early stages of development, while in the adult zebrafish, it was expressed at the highest level in the brain and at the lowest level in the heart; its expression was also observed in the testes, eyes, liver, skeletal muscles, brain, and heart. These expression patterns are similar to those reported in mouse, quail, and human tissues (Bijvoet et al., 1998; Kikuchi et al., 1998; Raben et al., 2001).
A marked decrease in GAA activity is a common feature of both the infantile- and late-onset forms of Pompe disease (Steckel et al., 1982). As a secondary consequence of this decreased GAA activity, the widespread accumulation of glycogen has been observed in both human and GAA-KO mouse tissues (Van der Ploeg and Reuser, 2008; Turner et al., 2016). We successfully disrupted the gaa gene in zebrafish using the TALEN technique with high efficiency. Surprisingly, the sharp decreases in mRNA expression and Gaa enzymatic activity in the gaa mutant larvae were restored from 2 dpf, although the Δ13 deletion was still confirmed to be present in the gaa mutant zebrafish. In addition, the Gaa protein levels were only slightly decreased in both the larvae and adult gaa mutant zebrafish, suggesting that the phenotype of the gaa mutants resembled the mild phenotype of Pompe disease in humans. gaa expression was decreased in both the mutant and WT zebrafish; this likely resulted from the persistence of maternal RNA, according to previous studies (Abrams and Mullins, 2009). Moreover, PAS staining of the heart, liver, and skeletal muscles suggested that more than half of the adult gaa mutant zebrafish exhibited abnormal glycogen storage. In addition, TEM revealed that the gaa mutants exhibited increased glycogen accumulation and that they possessed autophagic vacuoles filled with glycogen particles, similar to what has been found in other studies (Katona et al., 2014; Lim et al., 2014). Taken together, the findings demonstrated that the Δ13 deletion of the gaa gene partially affects the glycogen processing capacity in zebrafish.
We also attempted to identify whether alternative splicing isoforms existed in the mutants that may respond to TALEN mRNAs; however, no other splicing isoforms were found. A recent study has revealed an unexpected phenotypic variation between knockout and knockdown animals (Kok et al., 2015). Rossi et al. (2015) found that short-term morpholino knockdown induced a more severe disease phenotype in zebrafish than full gene knockout, which failed to induce an obvious phenotype. These authors deduced that a compensatory network may exist to buffer against the effects of deleterious mutations after translational or transcriptional knockdown. Moreover, organ or tissue regeneration is also a widely accepted phenomenon in some species. Higher vertebrate species, such as humans, have a limited capability for organ regeneration, while some lower vertebrate species, such as axolotls (McCusker and Gardiner, 2011), tadpoles (Slack et al., 2008), and zebrafish, have excellent regenerative capabilities (Goessling and North, 2014). The above two mechanisms could be the main underlying reasons why the gaa mutants in this study failed to develop the classical, severe, infantile-onset phenotype of Pompe disease. In view of this, the compensatory network and regenerative capacity of zebrafish may have great value in the search for more efficient alternative therapies for GSD II disease.
Conclusions
We have successfully generated a gaa gene frameshift mutation zebrafish model. The mild phenotype of the mutant zebrafish suggests that underlying compensatory mechanisms may exist in zebrafish. Further studies are necessary to elucidate the potential gaa gene network for the development of classic Pompe disease models in zebrafish.
Footnotes
Acknowledgments
The authors thank all the staff members of the zebrafish laboratory of Children's Hospital of Fudan University and School of Basic Medical Sciences, Fudan University, for their invaluable contributions to this project. They are also grateful to Dr. Jieaksu Su, Department of Pathology, Zhong Shan Hospital of Fudan University, for the kind support with pathological analysis. F.L. was supported by the Minhang District Talented Development Foundation and the Development Project of Shanghai Peak Disciplines-Integrated Chinese and Western Medicine. This study was supported by the Chinese National Twelfth Five Year Key Science and Technology Plan (2012BAI09B00) and the Shanghai Key Laboratory of Birth Defect, Children's Hospital of Fudan University new developing project (2014–2015).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.