
Abstract
It has been reported that microRNAs (miRNAs) are responsible for acquiring all the hallmarks of cancer cells, as well as have a significant impact on the clinical management of cancers at every stage, including prognosis, remission, relapse, and metastasis. In this study, we investigated the association of miR-29a-3p, miR-202-3p, miR-3713, miR-4768-3p, and miR-548aa expression with clinicopathologic features in patients suffering from laryngeal cancer (LC) and determined the potential role of studied miRNAs in the progression of LC. The study group consisted of 48 patients with untreated primary tumors of head and neck cancer localized in the larynx. Expression of the selected miRNAs was verified by the qRT-PCR technique. We showed that the expression of miR-29a as well as miR-548aa was positively correlated with tumor stage and lymph node metastasis, whereas the expression of miR-4768-3p was negatively correlated with lymph node metastasis. Furthermore, we investigated that exposure to cigarette smoke altered miRNA expression profile in LC. The expression level of miR-202-3p was significantly increased in smoking patients compared with nonsmokers, whereas the miR-4768-3p, miR-548aa, and miR-3713 were markedly decreased. Our research contributed toward better elucidating the mechanisms underlying the progression of LC as well as the use of miRNAs inhibitors as novel agents against progression and metastasis of LC.
Introduction
O
In recent years, much research has focused on the role of miRNAs in metastasis. The results of these studies have shown that specific miRNAs play a role in the different stages of metastasis, including the ability to modulate epithelial–mesenchymal transition (EMT), migration/invasion, angiogenesis, anoikis survival, and distant organ colonization. In the vast majority (∼90%), tumors are of an epithelial origin (Chan and Wang, 2015). It is considered that the process by which epithelial cells acquire greater mobility and the ability to migrate is EMT. It is connected with the change of the rate of proteins synthesis, including cell surface markers (e.g., E-cadherin), proteins that are responsible for intercellular contacts and interactions with the extracellular matrix (ECM), (integrins, ECM proteins, focal adhesion kinase [FAK] protein family), and the rearrangement of cytoskeletal membrane protein (the Rho protein family) (Christiansen and Rajasekaran, 2006).
The epithelial cancer cells are characterized by reductions of E-cadherin expression. In colorectal and breast cancers, the effect of E-cadherin repression by miR-9 and metastasis promotion has been directly observed (Ma et al., 2010; Lu et al., 2012). On the other hand, miR-30a by targeting Snai1 leads to an increase of E-cadherin expression, thereby inhibiting the ETM (Kumarswamy et al., 2012). Furthermore, in breast cancer, upregulation of miR-29a is observed, which suppressed the expression of tristetraprolin, a protein involved in the degradation of messenger RNAs with AU-rich 3′-untranslated regions, and, consequently, caused ETM and metastasis (Gebeshuber et al., 2009). In turn, another study has demonstrated that miR-126 has a binding site in 3′-UTR region of the VEGF-A mRNA, thereby causing a reduction of vascular endothelial growth factor (VEGF) level, which may inhibit angiogenesis.
Studies performed in eight lung cancer cell lines have shown that the expression level of miR-126 was significantly decreased compared with healthy cell lines (Liu et al., 2009). miRNAs belong to the miR-548 family and have also been implicated in tumor progression. In ovarian cancer, overexpression of miR-548c directly downregulates Twist, an oncogene that promotes the induction of EMT, proliferation, and angiogenesis (Sun et al., 2016). A laryngeal cancer (LC) study has demonstrated that miR-192, miR-143, miR-409, and miR-634 might promote ETM by targeting inhibition KLF17 factor, E-cadherin, and kinase PI3K (Tai et al., 2013). In turn, miR-206 suppresses LC cell proliferation, migration, and invasion through inhibiting VEGF (Li et al., 2014). Similarly, miR-139 inhibits proliferation and metastasis of LC by targeting CXCR4 (Luo et al., 2014).
One of the main risk factors for LC is cigarette smoking. Many studies have shown that cigarette smoking has an influence on changes in the expression level of many miRNAs. Moreover, researchers have shown that cigarette smoke causes carcinogenesis by miRNA deregulation. Cigarette smoking affects the genetic or epigenetic damage in miRNA genes, mainly in single-nucleotide polymorphism regions (Russ and Slack, 2012). In addition, it has been found that the miRNA dysregulation by tobacco smoke may cause disrupting mechanisms that are regulated by miRNAs, including the P53 pathway, which is the main pathway regulating cell cycle, cell growth, apoptosis, or angiogenesis (Ahrendt et al., 2000).
Due to the lack of data in the literature, the aim of our study was to evaluate the level of miR-29a-3p, miR-202-3p, miR-3713, miR-4768-3p, and miR-548aa expression in different stages of LC progression. In addition, we investigated the correlation of differentially expressed miRNAs with tobacco exposure among LC patients.
Materials and Methods
Study subjects
All subjects included in the study were unrelated Caucasian people living in Poland. The study group consisted of 48 patients—39 men (mean age 61 ± 9) and 9 women (mean age 56 ± 6) with newly diagnosed, untreated primary tumors of head and neck cancer localized in the larynx. Cancer type was confirmed with a histopathology examination. The study cohort included consecutive patients who underwent total or partial laryngectomy surgery at Public Central Clinical Hospital in Warsaw, Poland. Macroscopic tumor samples were collected as far as possible from the necrosis area. Tissue samples obtained during surgery were immediately fixed in 10% buffered formalin for at least 4 h and processed for paraffin embedding.
Before examination, patients did not receive medicaments such as antibiotics or steroids. Subjects enrolled to the examination were analyzed according to the cancer staging system of the TNM Classification of Malignant Tumors (Edge and Compton, 2010). Among the patient groups, seven subjects were classified as T1, 12 as T2, 12 as T3, and 17 as T4. Eleven patients were considered as having two local lymph nodes metastases (N2), and one person had three local lymph nodes metastases (N3). All the subjects were free from distant metastases (M0).
We interviewed each eligible participant to obtain data on tobacco smoking. The patient group subjects were classified as nonsmokers (n = 5), smokers for less than 20 years (n = 18), or smokers for more than 20 years (n = 24). Smoking subjects enrolled into the study are heavy smokers, smoking at least 20 cigarettes daily. All patients were recruited from medical units of Head and Neck Neoplasm Surgery Departments, Medical University of Warsaw, Poland. The study was approved by the Ethic Committee of the Medical University of Warsaw, and written consent was obtained from each patient or healthy subject before enrolling them into the study.
RNA isolation, cDNA synthesis, and real-time quantitative PCR
RNA was extracted from the HNSCC paraffin-embedded tissues by using the BiOstic® FFPE Tissue RNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA) according to the manufacturer's instructions. RNA quantity and purity were assessed via spectrophotometry by using Synergy HT (Bio-Tek, Winooski, VT). RNA with a 260/280 nm absorbance ratio in the range of 1.8–2.0 was used for further experiments. RNA quality was examined for 20% samples by electrophoresis on a 1% standard agarose gel. The ribosomal RNA (rRNA) bands were examined. If the upper ribosomal band (28S rRNA) was about twice the intensity of the lower band (18S rRNA) and smears occurred below and above rRNA bands, isolated RNA was utilized for cDNA synthesis.
RNA was reverse transcribed into cDNA by using the TaqMan MicroRNA Reverse Transcription Kit with specific, validated TaqMan primers for mature miRNA (Table 1) and RNU6B as a normalizer gene (Applied Biosystems, Foster City, CA). Quantitative real-time PCR for mature miRNAs and internal control RNU6B were performed according to the manufacturer's instructions for TaqMan MicroRNA Assays by using TaqMan Universal Master Mix II, no. UNG (Applied Biosystems). Samples were run in three repeats, and all qRT-PCR experiments included both no-reverse transcriptase (RT) and no-template controls. The RT-qPCR reaction was carried out by using the Mx3005P from Agilent Technologies.
Applied Biosystems, Carlsbad, CA.
Statistical analysis
miRNA expression was normalized to RNU6B by using the 2−ΔΔCt method (Livak and Schmittgen, 2001). Otherwise, the fold difference for each sample was obtained by using the following equation 2−ΔCt, where Ct is the Threshold Cycle, the cycle number at which the fluorescence generated within a reaction crosses the threshold; ΔCt = Ct average sample − Ct average RNU6B.
The mean ± standard deviation of fold changes in expression levels were calculated. To estimate the statistical significance, the following tests were performed: Shapiro-Wilk Normality test, F-test for equality of two variances followed by proper t test (Student's t-test or Welch t-test if the variances were not equal).
All tests were two sided, and values of p < 0.05 were considered statistically significant. Benjamini and Hoghberg correction of the p values (p adj) was used when the testing of multiple hypotheses was performed to control the false discovery rate, with a false discovery rate of 0.05. All analyses were performed by using STATISTICA 12 software (Statsoft, Tulsa, OK).
Results
miR expression analysis according to tumor stage
First, we compared the levels of different miRNAs levels at every stage of tumor development. Evaluated samples were subdivided according to tumor stage as T1, T2, T3, and T4 according to TNM classification. Only tissues from patients without metastasis were analyzed. miR-29a level was significantly lower in patients with T1 (8.610-fold) than in T2 (22.534-fold) (p adj = 0.037) or in comparison to T3 (19.744-fold, p adj = 0.038) (Fig. 1A). In addition, differences in levels of miR-29a were statistically significant between T2 and T3 groups (p adj < 0.001) (Fig. 1A).

Analysis of miR expression level according to T stage in larynx cancer tissues: miR-29a
miR-202 expression was decreased in the T1 group (0.035-fold) in comparison to the T3 group (0.063-fold, p adj = 0.029), but after Benjamini and Hoghberg correction, it lost the statistical significance (p adj = 0.087; Fig. 1B).
miR-548 was detected only in the T4 group, whereas it was not detected in other groups (Fig. 1C). There were no statistically significant changes of miR-3713 and miR-4768 levels between the investigated groups (p > 0.05) (Fig. 1D, E).
miR expression analysis according to lymph node metastases
Second, the miR expression level correlations with lymph node metastasis in different cancer stages were evaluated. Analyzed tissues were divided according to tumor stage as T2, T3, and T4 and subdivided according to the lymph node metastasis presence (N+)/absence (N−).
Performed analysis showed the statistically significant decrease in miR-29a level in T2N+ patients compared with T2 metastases-free subjects (0.514-fold, p = 0.034) (Fig. 2). On the other hand, miR-548 level was significantly increased in T4 patients with regional lymph node metastasis (2.454-fold; p = 0.030) (Fig. 2).
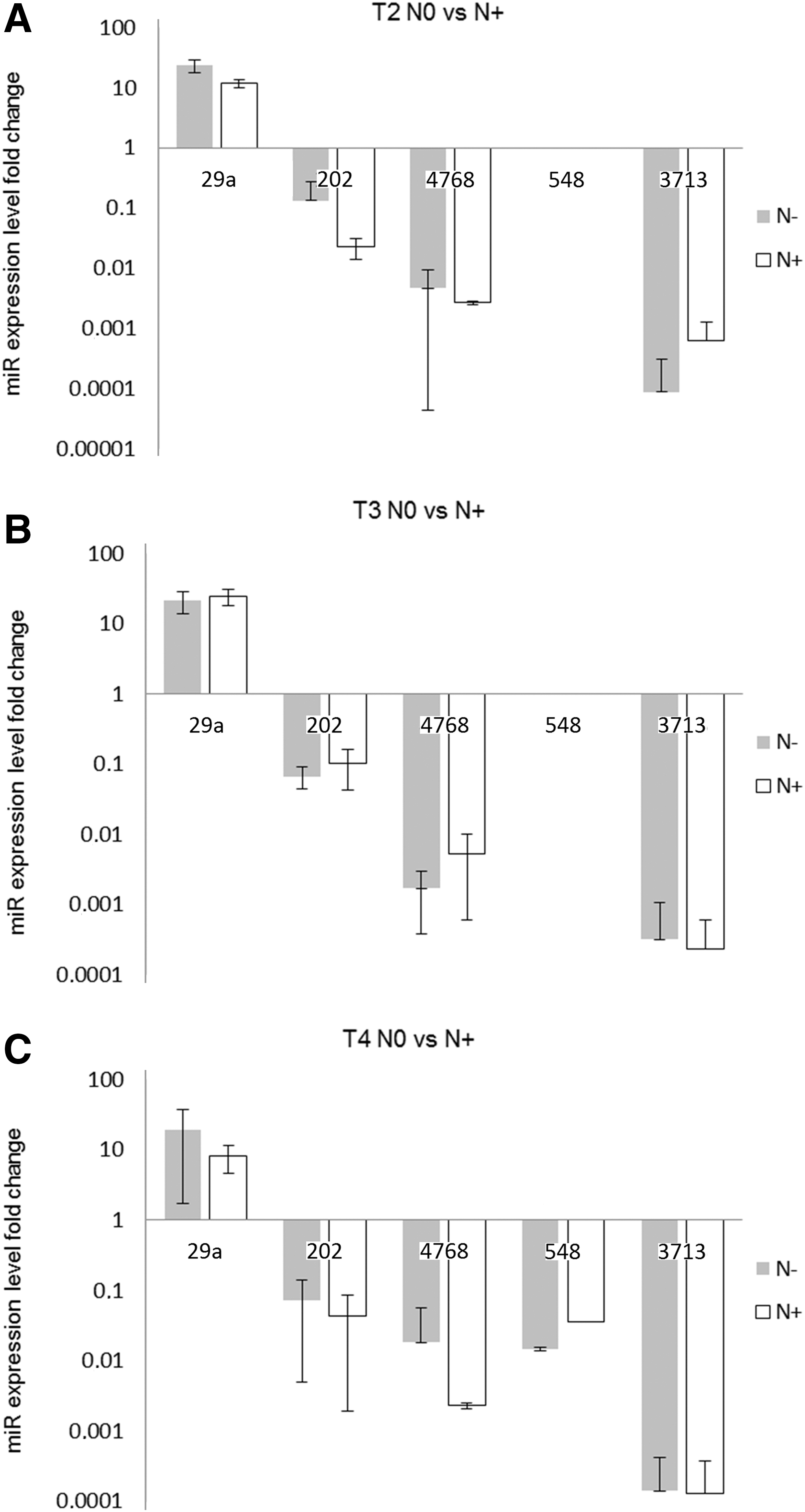
Analysis of miR expression level according to lymph node metastases versus metastases-free subjects in larynx cancer tissues by using the ΔCt method: T2N0 versus N+
In addition, miR expression levels of samples from patients with lymph node metastases were normalized to samples with the same tumor stage, but they were metastases free by using the ΔΔCt method. The analysis of normalized miR expression between different tumor stages revealed that the level of miR-29a in T2N+ is lower than in T3N+ patients (0.514-fold vs. 1.214-fold, respectively, p adj = 0.044) (Fig. 3). Moreover, there was a statistically significant difference between T2N+ and T4N+ subjects in miR-4768 levels (1.297- and 0.855-fold, respectively, p adj = 0.018) (Fig. 3).
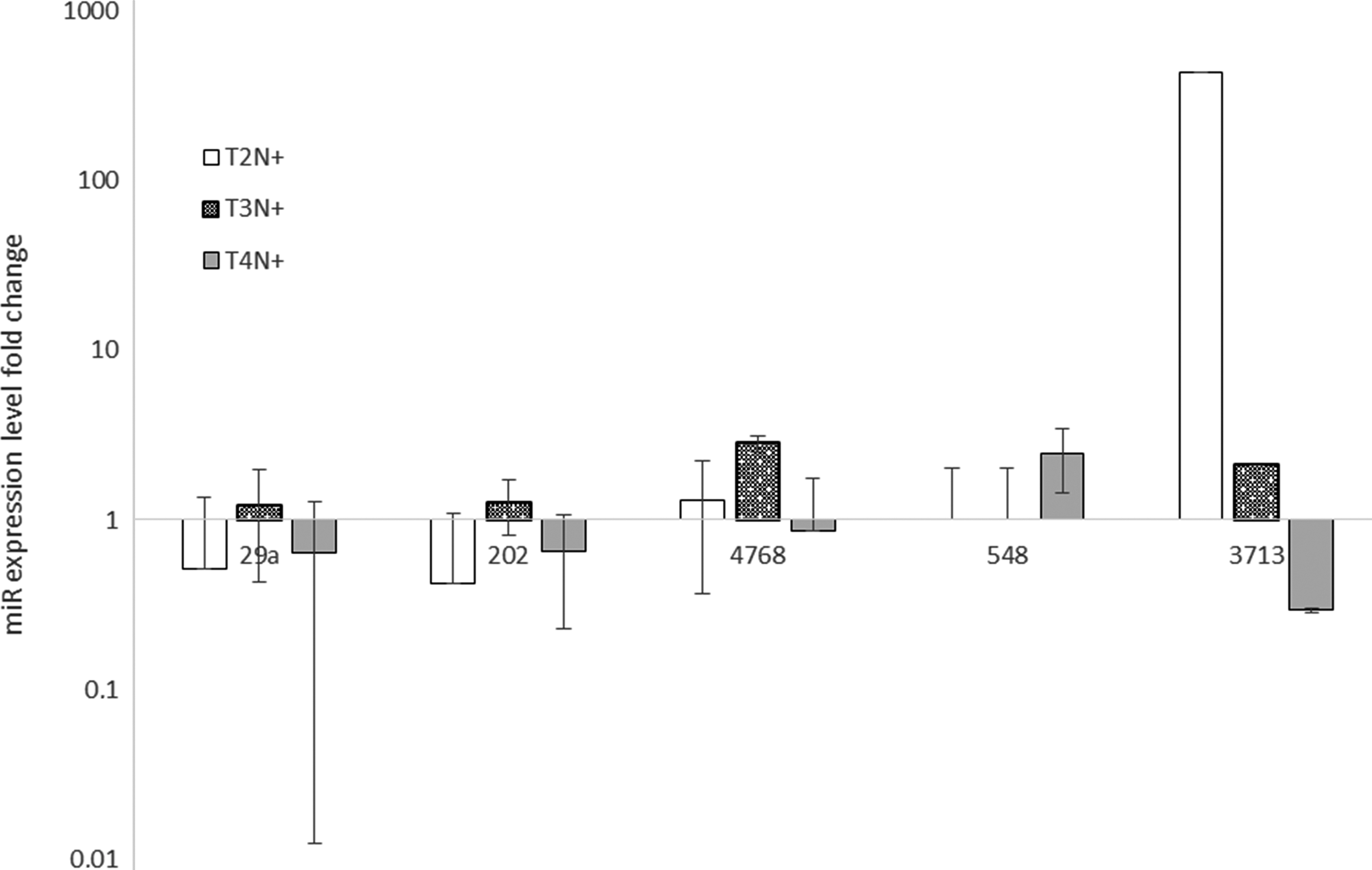
Analysis of miR expression level according to lymph node metastases in larynx cancer tissues by using the ΔΔCt method. miR expression was examined in primary tumor samples (n = 48) of head and neck cancer patients by quantitative RT-PCR, and statistical significance was analyzed by t-test. Error bars represent SD.
miR expression analysis according to smoking status
The relationship of expression level with smoking status was also evaluated. Tissue samples were divided according to patients' tobacco smoking habits as nonsmokers, subjects smoking less than 20 years, and patients smoking more than 20 years. The miR expression levels of samples from smoking patients were normalized to samples obtained from nonsmokers by using the ΔΔCt method.
First, we evaluated the differences in miRs expression level between smokers and nonsmokers (Fig. 4). Only miR-202 expression was markedly upregulated in patients smoking more than 20 years (7.916-fold, p adj = 0.005) (Fig. 4). The miR-29a level in this group was also elevated, but the level remained statistically insignificant (2.03-fold, p adj = 0.096). Other miRs levels were decreased in comparison with nonsmokers. miR-4768 expression was downregulated in the group of patients smoking less than 20 years (0.112-fold, p adj = 0.036). On the other hand, miR-548 level was markedly lower in patients smoking more than 20 years (2.577 × 10−8-fold, p adj = 0. 004). The miR-3713 expression was decreased in both groups: subjects smoking less than 20 years and those smoking more than 20 years (0.003- and 0.011-fold, p adj = 0.049 and p adj = 0.049, respectively).
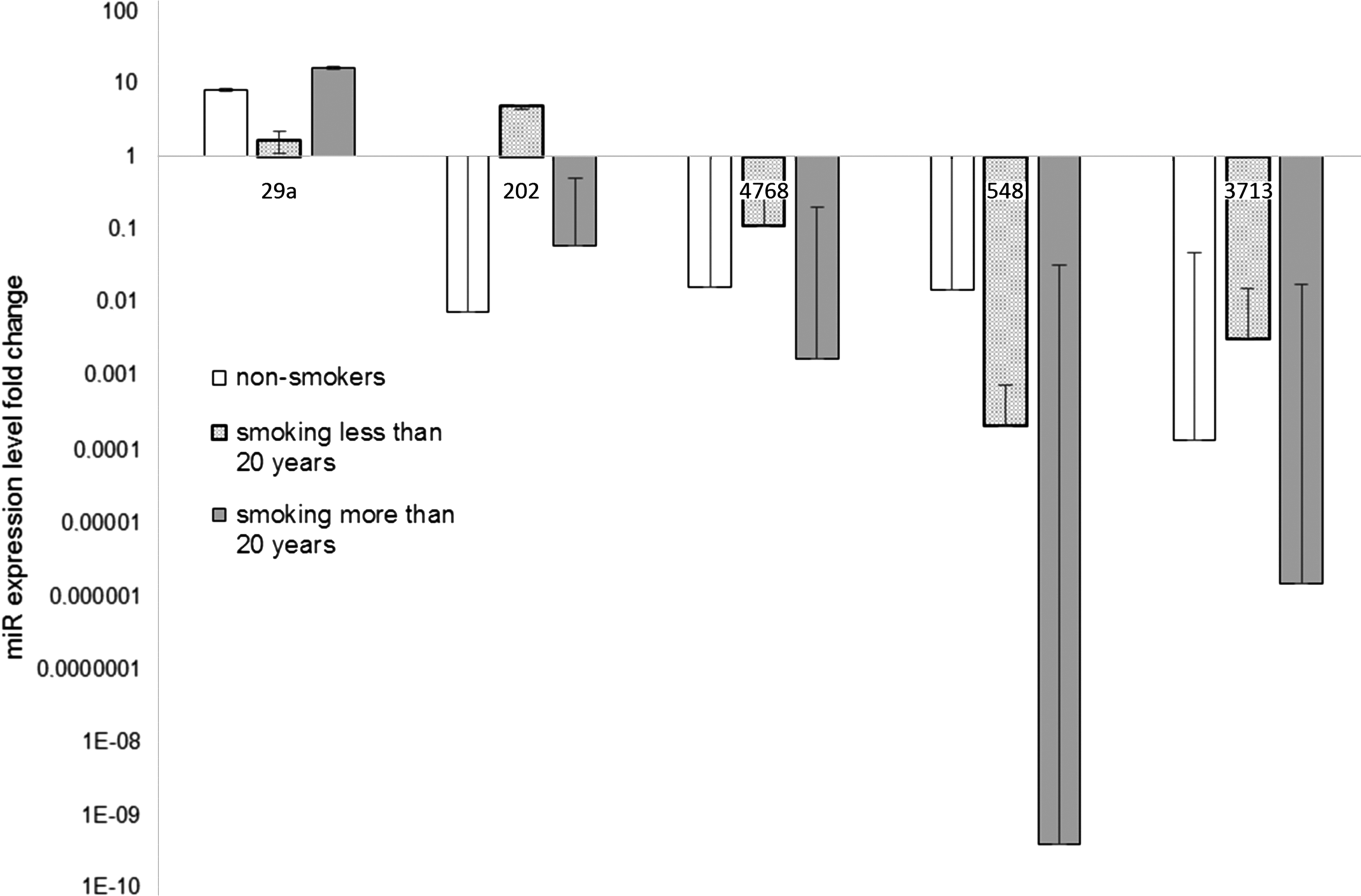
Analysis of miR expression level according to smoking status in larynx cancer tissues by using the ΔCt method. miR expression was examined in primary tumor samples (n = 48) of head and neck cancer patients by quantitative RT-PCR, and statistical significance was analyzed by t-test. Error bars represent SD.
We also compared levels of miR expression between samples obtained from people smoking more than 20 years and those smoking less than 20 years normalized to nonsmokers. We did not observe any significant differences in the investigated miR levels between the investigated groups (Fig. 5).
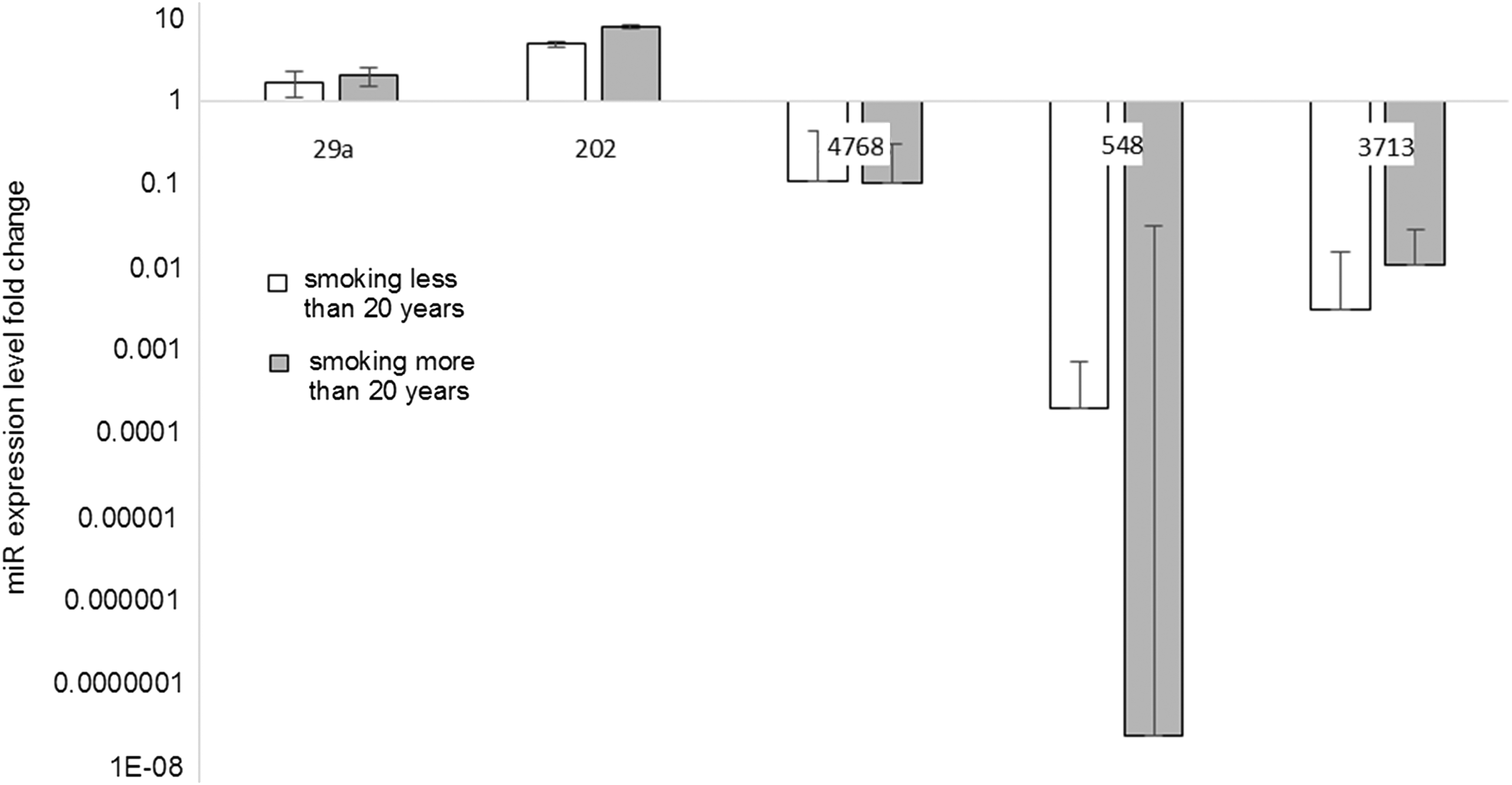
Analysis of miR expression level according to smoking status in larynx cancer tissues by using the ΔΔCt method. miR expression was examined in primary tumor samples (n = 48) of head and neck cancer patients by quantitative RT-PCR, and statistical significance was analyzed by t-test. Error bars represent SD.
Discussion
The molecular mechanisms of LC still remain unexplained. Much research has demonstrated that defective DNA repair mechanisms, impaired cell cycle, and aberrant expression of the metalloproteinases might increase the risk of LC occurrence (Peschos et al., 2005; Rusin et al., 2009; Walczak et al., 2012; Liu et al., 2015). In recent years, the increasing number of reports about the role of miRNAs in the development of LC has been noticed.
Our study has shown that the expression of miR-29a was positively correlated with TNM stage. In relation to tumor stage, the expression level of miR-29a was statistically significantly lower in patients with T1 (8.61-fold) than in T2 (22.53-fold) (p adj = 0.037) or in comparison to T3 (19.74-fold, p adj = 0.038) (Fig. 1A). In addition, the differences between levels of miR-29a were statistically significant between T2 and T3 (p adj < 0.001) (Fig. 1A). In the case of metastasis to the lymph nodes, the expression level of miR-29a was significantly decreased in T2N+ patients compared with T2 metastases-free subjects (0.514-fold, p = 0.034). Nevertheless, we observed that miR-29a is overexpressed in T3N+ compared with T2N+ patients (0.514-fold vs. 1.214-fold, respectively, p adj = 0.044). miR-29a belongs to the miR-29 family, which also includes miR-29b and miR-29c (Jiang et al., 2014).
The miR-29 family is expressed in various tissues, where it plays a key role in the homeostasis of ECM. However, many studies have indicated that ECM proteins are significantly deregulated and specific matrix components promote tumor progression and metastatic spread (Oskarsson, 2013). Tang et al. (2014) have demonstrated that increased expression of miR-29a promoted colorectal cancer metastasis by regulating matrix metalloproteinase 2, the enzyme involved in the remodeling of ECM components, through direct targeting of KLF4.
Another study, conducted by Qiu et al., has also confirmed our findings regarding the participation of miR-29a in the migration and invasion of cancer cells. They have found overexpression of miR-29a as well as miR-29b in tissue and serum samples of nasopharyngeal carcinoma. Deregulation of miR-29a/b expression was correlated with decreased expression of SPARC and COL3A1, other proteins involved in the remodeling of ECM, which suggested that SPARC and COL3A1 could be targets of miR-29a/b (Qiu et al., 2015).
In our research, we also observed the relationship of miR-548aa expression level with the progression of LC. MiR-548aa was detected only in the T4 group, whereas it was not detected in other groups. Moreover, its expression level in T4 patients with regional lymph node metastasis was significantly higher than in T4 metastases-free subjects (2.45-fold; p = 0.030).
miR-548aa is a member of the miR-548 family, which has been demonstrated to be involved in the development of several cancers. In the literature, there are no reports on the role of miR-548aa in the malignancies; however, there are new data about other members of the miR-548 family. Sun et al. have demonstrated the downregulation of miR-548c in endometrial and ovarian cancer tissues compared with normal tissues, which is correlated with poor prognosis in cancer patients. Furthermore, knockdown of miR-548c in endometrial and ovarian cancer cells lines (HEC-1 and OVCAR3, respectively) dramatically induces cancer cell proliferation, migration, and invasion (Sun et al., 2016).
Other studies, which have shown an inverse correlation between miR-548 expression and progression of cancer, were conducted on breast cancer cell lines. It has been observed that miR-548-3p was downregulated in clinical specimens and in breast cancer cell lines. Moreover, overexpression of miR-548-3p inhibited the proliferation and promoted the apoptosis of breast cancer cells (Shi et al., 2015).
Our study has also shown that the expression level of miR-4768-3p was significantly lower in T4 with lymph nodes metastasis compared with T2 metastases-free subjects (0.855- and 1.297-fold, respectively, p adj = 0.018). MiR-4768-3p also seems to have an effect on the progression of other types of cancer. In colorectal cancer, miR-4768-3p was significantly downregulated in stage IV tumors (Xu et al., 2014). In turn, our results have also demonstrated that miR-202-3p has a tendency to be upregulated in the T3 group compared with the T1 group (0.035- and 0.063-fold, respectively, p adj = 0.087).
This is the first report on the role of miR-202-3p in the LC. Until now, miR-202-3p was observed to be deregulated in gastric cancer, breast cancer, colorectal cancer, follicular lymphoma, and leukemia (Iorio et al., 2005; Ng et al., 2009; Hoffman et al., 2013; Zhao et al., 2013). In gastric cancer, miR-202-3p expression levels did not show any relationship with tumor location, local invasion of cancer, lymph node metastasis, or TNM stage (Zhao et al., 2013). However, we indicated no statistical difference in the expression level of miR-3713 depending on the TNM stage, as well as the literature data do not describe the association of this miRNA with LC and other type of cancers.
Several studies have recently demonstrated that exposure to cigarette smoking in human and animal models causes alternations in miRNAs expression (Izzotti et al., 2009, 2011; Schembri et al., 2009; Seike et al., 2009). Dysregulation of the miRNAs is closely related with an altered expression of their target mRNA, and therefore may be implicated with carcinogenesis.
In our study, we revealed that some miRNAs were dysregulated in smoking and nonsmoking LC patients. miR-202-3p was overexpressed in patients smoking more than 20 years compared with nonsmokers (7.916-fold, p adj = 0.005). In turn, miR-4768-3p was downregulated in the group of patients smoking less than 20 years (0.112-fold, p adj = 0.036). On the other hand, miR-548aa level was markedly lower in patients smoking more than 20 years (2.577 × 10−8-fold, p adj = 0.004). The miR-3713 was decreased in both groups: subjects smoking less than and more than 20 years (0.003- and 0.011-fold, p adj = 0.049 and p adj = 0.049, respectively).
The obtained results have shown that cigarette smoking, as one of the major risk factors of LC, may also modulate the level of expression of various miRNAs. Among lung cancer patients with or without smoking history, it has also been observed that mir-138 and let-7c were significantly downregulated in nonsmokers (Momi et al., 2014).
Conclusions
In conclusion, our research has provided insight into the role of some miRNAs in LC progression. We showed that the expression of miR-29a as well as miR-548aa was positively correlated with TNM stage, whereas the expression of miR-4768-3p was negatively correlated with lymph node metastasis. These findings suggested that miR-29a and miR-548aa might act as oncogenes, whereas miR-4768-3p may be a tumor suppressor in LC. Furthermore, we investigated that exposure to cigarette smoke-altered miRNA expression profile in LC. The expression level of miR-202-3p was significantly increased in smoking patients compared with nonsmokers, whereas the miR-4768-3p, miR-548aa, and miR-3713 were markedly decreased. However, further studies are needed to fully elucidate the miRNA-involved pathological regulatory mechanisms in LC.
Footnotes
Acknowledgments
This study was supported by grants from the National Centre for Research and Development NR 13-0101-10 and from the Medical University in Lodz 503/5-108-05/503-51-001.
Disclosure Statement
No competing financial interests exist.